Placenta, Pericarp, and Seeds of Tabasco Chili Pepper Fruits Show a Contrasting Diversity of Bioactive Metabolites



Abstract
:1. Introduction
2. Results
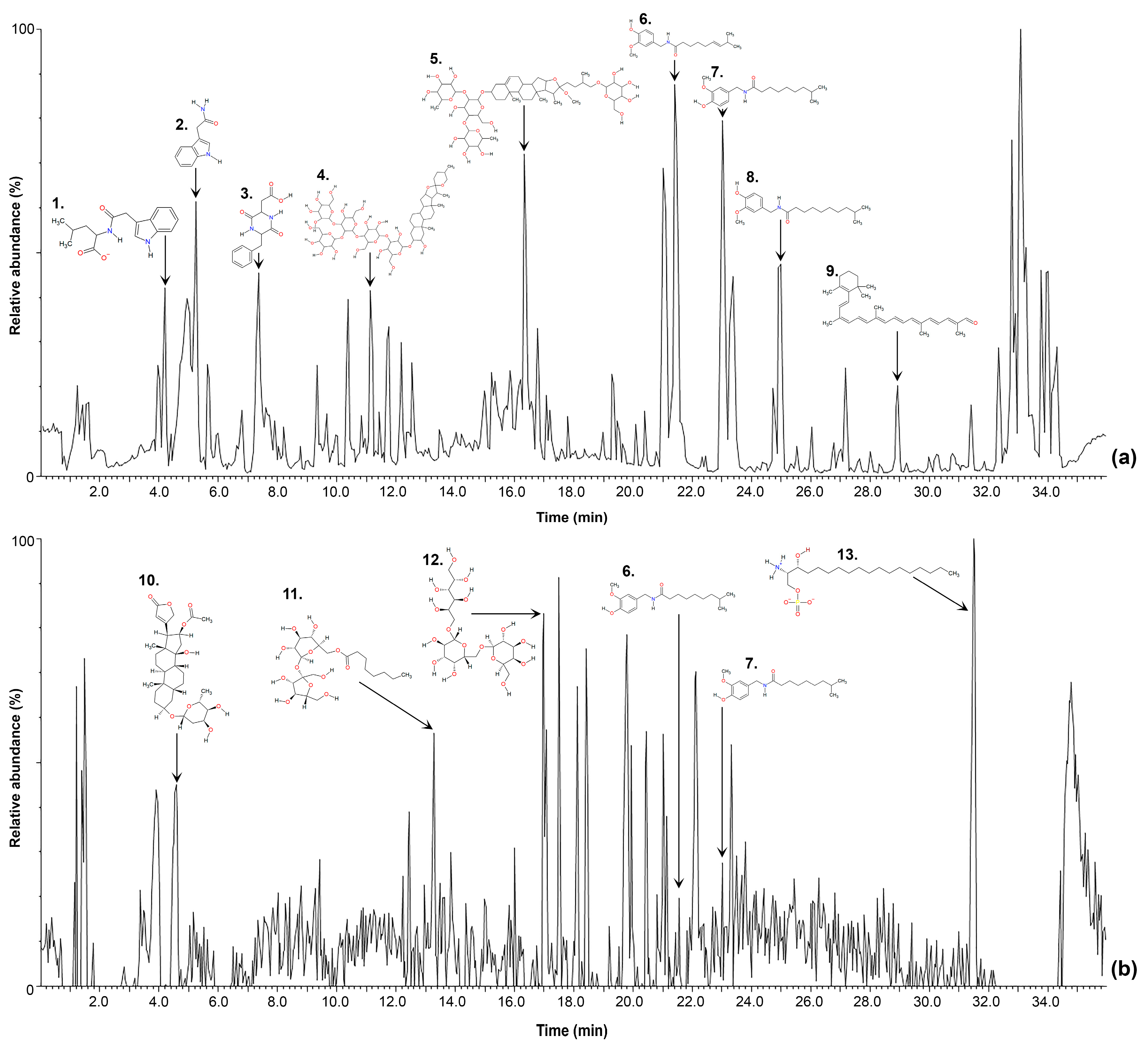
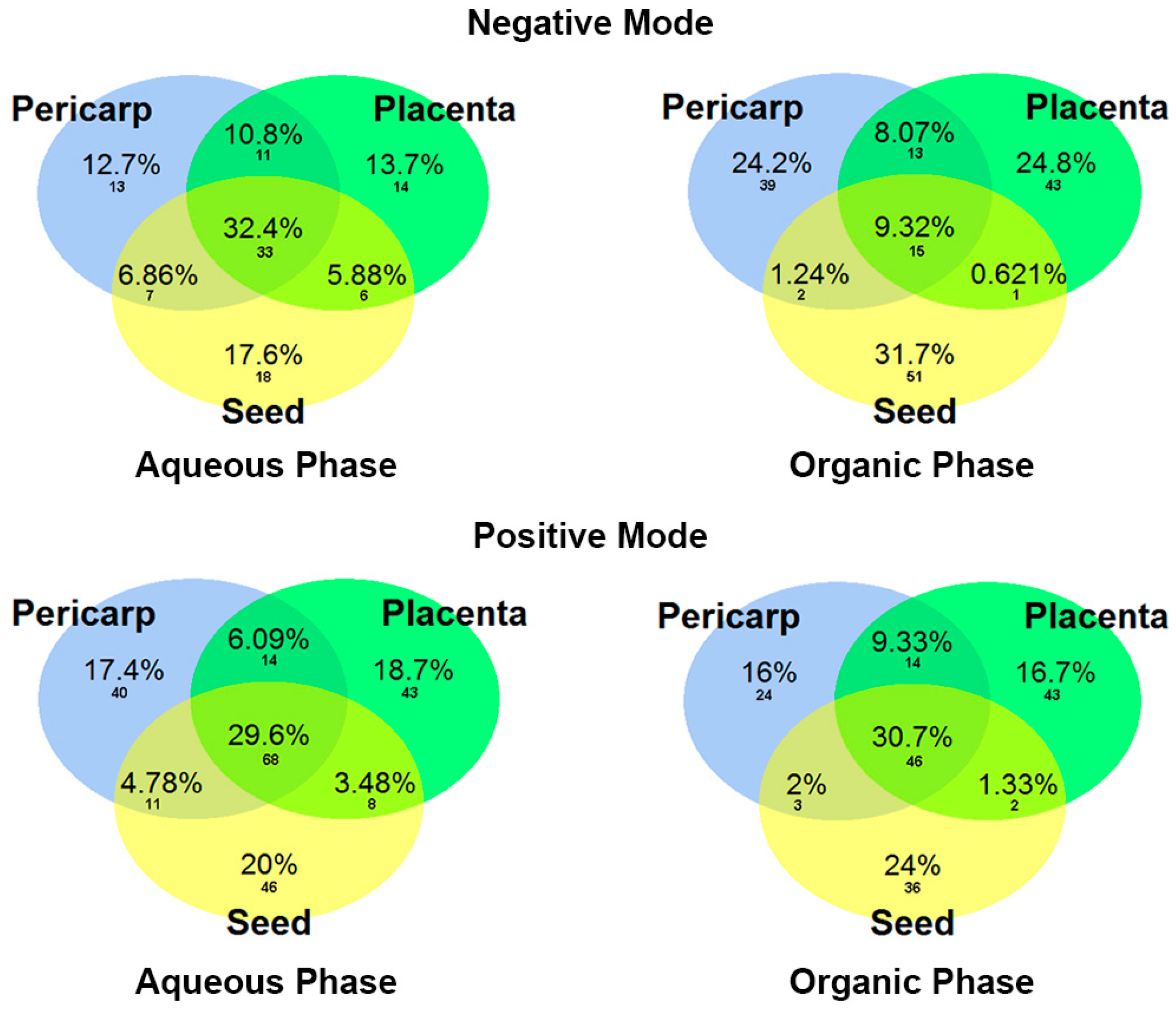
2.1. Non-Targeted Metabolomic Analysis
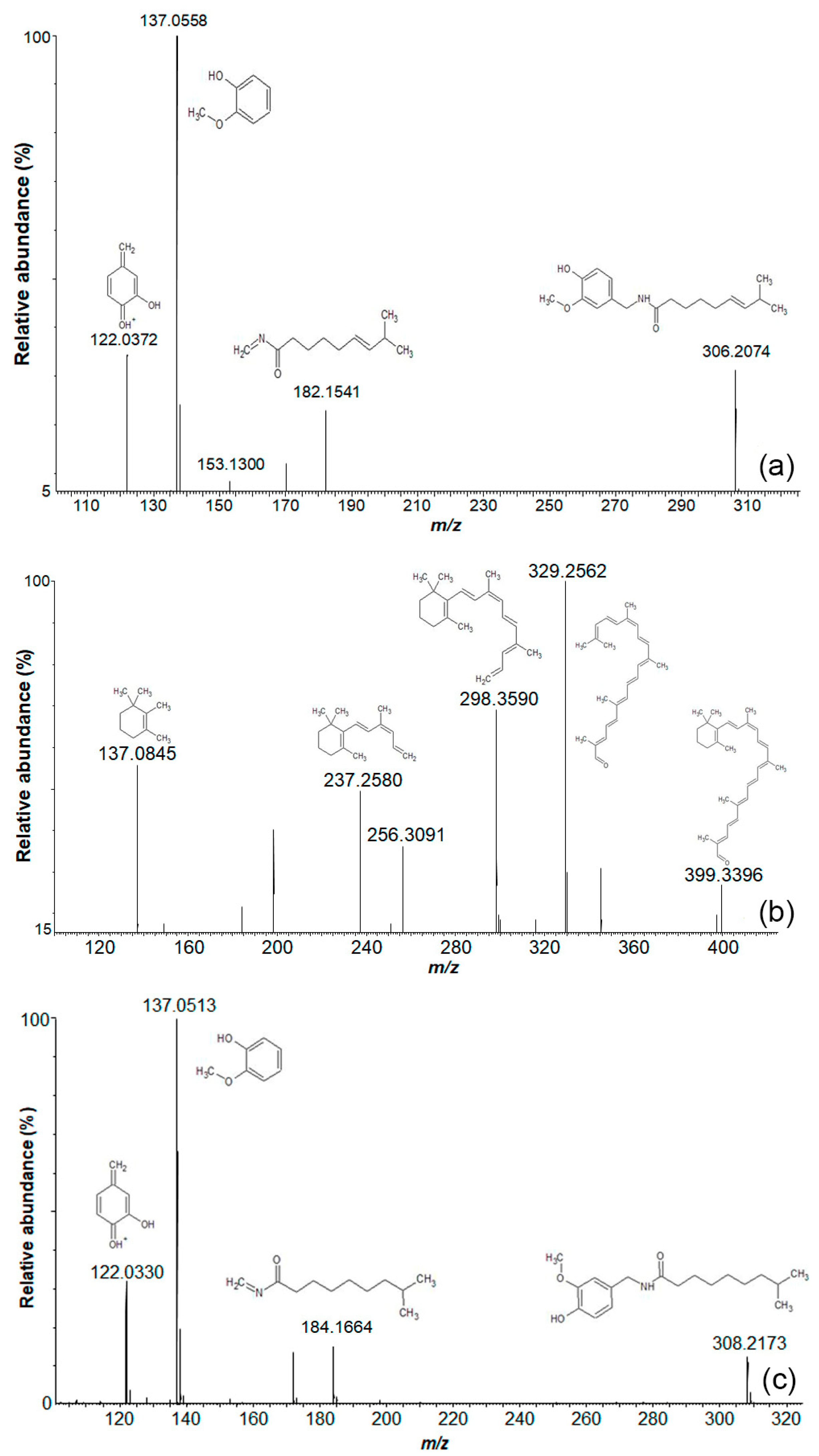
