Improved Algal Toxicity Test System for Robust Omics-Driven Mode-of-Action Discovery in Chlamydomonas reinhardtii


Abstract
:1. Introduction
2. Results
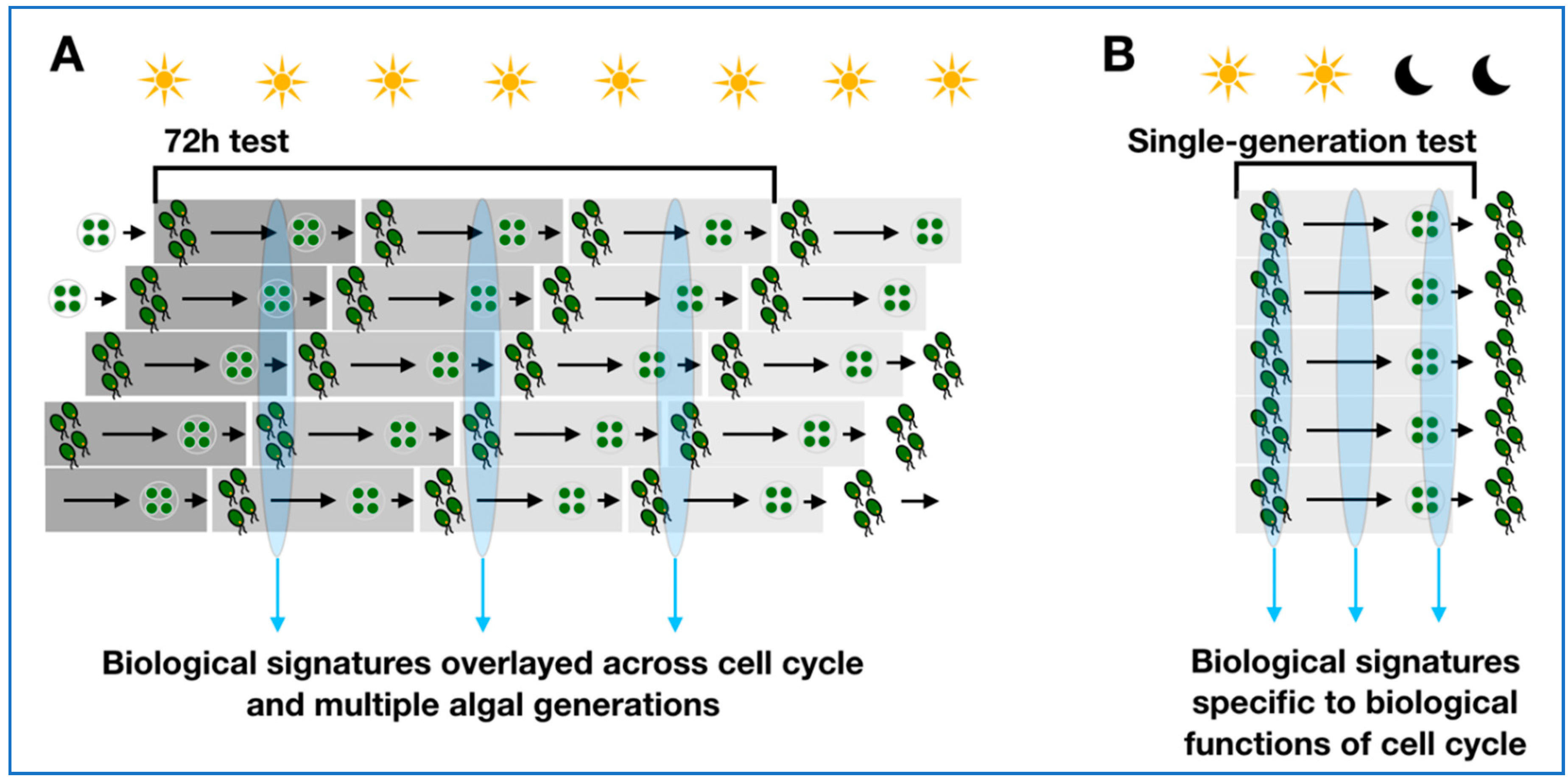
2.1. Synchronised Versus Non-Synchronised Algae Cultures for Molecular Studies
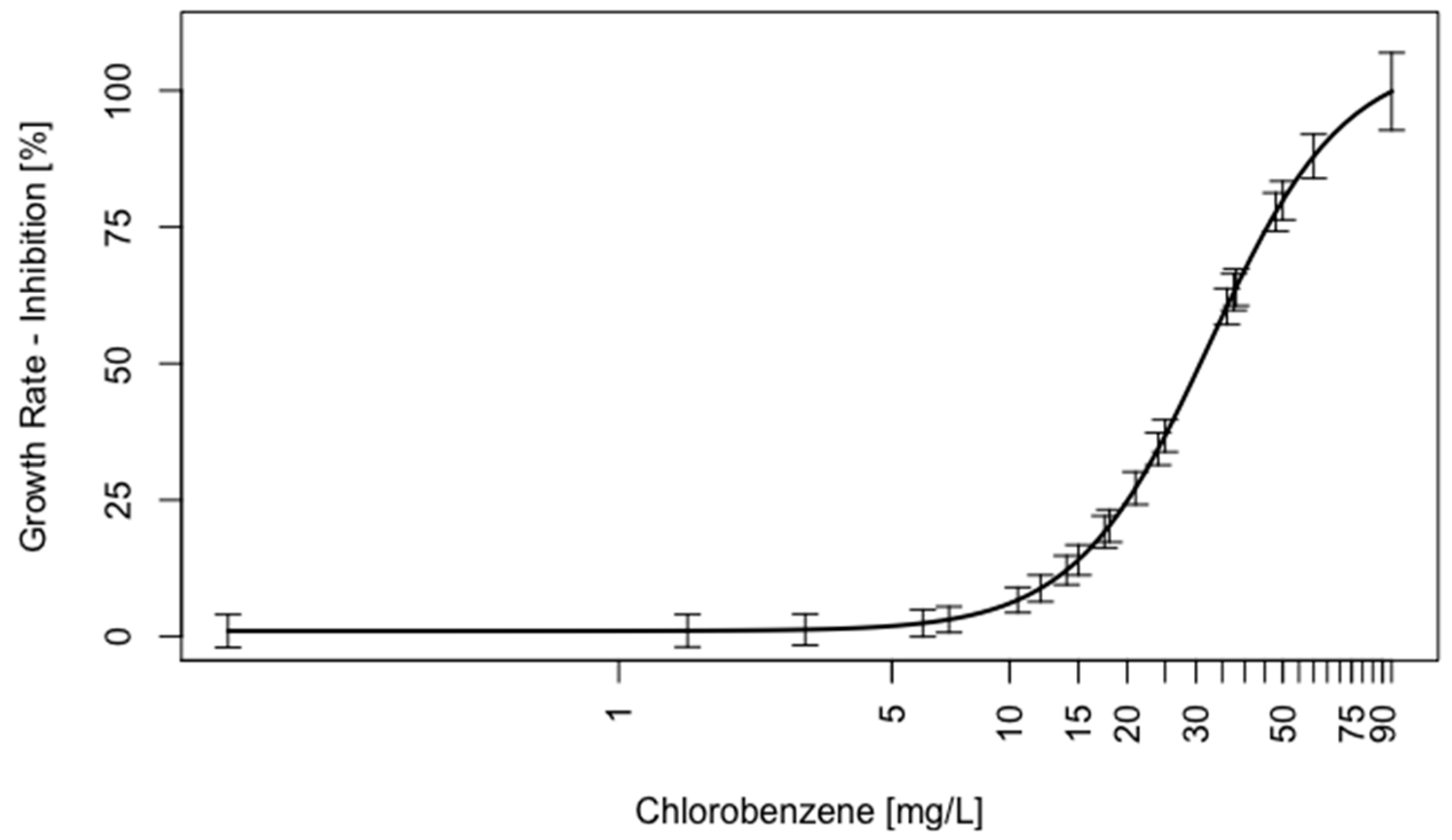
2.2. Validation of C. reinhardtii Test System for Volatile Substances
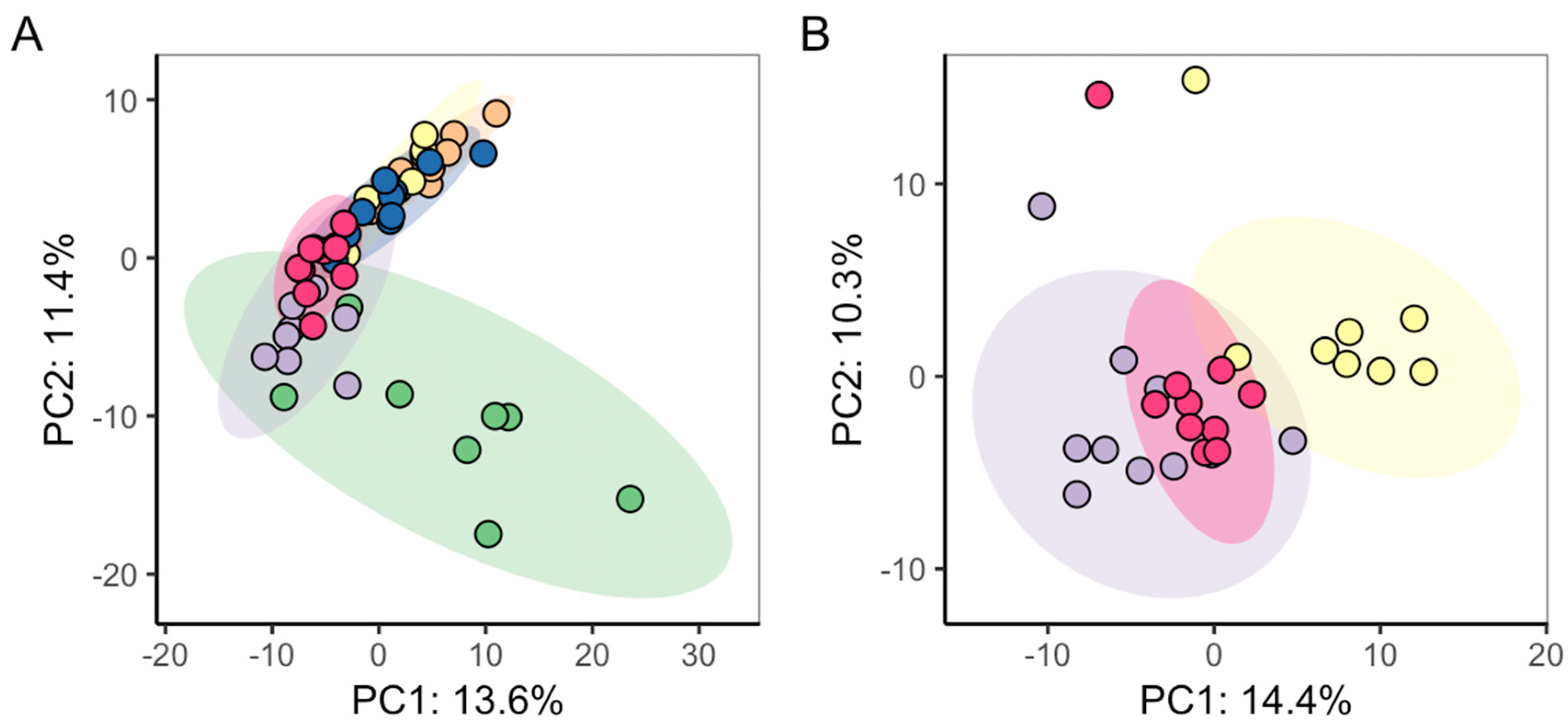
2.3. Repeatability of C. reinhardtii Metabolic Phenotypes in Test System
3. Discussion
3.1. Synchronised Cultures in Algal Toxicity Testing
3.2. Design and Validation of an Alternative Volatile Testing System
3.3. Characterising Variability and Repeatability of Metabolomics Data
4. Materials and Methods
4.1. Algae Culturing
4.2. Chlorobenzene Exposures
4.3. Sampling and Metabolite Extraction for DIMS Metabolomics
4.3.1. Sampling Procedure
4.3.2. Metabolite Extractions
4.4. Direct Infusion Mass-Spectrometry (DIMS) Metabolomics
