Bronchoalveolar Lavage Fluid from COPD Patients Reveals More Compounds Associated with Disease than Matched Plasma

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
2.1. Cohort Characteristics
2.2. Compounds Detected in BAL and Plasma
2.3. Compound Associations with Clinical Covariates
2.4. Compound Associations with Plasma Cell-Counts
2.5. Compound Associations with BAL Cell-Counts
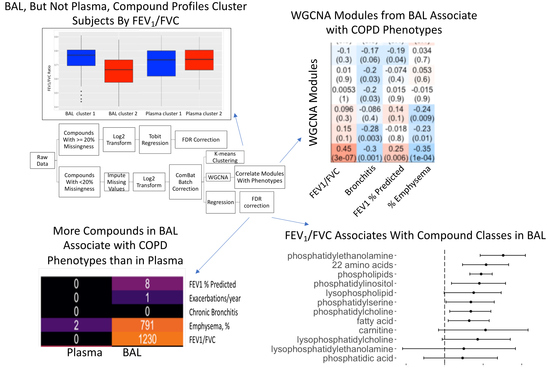
2.6. Compound Associations with COPD Phenotypes
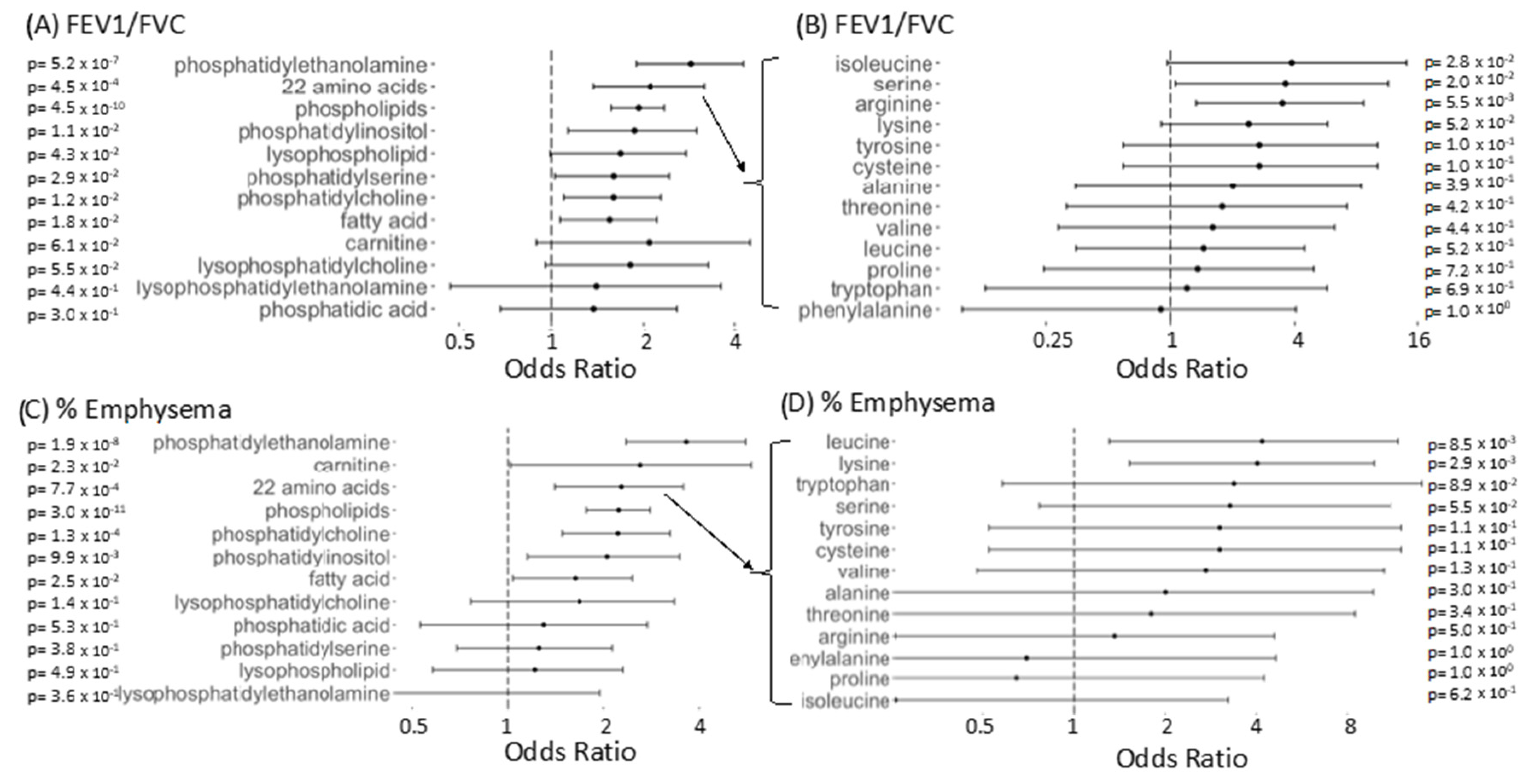
2.7. Compounds Most Highly Associated With Spirometry
2.8. Significantly Enriched Compound Classes
2.9. Co-Expressed BAL Compounds Grouped into Modules Associated with COPD Phenotypes
2.10. Grouping on Compound Profile Separated People with Differing Lung Function
3. Discussion
4. Materials and Methods
4.1. SPIROMICS
4.2. Clinical Variables and Definitions
4.3. Sample Preparation
4.4. Liquid Chromatography–Mass Spectrometry—Reversed Phase
4.5. Liquid Chromatography–Mass Spectrometry—Hydrophilic Interaction
4.6. Tandem Mass Spectrometry (MSMS)
4.7. Spectral Peak Extraction
4.8. Compound Identification
4.9. Data Processing and Analysis
4.10. Weighted Gene Co-Expression Network Analysis (WGCNA) Technique
4.11. Clustering
4.12. Classification of Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sullivan, J.; Pravosud, V.; Mannino, D.M.; Siegel, K.; Choate, R.; Sullivan, T. National and State Estimates of COPD Morbidity and Mortality — United States, 2014–2015. Chronic Obstr. Pulm. Dis. 2018, 5, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, A.L.; Lynch, D.; Dyar, L.A.; Bowler, R.P. Phenotypes of Chronic Obstructive Pulmonary Disease. COPD: J. Chronic Obstr. Pulm. Dis. 2007, 4, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Ubhi, B.K.; Riley, J.H.; Shaw, P.A.; Lomas, D.A.; Tal-Singers, R.; MacNeef, W.; Griffin, J.L.; Connor, S.C. Metabolic profiling detects biomarkers of protein degradation in COPD patients. Eur. Respir. J. 2012, 40, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Kan, M.; Shumyatcher, M.; Himes, B.E. Using omics approaches to understand pulmonary diseases. Respir. Res. 2017, 18, 149. [Google Scholar] [CrossRef] [PubMed]
- Haenen, S.; Clynen, E.; Nemery, B.; Hoet, P.H.M.; Vanoirbeek, J.A.J. Biomarker discovery in asthma and COPD: Application of proteomics techniques in human and mice. EuPA Open Proteom. 2014, 4, 101–112. [Google Scholar] [CrossRef]