2.2. Level 1 and 2 Metabolomic Identification Analysis
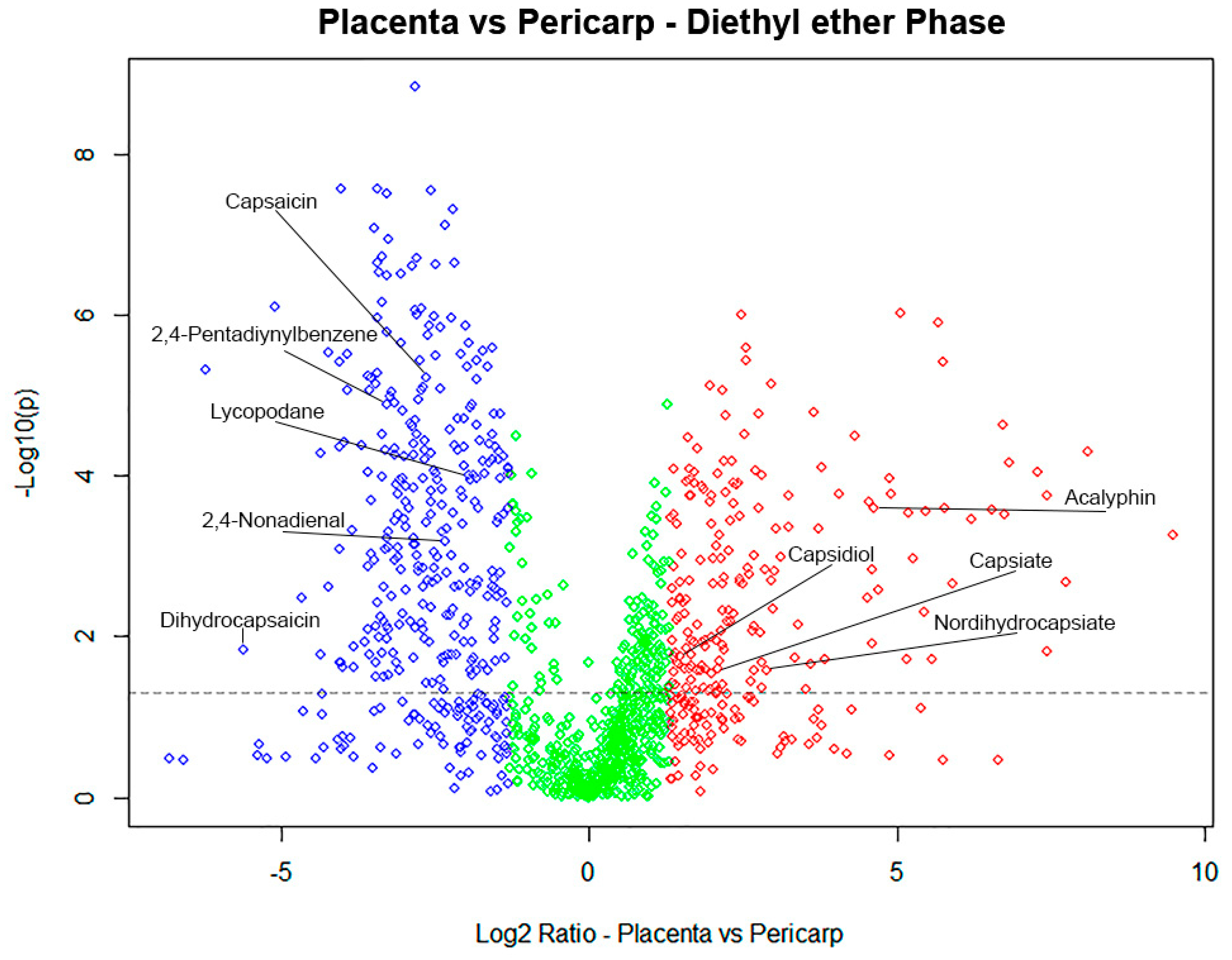
2.3. Different Metabolomic Profiles in Capsicum Sections
3. Discussion
4. Materials and Methods
4.1. Plant Material and Dissection of Tissues and Seed
4.2. Chemicals, Reagents, and Standards
4.3. Sample Extraction and UHPLC-MS Analysis
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guzman, I.; Bosland, P.W.; O’Connell, M.A. Heat, Color, and Flavor Compounds in Capsicum Fruit. In The Biological Activity of Phytochemicals; Springer: New York, NY, USA, 2011; pp. 109–126. [Google Scholar] [CrossRef]
- Bebeli, P.J.; Mazzucato, A. The Solanaceae—A Review of Recent Research on Genetic Resources and Advances in the Breeding of Tomato, Pepper and Eggplant. Eur. J. Plant Sci. Biotechnol. 2008, 2, 3–30. [Google Scholar]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 2 November 2018).