4.5. Water Chemistry of Chlorobenzene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ECHA. Guidance on Information Requirements and Chemical Safety Assessment. Chapter R.10: Characterisation of Dose [Concentration]-Response for Environment; ECHA: Helsinki, Finland, 2008.
- ISO (International Organisation for Standardisation). ISO Water quality—Algal Growth Inhibition Test; ISO/DIS 8692; ISO: Geneva, Switzerland, 2012. [Google Scholar]
- OECD. Test No. 201: Alga, Growth Inhibition Test. In OECD Guidelines for the Testing of Chemicals, Section 2: Effects on Biotic Systems; OECD Publishing: Paris, France, 2011. [Google Scholar]
- ECHA. Usage of (Eco)Toxicological Data for Bridging Data Gaps between and Grouping of Nanoforms of the Same Substance; Elements to Consider ED-02-16-228-EN-N; ECHA: Helsinki, Finland, 2016. [Google Scholar]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef]
- Sauer, U.G.; Deferme, L.; Gribaldo, L.; Hackermüller, J.; Tralau, T.; van Ravenzwaay, B.; Yauk, C.; Poole, A.; Tong, W.; Gant, T.W. The challenge of the application of omics technologies in chemicals risk assessment: Background and outlook. Regul. Toxicol. Pharmacol. 2017, 91, 14–26. [Google Scholar] [CrossRef]
- Keller, D.A.; Juberg, D.R.; Catlin, N.; Farland, W.H.; Hess, F.G.; Wolf, D.C.; Doerrer, N.G. Identification and Characterization of Adverse Effects in 21st Century Toxicology. Toxicol. Sci. 2012, 126, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shostak, S. Exposed Science: Genes, the Environment and the Politics of Population Health; University of California Press: Berkeley, CA, USA, 2013. [Google Scholar]
- ECETOC. Workshop Report No. 25: Omics and Risk Assessment Science; ECETOC: Málaga, Spain, 2013. [Google Scholar]
- ICCVAM. Validation and Regulatory Acceptance of Toxicological Test Methods. In A Report of the ad hoc Interagency Coordinating Committee on the Validation of Alternative Methods; NIEHS: Durham, NC, USA, 1997; p. 123. Available online: https://ntp.niehs.nih.gov/iccvam/docs/about_docs/validate.pdf (accessed on 19 February 2019).