- Bowler, R.P.; Jacobson, S.; Cruickshank, C.; Hughes, G.J.; Siska, C.; Ory, D.S.; Petrache, I.; Schaffer, J.E.; Reisdorph, N.; Kechris, K. Plasma Sphingolipids Associated with Chronic Obstructive Pulmonary Disease Phenotypes. Am. J. Respir. Crit. Care Med. 2015, 191, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Nobakht M. Gh, B.F.; Aliannejad, R.; Rezaei-Tavirani, M.; Taheri, S.; Oskouie, A.A. The metabolomics of airway diseases, including COPD, asthma and cystic fibrosis. Biomarkers 2015, 20, 516. [Google Scholar]
- Yu, B.; Flexeder, C.; McGarrah, R.; Wyss, A.; Morrison, A.; North, K.; Boerwinkle, E.; Kastenmüller, G.; Gieger, C.; Suhre, K.; et al. Metabolomics Identifies Novel Blood Biomarkers of Pulmonary Function and COPD in the General Population. Metabolites 2019, 9, 61. [Google Scholar] [CrossRef]
- Gregory, A.C.; Sullivan, M.B.; Segal, L.N.; Keller, B.C. Smoking is associated with quantifiable differences in the human lung DNA virome and metabolome. Respir. Res. 2018, 19, 174. [Google Scholar] [CrossRef]
- Evans, C.R.; Karnovsky, A.; Kovach, M.A.; Standiford, T.J.; Burant, C.F.; Stringer, K.A. Untargeted LC-MS metabolomics of bronchoalveolar lavage fluid differentiates acute respiratory distress syndrome from health. J. Proteome Res. 2014, 13, 640–649. [Google Scholar] [CrossRef]
- Conlon, T.M.; Bartel, J.; Ballweg, K.; Günter, S.; Prehn, C.; Krumsiek, J.; Meiners, S.; Theis, F.J.; Adamski, J.; Eickelberg, O.; et al. Metabolomics screening identifies reduced L-carnitine to be associated with progressive emphysema. Clin. Sci. 2016, 130, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.J.; Rebeles, J.; Dhungana, S.; Stewart, D.A.; Sumner, S.C.J.; Meyers, M.H.; Mancuso, P.; Beck, M.A. Obesity Increases Mortality and Modulates the Lung Metabolome during Pandemic H1N1 Influenza Virus Infection in Mice. J. Immunol. 2015, 194, 4846–4859. [Google Scholar] [CrossRef] [PubMed]
- Seemungal, T.A.R.; Lun, J.C.F.; Davis, G.; Neblett, C.; Chinyepi, N.; Dookhan, C.; Drakes, S.; Mandeville, E.; Nana, F.; Setlhake, S.; et al. Plasma homocysteine is elevated in COPD patients and is related to COPD severity. Int. J. Chron. Obstruct. Pulmon. Dis. 2007, 2, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Durga, J.; van Tits, L.J.H.; Schouten, E.G.; Kok, F.J.; Verhoef, P. Effect of Lowering of Homocysteine Levels on Inflammatory Markers. Arch. Intern. Med. 2005, 165, 1388. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, R.V.; Patel, S.G.; Guthikonda, A.P.; Reid, M.; Balasubramanyam, A.; Taffet, G.E.; Jahoor, F. Deficient synthesis of glutathione underlies oxidative stress in aging and can be corrected by dietary cysteine and glycine supplementation. Am. J. Clin. Nutr. 2011, 94, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Yoder, M.; Zhuge, Y.; Yuan, Y.; Holian, O.; Kuo, S.; van Breemen, R.; Thomas, L.L.; Lum, H. Bioactive lysophosphatidylcholine 16:0 and 18:0 are elevated in lungs of asthmatic subjects. Allergy Asthma Immunol. Res. 2014, 6, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Okazaki, T. The role of sphingomyelin and sphingomyelin synthases in cell death, proliferation and migration—From cell and animal models to human disorders. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2014, 1841, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Udyavar, A.R.; Hoeksema, M.D.; Clark, J.E.; Zou, Y.; Tang, Z.; Li, Z.; Li, M.; Chen, H.; Statnikov, A.; Shyr, Y.; et al. Co-expression network analysis identifies Spleen Tyrosine Kinase (SYK) as a candidate oncogenic driver in a subset of small-cell lung cancer. BMC Syst. Biol. 2013, 7 (Suppl. 5), S1. [Google Scholar] [CrossRef]
- Keck, M.; Androsova, G.; Gualtieri, F.; Walker, A.; von Rüden, E.-L.; Russmann, V.; Deeg, C.A.; Hauck, S.M.; Krause, R.; Potschka, H. A systems level analysis of epileptogenesis-associated proteome alterations. Neurobiol. Dis. 2017, 105, 164–178. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.-Z.; Zeng, Y.; Zhu, W.; Zhao, Y.-C.; Zhang, J.-G.; Zhu, J.-Q.; He, H.; Shen, H.; Tian, Q.; et al. Network-based proteomic analysis for postmenopausal osteoporosis in Caucasian females. Proteomics 2016, 16, 12–28. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, C.; Gearing, M.; Wang, P.G.; Chin, L.-S.; Li, L. Integrated proteomics and network analysis identifies protein hubs and network alterations in Alzheimer’s disease. Acta Neuropathol. Commun. 2018, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Kilk, K.; Aug, A.; Ottas, A.; Soomets, U.; Altraja, S.; Altraja, A. Phenotyping of Chronic Obstructive Pulmonary Disease Based on the Integration of Metabolomes and Clinical Characteristics. Int. J. Mol. Sci. 2018, 19, 666. [Google Scholar] [CrossRef]
- Halper-Stromberg, E.; Yun, J.H.; Parker, M.M.; Singer, R.T.; Gaggar, A.; Silverman, E.K.; Leach, S.; Bowler, R.P.; Castaldi, P.J. Systemic Markers of Adaptive and Innate Immunity Are Associated with Chronic Obstructive Pulmonary Disease Severity and Spirometric Disease Progression. Am. J. Respir. Cell Mol. Biol. 2018, 58, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Arenberg, D.A.; Barjaktarevic, I.; Bhatt, S.P.; Bowler, R.P.; Christenson, S.A.; Couper, D.J.; Dransfield, M.T.; Han, M.K.; Hoffman, E.A.; et al. Safety and Tolerability of Comprehensive Research Bronchoscopy in Chronic Obstructive