- Velázquez-Ventura, J.C.; de la Cruz-Lázaro, E.; Osorio-Osorio, R.; Preciado-Rangel, P. Morphological Variation of Wild Peppers (Capsicum spp.) from the State of Tabasco, Mexico. Emir. J. Food Agric. 2018, 30, 115–121. [Google Scholar] [CrossRef]
- Hayano-Kanashiro, C.; Gámez-Meza, N.; Medina-Juárez, L.Á. Wild Pepper Capsicum annuum L. var. glabriusculum: Taxonomy, Plant Morphology, Distribution, Genetic Diversity, Genome Sequencing, and Phytochemical Compounds. Crop Sci. 2016, 56, 1–11. [Google Scholar] [CrossRef]
- Ritota, M.; Marini, F.; Sequi, P.; Valentini, M. Metabolomic Characterization of Italian Sweet Pepper (Capsicum annum L.) by Means of HRMAS-NMR Spectroscopy and Multivariate Analysis. J. Agric. Food Chem. 2010, 58, 9675–9684. [Google Scholar] [CrossRef] [PubMed]
- Marín, A.; Ferreres, F.; Tomás-Barberán, F.A.; Gil, M.I. Characterization and Quantitation of Antioxidant Constituents of Sweet Pepper (Capsicum annuum L.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef]
- Fayos, O.; Savirón, M.; Orduna, J.; Barbero, G.F.; Mallor, C.; Garcés-Claver, A. Quantitation of Capsiate and Dihydrocapsiate and Tentative Identification of Minor Capsinoids in Pepper Fruits (Capsicum spp.) by HPLC-ESI-MS/MS(QTOF). Food Chem. 2019, 270, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Ananthan, R.; Subhash, K.; Longvah, T. Capsaicinoids, Amino Acid and Fatty Acid Profiles in Different Fruit Components of the World Hottest Naga King Chilli (Capsicum chinense Jacq). Food Chem. 2018, 238, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Aranha, B.C.; Hoffmann, J.F.; Barbieri, R.L.; Rombaldi, C.V.; Chaves, F.C. Untargeted Metabolomic Analysis of Capsicum spp. by GC-MS. Phytochem. Anal. 2017, 28, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Fayos, O.; De Aguiar, A.C.; Jiménez-Cantizano, A.; Ferreiro-González, M.; Garcés-Claver, A.; Martínez, J.; Mallor, C.; Ruiz-Rodríguez, A.; Palma, M.; Barroso, C.G.; et al. Ontogenetic Variation of Individual and Total Capsaicinoids in Malagueta Peppers (Capsicum frutescens) during Fruit Maturation. Molecules 2017, 22, 736. [Google Scholar] [CrossRef]
- Kumar, O.A.; Tata, S.S. Ascorbic Acid Contents in Chili Peppers (Capsicum L.). Not. Sci. Biol. 2009, 1, 50–52. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; Menichini, F.; Tundis, R. Evaluation of Chemical Profile and Antioxidant Activity of Twenty Cultivars from Capsicum annuum, Capsicum baccatum, Capsicum chacoense and Capsicum chinense: A Comparison between Fresh and Processed Peppers. LWT-Food Sci. Technol. 2015, 64, 623–631. [Google Scholar] [CrossRef]
- Nagy, Z.; Daood, H.; Koncsek, A.; Molnár, H.; Helyes, L. The Simultaneous Determination of Capsaicinoids, Tocopherols, and Carotenoids in Pungent Pepper Powder. J. Liq. Chromatogr. Relat. Technol. 2017, 40, 199–209. [Google Scholar] [CrossRef]
- Kobata, K.; Tate, H.; Iwasaki, Y.; Tanaka, Y.; Ohtsu, K.; Yazawa, S.; Watanabe, T. Isolation of Coniferyl Esters from Capsicum baccatum L., and Their Enzymatic Preparation and Agonist Activity for TRPV1. Phytochemistry 2008, 69, 1179–1184. [Google Scholar] [CrossRef]
- Huang, W.; Cheang, W.S.; Wang, X.; Lei, L.; Liu, Y.; Ma, K.Y.; Zheng, F.; Huang, Y.; Chen, Z.-Y. Capsaicinoids but Not Their Analogue Capsinoids Lower Plasma Cholesterol and Possess Beneficial Vascular Activity. J. Agric. Food Chem. 2014, 62. [Google Scholar] [CrossRef] [PubMed]