- Kanesaki, Y.; Imamura, S.; Minoda, A.; Tanaka, K. External Light Conditions and Internal Cell Cycle Phases Coordinate Accumulation of Chloroplast and Mitochondrial Transcripts in the Red Alga Cyanidioschyzon merolae. DNA Res. 2012, 19, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Kluender, C.; Sans-Piché, F.; Riedl, J.; Altenburger, R.; Härtig, C.; Laue, G.; Schmitt-Jansen, M. A metabolomics approach to assessing phytotoxic effects on the green alga Scenedesmus vacuolatus. Metabolomics 2009, 5, 59–71. [Google Scholar] [CrossRef]
- Vogs, C.; Bandow, N.; Altenburger, R. Effect propagation in a toxicokinetic/toxicodynamic model explains delayed effects on the growth of unicellular green algae Scenedesmus vacuolatus. Environ. Toxicol. Chem. 2013, 32, 1161–1172. [Google Scholar] [CrossRef]
- Antczak, P.; White, T.A.; Giri, A.; Michelangeli, F.; Viant, M.R.; Cronin, M.T.D.; Vulpe, C.; Falciani, F. Systems Biology Approach Reveals a Calcium-Dependent Mechanism for Basal Toxicity in Daphnia magna. Environ. Sci. Technol. 2015, 49, 11132–11140. [Google Scholar] [CrossRef]
- Bridges, J.; Sauer, U.G.; Buesen, R.; Deferme, L.; Tollefsen, K.E.; Tralau, T.; van Ravenzwaay, B.; Poole, A.; Pemberton, M. Framework for the quantitative weight-of-evidence analysis of ‘omics data for regulatory purposes. Regul. Toxicol. Pharmacol. 2017, 91, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Zhang, B.; He, Y.; Hu, C.; Ng, Q.X.; Zhang, H.; Ong, C.N. ZhifenLin Biological effect of aqueous C60 aggregates on Scenedesmus obliquus revealed by transcriptomics and non-targeted metabolomics. J. Hazard. Mater. 2017, 324, 221–229. [Google Scholar] [CrossRef]
- Norris, J.L.; Farrow, M.A.; Gutierrez, D.B.; Palmer, L.D.; Muszynski, N.; Sherrod, S.D.; Pino, J.C.; Allen, J.L.; Spraggins, J.M.; Lubbock, A.L.R.; et al. Integrated, High-Throughput, Multiomics Platform Enables Data-Driven Construction of Cellular Responses and Reveals Global Drug Mechanisms of Action. J. Proteome Res. 2017, 16, 1364–1375. [Google Scholar] [CrossRef]
- Nestler, H.; Groh, K.J.; Schönenberger, R.; Eggen, R.I.L.; Suter, M.J.F. Linking proteome responses with physiological and biochemical effects in herbicide-exposed Chlamydomonas reinhardtii. J. Proteom. 2012, 75, 5370–5385. [Google Scholar] [CrossRef]
- Jamers, A.; Van der Ven, K.; Moens, L.; Robbens, J.; Potters, G.; Guisez, Y.; Blust, R.; De Coen, W. Effect of copper exposure on gene expression profiles in Chlamydomonas reinhardtii based on microarray analysis. Aquat. Toxicol. 2006, 80, 249–260. [Google Scholar] [CrossRef]
- Pillai, S.; Behra, R.; Nestler, H.; Suter, M.J.F.; Sigg, L.; Schirmer, K. Linking toxicity and adaptive responses across the transcriptome, proteome, and phenotype of Chlamydomonas reinhardtii exposed to silver. Proc. Natl. Acad. Sci. USA 2014, 111, 3490–3495. [Google Scholar] [CrossRef]
- Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Cellular toxicity pathways of inorganic and methyl mercury in the green microalga Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 8034. [Google Scholar] [CrossRef] [PubMed]
- Esperanza, M.; Seoane, M.; Rioboo, C.; Herrero, C.; Cid, Á. Early alterations on photosynthesis-related parameters in Chlamydomonas reinhardtii cells exposed to atrazine: A multiple approach study. Sci. Total Environ. 2016, 554, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K.J. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 2013, 79, 4774–4785. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.F.; Descombes, P.; Zerges, W.; Wilkinson, K.J. Global expression profiling of Chlamydomonas reinhardtii exposed to trace levels of free cadmium. Environ. Toxicol. Chem. 2008, 27, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Yoo, W.I.; Lee, S.H.; Lee, M.Y. Proteomic analysis of cadmium-induced protein profile alterations from marine alga Nannochloropsis oculata. Ecotoxicol. Lond. Engl. 2005, 14, 589–596. [Google Scholar] [CrossRef]
- Pan, C.G.; Peng, F.J.; Shi, W.J.; Hu, L.X.; Wei, X.D.; Ying, G.G. Triclosan-induced transcriptional and biochemical alterations in the freshwater green algae Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 2018, 148, 393–401. [Google Scholar] [CrossRef] [PubMed]
- González-Pleiter, M.; Rioboo, C.; Reguera, M.; Abreu, I.; Leganés, F.; Cid, Á.; Fernández-Piñas, F. Calcium mediates the cellular response of Chlamydomonas reinhardtii to the emerging aquatic pollutant Triclosan. Aquat. Toxicol. 2017, 186, 50–66. [Google Scholar] [CrossRef]