Pulmonary Disease. Results from the SPIROMICS Bronchoscopy Substudy. Ann. Am. Thorac. Soc. 2019, 16, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.M.; Crudgington, S.; Stolberg, V.R.; Brown, J.P.; Sonstein, J.; Alexis, N.E.; Doerschuk, C.M.; Basta, P.V.; Carretta, E.E.; Couper, D.J.; et al. Design of a multi-center immunophenotyping analysis of peripheral blood, sputum and bronchoalveolar lavage fluid in the Subpopulations and Intermediate Outcome Measures in COPD Study (SPIROMICS). J. Transl. Med. 2015, 13, 19. [Google Scholar] [CrossRef]
- Cruickshank-Quinn, C.; Quinn, K.D.; Powell, R.; Yang, Y.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. Multi-step Preparation Technique to Recover Multiple Metabolite Compound Classes for In-depth and Informative Metabolomic Analysis. J. Vis. Exp. 2014, 89, 51670. [Google Scholar] [CrossRef] [PubMed]
- Stein, S.E.; Leader, W.W.T.; Leader, G.; Ji, W.; Tretyakov, D.S.; Edward, W.V.; Vladimir, Z.; Igor, Z.; Damo, Z.; Peter, L.; et al. NIST 17 MS Database and MS Search Program v.2.3 NIST Standard Reference Database 1A NIST/EPA/NIH Mass Spectral Library (NIST 17) and NIST Mass Spectral Search Program (Version 2.3) For Use with Microsoft ® Windows User’s Guide The NIST Mass Spectrometry Data Center 17 MS Database and MS Search Program v.2.2. 2017. Available online: https://www.nist.gov/srd/nist-standard-reference-database-1a-v17 (accessed on 19 June 2018).
- Yang, X.; Neta, P.; Stein, S.E. Quality Control for Building Libraries from Electrospray Ionization Tandem Mass Spectra. Anal. Chem. 2014, 86, 6393–6400. [Google Scholar] [CrossRef]
- Hughes, G.; Cruickshank-Quinn, C.; Reisdorph, R.; Lutz, S.; Petrache, I.; Reisdorph, N.; Bowler, R.; Kechris, K. MSPrep—Summarization, normalization and diagnostics for processing of mass spectrometry–based metabolomic data. Bioinformatics 2014, 30, 133–134. [Google Scholar] [CrossRef]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- Henningsen, A. Estimating Censored Regression Models in R Using the censReg Package. Available online: https://cran.r-project.org/web/packages/censReg/vignettes/censReg.pdf (accessed on 2 October 2019).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinformatics 2008, 9, 559. [Google Scholar] [CrossRef]
- Pei, G.; Chen, L.; Zhang, W. WGCNA: Application to Proteomic and Metabolomic Data Analysis. In Methods in Enzymology; Shukla, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 585, pp. 135–158. [Google Scholar]