- Iorizzi, M.; Lanzotti, V.; De Marino, S.; Zollo, F.; Blanco-Molina, M.; Macho, A.; Muñoz, E. New Glycosides from Capsicum annuum L. Var. acuminatum. Isolation, Structure Determination, and Biological Activity. J. Agric. Food Chem. 2001, 49, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Spiller, F.; Alves, M.K.; Vieira, S.M.; Carvalho, T.A.; Leite, C.E.; Lunardelli, A.; Poloni, J.A.; Cunha, F.Q.; de Oliveira, J.R. Anti-Inflammatory Effects of Red Pepper (Capsicum baccatum) on Carrageenan- and Antigen-Induced Inflammation. J. Pharm. Pharmacol. 2008, 60, 473–478. [Google Scholar] [CrossRef]
- Anandakumar, P.; Kamaraj, S.; Jagan, S.; Ramakrishnan, G.; Devaki, T. Capsaicin Provokes Apoptosis and Restricts Benzo(a)Pyrene Induced Lung Tumorigenesis in Swiss Albino Mice. Int. Immunopharmacol. 2013, 17, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Jolayemi, A.; Ojewole, J. Comparative Anti-Inflammatory Properties of Capsaicin and Ethyl Acetate Extract of Capsicum frutescens Linn [Solanaceae] in Rats. Afr. Health Sci. 2013, 13, 357–361. [Google Scholar] [CrossRef]
- M, S.; Gaur, R.; Sharma, V.; Chhapekar, S.S.; Das, J.; Kumar, A.; Yadava, S.K.; Nitin, M.; Brahma, V.; Abraham, S.K.; et al. Comparative Analysis of Fruit Metabolites and Pungency Candidate Genes Expression between Bhut Jolokia and Other Capsicum Species. PLoS ONE 2016, 11, e0167791. [Google Scholar] [CrossRef]
- Varghese, S.; Kubatka, P.; Rodrigo, L.; Gazdikova, K.; Caprnda, M.; Fedotova, J.; Zulli, A.; Kruzliak, P.; Büsselberg, D. Chili Pepper as a Body Weight-Loss Food. Int. J. Food Sci. Nutr. 2017, 68, 392–401. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant Activity and Phenolic Compounds of 112 Traditional Chinese Medicinal Plants Associated with Anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.K.; Higo, T.; Hunter, W.L.; Burt, H.M. The Antioxidants Curcumin and Quercetin Inhibit Inflammatory Processes Associated with Arthritis. Inflamm. Res. 2006, 55, 168–175. [Google Scholar] [CrossRef]
- Liu, R.H. Potential Synergy of Phytochemicals in Cancer Prevention: Mechanism of Action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite Biodiversity in Pepper (Capsicum) Fruits of Thirty-Two Diverse Accessions: Variation in Health-Related Compounds and Implications for Breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Becerra, R.; Ramírez-Chávez, E.; Molina-Torres, J.; Winkler, R. MSI.R Scripts Reveal Volatile and Semi-Volatile Features in Low-Temperature Plasma Mass Spectrometry Imaging (LTP-MSI) of Chilli (Capsicum annuum). Anal. Bioanal. Chem. 2015, 407, 5673–5684. [Google Scholar] [CrossRef] [PubMed]
- Nugroho, L.H. Red Pepper (Capsicum spp.) Fruit: A Model for the Study of Secondary Metabolite Product Distribution and Its Management. AIP Conf. Proc. 2016, 1744, 020034. [Google Scholar] [CrossRef]
- Aza-González, C.; Herrera-Isidrón, L.; Núñez-Palenius, H.G.; Martínez De La Vega, O.; Ochoa-Alejo, N. Anthocyanin Accumulation and Expression Analysis of Biosynthesis-Related Genes during Chili Pepper Fruit Development. Biol. Plant. 2013, 57, 49–55. [Google Scholar] [CrossRef]
- Materska, M. Bioactive Phenolics of Fresh and Freeze-Dried Sweet and Semi-Spicy Pepper Fruits (Capsicum annuum L.). J. Funct. Foods 2014, 7, 269–277. [Google Scholar] [CrossRef]
- Li, B.; Bhandari, D.R.; Janfelt, C.; Römpp, A.; Spengler, B. Natural products in Glycyrrhiza glabra (licorice) rhizome imaged at the cellular level by atmospheric pressure matrix-assisted laser desorption/ionization tandem mass spectrometry imaging. Plant J. 2014, 80, 161–171. [Google Scholar] [CrossRef]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How Close Are We to Complete Annotation of Metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef]