- Jamers, A.; Blust, R.; De Coen, W.; Griffin, J.L.; Jones, O.A. Copper toxicity in the microalga Chlamydomonas reinhardtii: An integrated approach. BioMetals 2013, 26, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Walliwalagedara, C.; Keulen, H.; Willard, B.; Wei, R. Differential Proteome Analysis of Chlamydomonas reinhardtii Response to Arsenic Exposure. Am. J. Plant Sci. 2012, 3, 764–772. [Google Scholar] [CrossRef]
- Patel, N.; Cardoza, V.; Christensen, E.; Rekapalli, B.; Ayalew, M.; Stewart, C.N.N. Differential gene expression of Chlamydomonas reinhardtii in response to 2,4,6-trinitrotoluene (TNT) using microarray analysis. Plant Sci. 2004, 167, 1109–1122. [Google Scholar] [CrossRef]
- Wang, S.B.; Chen, F.; Sommerfeld, M.; Hu, Q. Proteomic analysis of molecular response to oxidative stress by the green alga Haematococcus pluvialis (Chlorophyceae). Planta 2004, 220, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Merrifield, R.; Williams, T.D.; Chipman, J.K.; Lead, J.R.; Viant, M.R. Molecular toxicity of cerium oxide nanoparticles to the freshwater alga Chlamydomonas reinhardtii is associated with supra-environmental exposure concentrations. Nanotoxicology 2015, 5390, 32–41. [Google Scholar]
- Jiang, Y.; Xiao, P.; Shao, Q.; Qin, H.; Hu, Z.; Lei, A.; Wang, J. Metabolic responses to ethanol and butanol in Chlamydomonas reinhardtii. Biotechnol. Biofuels 2017, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Rubinelli, P.; Siripornadulsil, S.; Gao-Rubinelli, F.; Sayre, R.T. Cadmium- and iron-stress-inducible gene expression in the green alga Chlamydomonas reinhardtii: Evidence for H43 protein function in iron assimilation. Planta 2002, 215, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gillet, S.; Decottignies, P.; Chardonnet, S.; Le Maréchal, P. Cadmium response and redoxin targets in Chlamydomonas reinhardtii: A proteomic approach. Photosynth. Res. 2006, 89, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, E.K.; Hodges, G.; Hutchinson, T.H.; Butler, E.; Hecker, M.; Tollefsen, K.E.; Garcia-Reyero, N.; Kille, P.; Becker, D.; Chipman, K.; et al. The Role of Omics in the Application of Adverse Outcome Pathways for Chemical Risk Assessment. Toxicol. Sci. 2017, 158, 252–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamers, A.; Blust, R.; De Coen, W.; Griffin, J.L.; Jones, O.A. An omics-based assessment of cadmium toxicity in the green alga Chlamydomonas reinhardtii. Aquat. Toxicol. 2013, 126, 355–364. [Google Scholar] [CrossRef]
- Parsons, H.M.; Ekman, D.R.; Collette, T.W.; Viant, M.R. Spectral relative standard deviation: A practical benchmark in metabolomics. Analyst 2009, 134, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Southam, A.D.; Weber, R.J.M.; Engel, J.; Jones, M.R.; Viant, M.R. A complete workflow for high-resolution spectral-stitching nanoelectrospray direct-infusion mass-spectrometry-based metabolomics and lipidomics. Nat. Protoc. 2017, 12, 255–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corvi, R.; Ahr, H.J.; Albertini, S.; Blakey, D.H.; Clerici, L.; Coecke, S.; Douglas, G.R.; Gribaldo, L.; Groten, J.P.; Haase, B.; et al. Meeting report: Validation of toxicogenomics-based test systems: ECVAM-ICCVAM/NICEATM considerations for regulatory use. Environ. Health Perspect. 2006, 114, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Faetsch, S.; Matzke, M.; Stolte, S. Application Note Omni Life Sciences: Monitoring the Cell Cycle of the Unicellular Green Algae Raphidocelis subcapitata. Analysis of Cell Growth and Proliferation Using CASY. 2017. Available online: https://www.ols-bio.de/media/pdf/Algae-CellCycle_CASY-AppNote_OLS.pdf (accessed on 10 May 2019).
- Grabski, K.; Aksmann, A.; Mucha, P.; Tukaj, Z. Conditioned medium factor produced and released by Desmosdemus subspicatus and its effect on the cell cycle of the producer. J. Appl. Phycol. 2010, 22, 517–524. [Google Scholar] [CrossRef]
- Vítová, M.; Bišová, K.; Umysová, D.; Hlavová, M.; Kawano, S.; Zachleder, V.; Cížková, M. Chlamydomonas reinhardtii: Duration of its cell cycle and phases at growth rates affected by light intensity. Planta 2011, 233, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Bisova, K.; Zachleder, V. Cell-cycle regulation in green algae dividing by multiple fission. J. Exp. Bot. 2014, 65, 2585–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kacew, S. Confounding factors in toxicity testing. Toxicology 2001, 160, 87–96. [Google Scholar] [CrossRef]
- OECD. Test Guidelines for the Chemicals; OECD Publishing: Paris, France, 1984–2017.