- Langfelder, P.; Horvath, S. Tutorial for the WGCNA Package for R: I. Network Analysis of Liver Expression Data in Female Mice 2.b Step-by-Step Network Construction and Module Detection. 2014. Available online: https://horvath.genetics.ucla.edu/html/CoexpressionNetwork/Rpackages/WGCNA/Tutorials/FemaleLiver-02-networkConstr-man.pdf (accessed on 3 May 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Non-Smokers | Smoking Controls | COPD | p-Value |
|---|---|---|---|---|
| n | 12 | 56 | 47 | |
| Sex, % men | 33 | 45 | 62 | 0.104 |
| Race,% White | 50 | 73 | 87 | 7.48 × 10−3 * |
| Race, % Black | 25 | 21 | 6 | 7.48 × 10−3 * |
| Race, % Asian | 17 | 2 | 4 | 7.48 × 10−3 * |
| Race, % other | 8 | 4 | 2 | 7.48 × 10−3 * |
| Age, yr | 56 (50–60) | 58 (50–66) | 64 (58–68) | 7.95 × 10−4 * |
| Current smokers, % | 0 | 36 | 36 | 2.68 × 10−2 * |
| Pack–years | 0 (0–0) | 34 (26–44) | 42 (34–60) | 3.95 × 10−11 * |
| Body mass index | 26.21 (5.46) | 28.78 (4.47) | 28.9 (5.27) | 0.198 |
| Chronic bronchitis, % | 0 (0) | 7 (26) | 15 (36) | 0.294 |
| Exacerbations/yr | 0.08 (0.29) | 0.12 (0.43) | 0.39(0.68) | 0.117 |
| Emphysema, % | 0.15 (0.06–1.22) | 0.16 (0.05–0.4) | 1.05 (0.32–2.5) | 2.90 × 10−3 * |
| FEV1 % | 99.29 (7.31) | 100.23 (13.1) | 78.97 (19.92) | 3.87 × 10−8 * |
| FEV1/FVC | 81 (77–87) | 78 (75–81) | 61 (55–67) | 5.31 × 10−24 * |
| Variable | BAL | Plasma |
|---|---|---|
| Sex | 1 | 240 |
| Current Smoker | 249 | 7 |
| Age | 0 | 177 |
| Menopause | 0 | 0 |
| Neutrophil Count | 665 | 0 |
| Lymphocyte Count | 5 | 0 |
| Eosinophil Count | 0 | 4 |
| BAL Neutrophil Count | 0 | 4 |
| BAL Lymphocyte Count | 1 | 0 |
| BAL Eosinophil Count | 0 | 7 |
| BAL Monocyte Count | 1 | 0 |
| BAL Macrophage Count | 1 | 1 |
| Hemoglobin | 0 | 63 |
| Hematocrit | 0 | 80 |
| FEV1/FVC | 1230 | 0 |
| Emphysema, % | 791 | 2 |
| Chronic Bronchitis | 0 | 0 |
| Exacerbations/yr | 1 | 0 |
| FEV1 % | 8 | 0 |
| Compound | FDR BAL | Estimate BAL | SE BAL | FDR Plasma | Estimate Plasma | SE Plasma |
|---|---|---|---|---|---|---|
| PS (37:3) | 7.6 × 10−5 | 0.45 | 0.089 | 1 | 0.0015 | 0.094 |
| Lophocerine | 7.6 × 10−5 | 0.42 | 0.084 | 1 | −0.0034 | 0.066 |
| p-cresol | 7.6 × 10−5 | 0.4 | 0.08 | 0.98 | −0.036 | 0.14 |
| PE (38:3) | 7.6 × 10−5 | 0.38 | 0.075 | 0.93 | 0.086 | 0.094 |
| PC (40:6) | 7.6 × 10−5 | 0.35 | 0.069 | 0.11 | 0.14 | 0.033 |
| PC (40:6) (isomer) | 7.6 × 10−5 | 0.34 | 0.063 | 0.68 | −0.16 | 0.079 |
| Ceramide (d18:1/16:0) * | 7.6 × 10−5 | −0.29 | 0.054 | 0.89 | 0.092 | 0.086 |
| PC (32:1) ** | 7.6 × 10−5 | 0.28 | 0.054 | 0.96 | −0.048 | 0.082 |
| Glycocholic acid * | 7.6 × 10−5 | 0.27 | 0.052 | 0.96 | 0.023 | 0.035 |
| MGDG (36:5) | 7.6 × 10−5 | 0.27 | 0.055 | 0.89 | 21 | 20 |