- Kamphorst, J.J.; Lewis, I.A. Editorial Overview: Recent Innovations in the Metabolomics Revolution. Curr. Opin. Biotechnol. 2017, 43, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Frank, T.; Engel, K.-H. 8-Metabolomic Analysis of Plants and Crops. In Metabolomics in Food and Nutrition; Woodhead Publishing Limited: Sawston, UK, 2013; pp. 148–191. [Google Scholar] [CrossRef]
- Schauer, N.; Fernie, A.R. Plant Metabolomics: Towards Biological Function and Mechanism. Trends Plant Sci. 2006, 11, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Pluskal, T.; Aubry, S.; Weng, J.-K. Contribution of Untargeted Metabolomics for Future Assessment of Biotech Crops. Trends Plant Sci. 2018, 23, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Fernie, A.R.; Luo, J. Exploring the Diversity of Plant Metabolism. Trends Plant Sci. 2019, 24, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Bijttebier, S.; Van der Auwera, A.; Foubert, K.; Voorspoels, S.; Pieters, L.; Apers, S. Bridging the Gap between Comprehensive Extraction Protocols in Plant Metabolomics Studies and Method Validation. Anal. Chim. Acta 2016, 935, 136–150. [Google Scholar] [CrossRef]
- Matsuda, F.; Yonekura-Sakakibara, K.; Niida, R.; Kuromori, T.; Shinozaki, K.; Saito, K. MS/MS Spectral Tag-Based Annotation of Non-Targeted Profile of Plant Secondary Metabolites. Plant J. 2009, 57, 555–577. [Google Scholar] [CrossRef]
- Blaženović, I.; Kind, T.; Ji, J.; Fiehn, O. Software Tools and Approaches for Compound Identification of LC-MS/MS Data in Metabolomics. Metabolites 2018, 8, 31. [Google Scholar] [CrossRef]
- Smith, E.; Williamson, E.; Zloh, M.; Gibbons, S. Isopimaric Acid from Pinus nigra Shows Activity against Multidrug-Resistant and EMRSA Strains of Staphylococcus aureus. Phyther. Res. 2005, 19, 538–542. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage Reserve Accumulation in Arabidopsis: Metabolic and Developmental Control of Seed Filling. Arab. B. 2008, e0113. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Alquézar, B.; Peña, L. Fruit Aromas in Mature Fleshy Fruits as Signals of Readiness for Predation and Seed Dispersal. New Phytol. 2013, 197, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Rabara, R.C.; Tripathi, P.; Rushton, P.J. Comparative Metabolome Profile between Tobacco and Soybean Grown under Water-Stressed Conditions. BioMed Res. Int. 2017, 2017, 3065251. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Sultana, B.; Anwar, F. LC—ESI—MS/MS Based Characterization of Phenolic Components in Fruits of Two Species of Solanaceae. J. Food Sci. Technol. 2017, 55, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- López-Gresa, M.P.; Lisón, P.; Campos, L.; Rodrigo, I.; Rambla, J.L.; Granell, A.; Conejero, V.; Bellés, J.M. A Non-Targeted Metabolomics Approach Unravels the VOCs Associated with the Tomato Immune Response against Pseudomonas Syringae. Front. Plant Sci. 2017, 8, 01188. [Google Scholar] [CrossRef] [PubMed]
- Heinig, U.; Aharoni, A. Analysis of Steroidal Alkaloids and Saponins in Solanaceae Plant Extracts Using UPLC-QTOF Mass Spectrometry. In Methods in Molecular Biology; Rodríguez-Concepción, M., Ed.; Springer: New York, NY, USA, 2014; Volume 1153, pp. 171–185. [Google Scholar] [CrossRef]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular Biology of Capsaicinoid Biosynthesis in Chili Pepper (Capsicum spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Escogido, D.M.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewksbury, J.J.; Nabhan, G.P. Seed Dispersal: Directed Deterrence by Capsaicin in Chillies. Nature 2001, 412, 403–404. [Google Scholar] [CrossRef]
- Yamazaki, K.; Iwashina, T.; Kitajima, J.; Gamou, Y.; Yoshida, A.; Tannowa, T. External and Internal Flavonoids from Madagascarian Uncarina Species (Pedaliaceae). Biochem. Syst. Ecol. 2007, 35, 743–749. [Google Scholar] [CrossRef]