- Chisholm, S.W.; Brand, L.E. Persistence of cell division phasing in marine phytoplankton in continuous light after entrainment to light: Dark cycles. J. Exp. Mar. Biol. Ecol. 1981, 51, 107–118. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Banfalvi, G. Overview of Cell Synchronization. In Cell Cycle Synchronization; Banfalvi, G., Ed.; Humana Press: Totowa, NJ, USA, 2011. [Google Scholar]
- Zones, J.M.; Blaby, I.K.; Merchant, S.S.; Umen, J.G. High-Resolution Profiling of a Synchronized Diurnal Transcriptome from Chlamydomonas reinhardtii Reveals Continuous Cell and Metabolic Differentiation. Plant Cell 2015, 27, 2743–2769. [Google Scholar] [PubMed]
- Mitchell, M.C.; Meyer, M.T.; Griffiths, H. Dynamics of carbon-concentrating mechanism induction and protein relocalization during the dark-to-light transition in synchronized Chlamydomonas reinhardtii. Plant Physiol. 2014, 166, 1073–1082. [Google Scholar] [CrossRef]
- Garz, A.; Sandmann, M.; Rading, M.; Ramm, S.; Menzel, R.; Steup, M. Cell-to-cell diversity in a synchronized chlamydomonas culture as revealed by single-cell analyses. Biophys. J. 2012, 103, 1078–1086. [Google Scholar] [CrossRef]
- Ehara, T.; Osafune, T.; Hase, E. Behavior of mitochondria in synchronized cells of Chlamydomonas reinhardtii (Chlorophyta). J. Cell Sci. 1995, 108, 499–507. [Google Scholar]
- Sorokin, C. Time course of oxygen evolution during photosynthesis in synchronized cultures of algae. Plant Physiol. 1961, 36, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Hlavová, M.; Vítová, M.; Bišová, K. Synchronization of Green Algae by Light and Dark Regimes for Cell Cycle and Cell Division Studies. In Plant Cell Division; Humana Press: New York, NY, USA, 2016; pp. 3–16. [Google Scholar]
- Jüppner, J.; Mubeen, U.; Leisse, A.; Caldana, C.; Brust, H.; Steup, M.; Herrmann, M.; Steinhauser, D.; Giavalisco, P. Dynamics of lipids and metabolites during the cell cycle of Chlamydomonas reinhardtii. Plant J. 2017, 92, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Kalucka, J.; Missiaen, R.; Georgiadou, M.; Schoors, S.; Lange, C.; De Bock, K.; Dewerchin, M.; Carmeliet, P. Metabolic control of the cell cycle. Cell Cycle 2015, 14, 3379–3388. [Google Scholar] [CrossRef] [Green Version]
- Sperber, A.M.; Herman, J.K. Metabolism shapes the cell. J. Bacteriol. 2017, 199, e00039. [Google Scholar] [CrossRef] [PubMed]
- Grigorov, M.G. Analysis of Time Course Omics Datasets. In Bioinformatic for Omics Data; Mayer, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Spies, D.; Ciaudo, C. Dynamics in Transcriptomics: Advancements in RNA-seq Time Course and Downstream Analysis. Comput. Struct. Biotechnol. J. 2015, 13, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Borgert, C.J.; Wise, K.; Becker, R. A Modernizing problem formulation for risk assessment necessitates articulation of mode of action. Regul. Toxicol. Pharmacol. 2015, 72, 538–551. [Google Scholar] [CrossRef]
- OECD. Users’ Handbook supplement to the Guidance Document for developing and accessing. In Adverse Outcome Pathways; OECD: Paris, France, 2018. [Google Scholar]
- Bisova, K.; Krylov, D.M.; Umen, J.G. Genome-Wide Annotation and Expression Profiling of Cell Cycle Regulatory Genes in. Society 2005, 137, 475–491. [Google Scholar]
- Shene, C.; Asenjo, J.A.; Chisti, Y. Metabolic modelling and simulation of the light and dark metabolism of Chlamydomonas reinhardtii. Plant J. 2018, 96, 1076–1088. [Google Scholar] [CrossRef] [PubMed]
- Johnson, X.; Alric, J. Central Carbon Metabolism and Electron Transport in Chlamydomonas reinhardtii: Metabolic Constraints for Carbon Partitioning between Oil and Starch. Eukaryot. Cell 2013, 12, 776–793. [Google Scholar] [CrossRef] [PubMed]
- Peltier, G.; Thibault, P. O2-Uptake in the Light in Chlamydomonas: Evidence for Persistent Mitochondrial Respiration. Plant Physiol. 1985, 79, 225–230. [Google Scholar] [CrossRef]
- Brack, W.; Rottler, H. Toxicity testing of highly volatile chemicals with green algae. Environ. Sci. Pollut. Res. 1994, 1, 223–228. [Google Scholar] [CrossRef]
- Galassi, S.; Vighi, M. Testing toxicity of volatile substances with algae. Chemosphere 1981, 10, 1123–1126. [Google Scholar] [CrossRef]
- Hailing-Sørensen, B.; Nyhohn, N.; Baun, A. Algal toxicity tests with volatile and hazardous compounds in air-tight test flasks with CO2 enriched headspace. Chemosphere 1996, 32, 1513–1526. [Google Scholar] [CrossRef]
- Lin, J.H.; Kao, W.C.; Tsai, K.P.; Chen, C.Y. A novel algal toxicity testing technique for assessing the toxicity of both metallic and organic toxicants. Water Res. 2005, 39, 1869–1877. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.J.; Chen, C.Y. Toxicity assessment of pesticides to Pseudokirchneriella subcapitata under air-tight test environment. J. Hazard. Mater. 2006, 131, 6–12. [Google Scholar] [CrossRef]
- Chen, C.Y.; Yan, Y.K.; Yang, C.F. Toxicity assessment of polycyclic aromatic hydrocarbons using an air-tight algal toxicity test. Water Sci. Technol. 2006, 54, 309. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.P.; Chen, C.Y. An algal toxicity database of organic toxicants derived by a closed-system technique. Environ. Toxicol. Chem. 2007, 26, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Mayer, P.; Nyholm, N.; Verbruggen, E.M.J.; Hermens, J.L.M.; Tolls, J. Algal growth inhibition test in filled, closed bottles for volatile and sorptive materials. Environ. Toxicol. Chem. 2000, 19, 2551. [Google Scholar] [CrossRef]
- Herman, D.C.; Inniss, W.E.; Mayfield, C.I. Impact of volatile aromatic hydrocarbons, alone and in combination, on growth of the freshwater alga Selenastrum capricornutum. Aquat. Toxicol. 1990, 18, 87–100. [Google Scholar] [CrossRef]
- Fairchild, J.F.; Ruessler, D.S.; Carlson, A.R. Comparative Sensitivity of Five Species of Macrophytes and Six Species of Algae to Atrazine, Metribuzin, Alachlor, and Metolachlor. Environ. Toxicol. Chem. 1998, 17, 1830–1834. [Google Scholar] [CrossRef]
- Shurong, D.; Fengfen, Z.; Min, Z. Toxicity of Fenvalerate to Six Species of Fish and Two Species of Fishfood Organisms; Ryans, R.C., Ed.; United States Environmental Protection Agency: Guangzhou, China, 1988; pp. 284–298.