| S-(Phenylacetothiohydroximoyl)-L-cysteine | 7.6 × 10−5 | 0.26 | 0.051 | 0.78 | −0.13 | 0.09 |
| SM (d18:1/24:1) ** | 7.6 × 10−5 | 0.26 | 0.051 | |||
| PE (35:1) | 7.6 × 10−5 | 0.26 | 0.05 | 0.96 | −0.036 | 0.075 |
| N-palmitoyl glycine | 7.6 × 10−5 | 0.25 | 0.05 | 0.92 | 17 | 20 |
| L-Threonylcarbamoyladenylate | 7.6 × 10−5 | 0.25 | 0.049 | 0.55 | −0.078 | 0.033 |
| Decaprenyl phosphate | 7.6 × 10−5 | 0.24 | 0.047 | 0.99 | −2.9 | 11 |
| Mycalamide B | 7.6 × 10−5 | 0.23 | 0.044 | 0.97 | −0.0099 | 0.027 |
| PC (36:4) * | 7.6 × 10−5 | 0.23 | 0.046 | 0.44 | 36 | 14 |
| PE (36:3) | 7.6 × 10−5 | 0.22 | 0.045 | 0.96 | 0.019 | 0.042 |
| PC (34:2) ** | 7.6 × 10−5 | 0.22 | 0.044 | 0.95 | 5.9 | 8.5 |
| Homocysteine * | 7.6 × 10−5 | 0.22 | 0.046 | 0.89 | 1.6 | 1.4 |
| SQMG (16:1) | 7.6 × 10−5 | 0.21 | 0.042 | 0.55 | −26 | 12 |
| PE (34:2) * | 7.6 × 10−5 | 0.2 | 0.039 | 0.98 | −0.019 | 0.081 |
| CL (70:0) | 9.2 × 10−5 | 0.27 | 0.056 | 0.98 | −0.015 | 0.071 |
| CL (72:7) | 9.4 × 10−5 | 0.40 | 0.082 | 1 | 0.001 | 0.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halper-Stromberg, E.; Gillenwater, L.; Cruickshank-Quinn, C.; O’Neal, W.K.; Reisdorph, N.; Petrache, I.; Zhuang, Y.; Labaki, W.W.; Curtis, J.L.; Wells, J.; et al. Bronchoalveolar Lavage Fluid from COPD Patients Reveals More Compounds Associated with Disease than Matched Plasma. Metabolites 2019, 9, 157. https://doi.org/10.3390/metabo9080157
Halper-Stromberg E, Gillenwater L, Cruickshank-Quinn C, O’Neal WK, Reisdorph N, Petrache I, Zhuang Y, Labaki WW, Curtis JL, Wells J, et al. Bronchoalveolar Lavage Fluid from COPD Patients Reveals More Compounds Associated with Disease than Matched Plasma. Metabolites. 2019; 9(8):157. https://doi.org/10.3390/metabo9080157
Chicago/Turabian StyleHalper-Stromberg, Eitan, Lucas Gillenwater, Charmion Cruickshank-Quinn, Wanda Kay O’Neal, Nichole Reisdorph, Irina Petrache, Yonghua Zhuang, Wassim W. Labaki, Jeffrey L. Curtis, James Wells, and et al. 2019. "Bronchoalveolar Lavage Fluid from COPD Patients Reveals More Compounds Associated with Disease than Matched Plasma" Metabolites 9, no. 8: 157. https://doi.org/10.3390/metabo9080157
APA StyleHalper-Stromberg, E., Gillenwater, L., Cruickshank-Quinn, C., O’Neal, W. K., Reisdorph, N., Petrache, I., Zhuang, Y., Labaki, W. W., Curtis, J. L., Wells, J., Rennard, S., Pratte, K. A., Woodruff, P., Stringer, K. A., Kechris, K., & Bowler, R. P. (2019). Bronchoalveolar Lavage Fluid from COPD Patients Reveals More Compounds Associated with Disease than Matched Plasma. Metabolites, 9(8), 157. https://doi.org/10.3390/metabo9080157