- Hsieh, S.H.; Tsai, K.P.; Chen, C.Y. The combined toxic effects of nonpolar narcotic chemicals to Pseudokirchneriella subcapitata. Water Res. 2006, 40, 1957–1964. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Fiehn, O. High quality metabolomic data for Chlamydomonas reinhardtii. Plant Methods 2008, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Song, D.; Kind, T.; Ma, Y.; Hoefkens, J.; Fiehn, O. Lipidomic analysis of Chlamydomonas reinhardtii under nitrogen and sulfur deprivation. PLoS ONE 2015, 10, e0137948. [Google Scholar] [CrossRef] [PubMed]
- Schmollinger, S.; Muhlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing Mechanisms in Chlamydomonas Affect the Transcriptome, the Proteome, and Photosynthetic Metabolism. Plant Cell 2014, 26, 1410–1435. [Google Scholar] [CrossRef]
- Lv, H.; Qu, G.; Qi, X.; Lu, L.; Tian, C.; Ma, Y. Transcriptome analysis of Chlamydomonas reinhardtii during the process of lipid accumulation. Genomics 2013, 101, 229–237. [Google Scholar] [CrossRef]
- Fang, W.; Si, Y.; Douglass, S.; Casero, D.; Merchant, S.S.; Pellegrini, M.; Ladunga, I.; Liu, P.; Spalding, M.H. Transcriptome-Wide Changes in Chlamydomonas reinhardtii Gene Expression Regulated by Carbon Dioxide and the CO2-Concentrating Mechanism Regulator CIA5/CCM1. Plant Cell 2012, 24, 1876–1893. [Google Scholar] [CrossRef]
- Wienkoop, S.; Weiss, J.; May, P.; Kempa, S.; Irgang, S.; Recuenco-Munoz, L.; Pietzke, M.; Schwemmer, T.; Rupprecht, J.; Egelhofer, V.; et al. Targeted proteomics for Chlamydomonas reinhardtii combined with rapid subcellular protein fractionation, metabolomics and metabolic flux analyses. Mol. Biosyst. 2010, 6, 1018–1031. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Dinney, C.P.; Shete, S.; Huang, M.; Hildebrandt, M.A.; Chen, Z.; Gu, J. Comprehensive pathway-based interrogation of genetic variations in the nucleotide excision DNA repair pathway and risk of bladder cancer. Cancer 2012, 118, 205–215. [Google Scholar] [CrossRef]
- Esperanza, M.; Seoane, M.; Rioboo, C.; Herrero, C.; Cid, Á. Chlamydomonas reinhardtii cells adjust the metabolism to maintain viability in response to atrazine stress. Aquat. Toxicol. 2015, 165, 64–72. [Google Scholar] [CrossRef]
- USEPA. ECOTOXicology Knowledgebase (ECOTOX); USEPA: Washington, DC, USA, 2019.
- USEPA. Ambient Water Quality Criteria for Chlorinated Benzenes; Report EPA 440/5-80-028; USEPA: Washington, DC, USA, 1980.
- Bedient, P.B.; Rifai, H.S.; Newell, C.J. Ground Water Contamination: Transport and Remediation, 2nd ed.; Prentice Hall PTR: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- USEPA. CERCLIS3. Available online: https://iaspub.epa.gov/sor_internet/registry/substreg/substance/details.do?displayPopup=&id=83723 (accessed on 9 May 2019).
- Puzanskiy, R.; Tarakhovskaya, E.; Shavarda, A.; Shishova, M. Metabolomic and physiological changes of Chlamydomonas reinhardtii (Chlorophyceae, Chlorophyta) during batch culture development. J. Appl. Phycol. 2017, 30, 808–818. [Google Scholar] [CrossRef]
- Altenburger, R.; Schmitt-Jansen, M.; Riedl, J. Bioassays with Unicellular Algae: Deviations from Exponential Growth and Its Implications for Toxicity Test Results. J. Environ. Qual. 2008, 37, 16. [Google Scholar] [CrossRef] [PubMed]
- Janssen, C.R.; Heijerick, D.G. Algal Toxicity Tests for Environmental Risk Assessments of Metals. In Reviews of Environmental Contamination and Toxicology; Springer Nature: Basingstoke, UK, 2003; pp. 23–52. [Google Scholar]
- Hörnström, E. Toxicity test with algae—A discussion on the batch method. Ecotoxicol. Environ. Saf. 1990, 20, 343–353. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MSEA: A web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res. 2010, 38, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.C.; Weljie, A.M.; Turner, R.J. Computational tools for the secondary analysis of metabolomics experiments. Comput. Struct. Biotechnol. J. 2013, 4, e201301003. [Google Scholar] [CrossRef]
- Rosato, A.; Tenori, L.; Cascante, M.; De Atauri Carulla, P.R.; Martins dos Santos, V.A.P.; Saccenti, E. From correlation to causation: Analysis of metabolomics data using systems biology approaches. Metabolomics 2018, 14, 37. [Google Scholar] [CrossRef]
- Chagoyen, M.; Pazos, F. MBRole: Enrichment analysis of metabolomic data. Bioinformatics 2011, 27, 730–731. [Google Scholar] [CrossRef]
- Sueoka, N. Mitotic Replication of Deoxyribonucleic Acid in Chlamydomonas reinhardi. Proc. Natl. Acad. Sci. USA 1960, 46, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Kropat, J.; Hong-Hermesdorf, A.; Casero, D.; Ent, P.; Castruita, M.; Pellegrini, M.; Merchant, S.S.; Malasarn, D. A revised mineral nutrient supplement increases biomass and growth rate in Chlamydomonas reinhardtii. Plant J. 2011, 66, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Renberg, L.; Johansson, A.I.; Shutova, T.; Stenlund, H.; Aksmann, A.; Raven, J.A.; Gardeström, P.; Moritz, T.; Samuelsson, G. A metabolomic approach to study major metabolite changes during acclimation to limiting CO2 in Chlamydomonas reinhardtii. Plant Physiol. 2010, 154, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, P.; Portais, J.C.; Vorholt, J.A. Quantitative metabolome analysis using liquid chromatography-high-resolution mass spectrometry. Anal. Biochem. 2008, 382, 94–100. [Google Scholar] [CrossRef]
- Iwasa, K.; Murakami, S. Palmelloid formation of Chlamydomonas II. Mechanism of palmelloid formation by organic acids. Physiol. Plant 1969, 22, 43–50. [Google Scholar] [CrossRef]
- Messerli, M.A.; Amaral-Zettler, L.A.; Zettler, E.; Jung, S.K.; Smith, P.J.S.; Sogin, M.L. Life at acidic pH imposes an increased energetic cost for a eukaryotic acidophile. J. Exp. Biol. 2005, 208, 2569–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küsel, A.C.; Sianoudis, J.; Leibfritz, D.; Grimme, L.H.; Mayer, A. The dependence of the cytoplasmic pH in aerobic and anaerobic cells of the green algae Chlorella fusca and Chlorella vulgaris on the pH of the medium as determined by 31P in vivo NMR spectroscopy. Arch. Microbiol. 1990, 153, 254–258. [Google Scholar] [CrossRef]
- Arensberg, P.; Hemmingsen, V.H.; Nyholm, N. A miniscale algal toxicity test. Chemosphere 1995, 30, 2103–2115. [Google Scholar] [CrossRef]
- Jutson, M.G.S.; Pipe, R.K.; Thomas, C.R. The Cultivation of Marine Phytoplankton; Tsaloglou, M.N., Ed.; Caister Acadmic Press: Poole, UK, 2016; pp. 11–26. [Google Scholar]
- Rowan, B.A.; Oldenburg, D.J.; Bendich, A.J. A high-throughput method for detection of DNA in chloroplasts using flow cytometry. Plant Methods 2007, 3, 5. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extration And Purification. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic Quotient Normalization as Robust Method to Account for Dilution of Complex Biological Mixtures. Application in 1 H NMR Metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples Analysed by PCA (All Three Batches) | PC1 | PC2 | PC3 |
|---|---|---|---|
| Controls only | p = 7.15 × 10−6 | p = 0.11 | p = 3.84 × 10−4 |
| CB only | p = 1.72 × 10−7 | p = 0.20 | p = 0.048 |
| CB normalized to batch-specific controls | p = 0.20 | p = 0.015 | p = 0.11 |
| Metabolite Set | Metric | Batch 1 | Batch 2 | Batch 3 |
|---|---|---|---|---|
| SB1 | NES | - | 1.39 | 1.33 |
| LE | - | 11 features | 72 features | |
| FDR | - | 0.004 | 0.006 | |
| SB2 | NES | 1.41 | - | 1.96 |
| LE | 53 features | - | 43 features | |
| FDR | <0.001 | - | <0.001 | |
| SB3 | NES | 1.38 | 2.02 | - |
| LE | 61 features | 50 features | - | |
| FDR | <0.001 | <0.001 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schade, S.; Butler, E.; Gutsell, S.; Hodges, G.; Colbourne, J.K.; Viant, M.R. Improved Algal Toxicity Test System for Robust Omics-Driven Mode-of-Action Discovery in Chlamydomonas reinhardtii. Metabolites 2019, 9, 94. https://doi.org/10.3390/metabo9050094
Schade S, Butler E, Gutsell S, Hodges G, Colbourne JK, Viant MR. Improved Algal Toxicity Test System for Robust Omics-Driven Mode-of-Action Discovery in Chlamydomonas reinhardtii. Metabolites. 2019; 9(5):94. https://doi.org/10.3390/metabo9050094
Chicago/Turabian StyleSchade, Stefan, Emma Butler, Steve Gutsell, Geoff Hodges, John K. Colbourne, and Mark R. Viant. 2019. "Improved Algal Toxicity Test System for Robust Omics-Driven Mode-of-Action Discovery in Chlamydomonas reinhardtii" Metabolites 9, no. 5: 94. https://doi.org/10.3390/metabo9050094