- Furuya, T.; Ikuta, A.; Syōno, K. Alkaloids from Callus Tissue of Papaver somniferum. Phytochemistry 1972, 11, 3041–3044. [Google Scholar] [CrossRef]
- Yoshihara, T.; Sakuma, T.; Ichihara, A. Yellow Fluorescent Stress Compounds, Pratenols A and B, from Red Clover (Trifolium pratense) Infected by Kahatiella Caulivora. Biosci. Biotechnol. Biochem. 1992, 56, 1955–1958. [Google Scholar] [CrossRef]
- Jia, S.S.; Ma, C.M.; Li, Y.H.; Hao, J.H. Glycosides of Phenolic Acid and Flavonoids from the Leaves of Glycyrrhiza uralensis Ficsh. Yao Xue Xue Bao 1992, 27, 441–444. [Google Scholar] [PubMed]
- Li, Q.; Somavat, P.; Singh, V.; Chatham, L.; Gonzalez de Mejia, E. A Comparative Study of Anthocyanin Distribution in Purple and Blue Corn Coproducts from Three Conventional Fractionation Processes. Food Chem. 2017, 231, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, Y.; Stahl-Hermes, V.; Ballester, A.R.; de Vos, R.C.H.; Voorrips, R.E.; Maharijaya, A.; Molthoff, J.; Zamora, M.V.; Sudarmonowati, E.; Arisi, A.C.M.; et al. Genetic Mapping of Semi-Polar Metabolites in Pepper Fruits (Capsicum sp.): Towards Unravelling the Molecular Regulation of Flavonoid Quantitative Trait Loci. Mol. Breed. 2014, 33, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Takayama, H.; Katakawa, K.; Kitajima, M.; Yamaguchi, K.; Aimi, N. Ten New Lycopodium Alkaloids Having the Lycopodane Skeleton Isolated from Lycopodium Serratum THUNB. Chem. Pharm. Bull. Tokyo 2003, 51, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Pappas, R.S.; Sturtz, G. Unusual Alkynes Found in the Essential Oil of Artemisia dracunculus L. Var. dracunculus from the Pacific Northwest. J. Essent. Oil Res. 2001, 13, 187–188. [Google Scholar] [CrossRef]
- Bahmani, M.; Golshahi, H.; Saki, K.; Rafieian-Kopaei, M.; Delfan, B.; Mohammadi, T. Medicinal Plants and Secondary Metabolites for Diabetes Mellitus Control. Asian Pac. J. Trop. Dis. 2014, 4, S687–S692. [Google Scholar] [CrossRef]
- Nohara, T.; Nishioka, I.; Tokubuchi, N.; Miyahara, K.; Kawasaki, T. Cinncassiol C1, a Novel Type of Diterpene from Cinnamomi Cortex. Chem. Pharm. Bull. Tokyo 1980, 28, 1969–1970. [Google Scholar] [CrossRef]
- Novillo, F.; Velasco, E.; Delgado, G. Tri- and Diterpenoids from Some Species of Euphorbiaceae. Evaluation of Their Antiinflammatory and Cytotoxic Properties. Planta Med. 2015, 81, PM_106. [Google Scholar] [CrossRef]
- Borges, R.M.; Ranganathan, Y.; Krishnan, A.; Ghara, M.; Pramanik, G. When Should Fig Fruit Produce Volatiles? Pattern in a Ripening Process. Acta Oecol. 2011, 37, 611–618. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid Extraction by Methyl-Tert-Butyl Ether for High-Throughput Lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www.R-project.org/ (accessed on 3 November 2018).
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Identification | Class | PC1 |
|---|---|---|
| Tuberoside J | SPNS | 0.2012 |
| Asparagoside B | SPNS | 0.1939 |
| Matesaponin 5 | SPNS | 0.1871 |
| Oleanolic acid 3-O-[O-β-d-glucopyranosyl-(1->4)-O-β-d-glucopyranosyl-(1->3)-O-α-l-rhamnopyranosyl- (1->2)-α -l-arabinopyranoside] | SPNS | 0.1822 |
| Capsicosin | SPNS | 0.1650 |
| PC2 | ||
| (3″-Apiosyl-6″-malonyl) astragalin | FLV | 0.1401 |
| Pratenol B | BZD | 0.0881 |
| Asparagoside B | SPNS | 0.0750 |
| Matesaponin 5 | SPNS | 0.0673 |
| Kaempferol 3-xylosylglucoside | FLV | 0.0658 |
| Putative Identification | Class | PC1 |
|---|---|---|
| Abietane | TER | 0.1450 |
| (5cis,5′cis,9cis,11′cis)-1,2,7,7′,8,8′-Hexahydro-1,2-epoxy-ψ, ψ-carotene | CARO | 0.1429 |
| Lycoperoside D | SPNS | 0.1324 |
| Phyllohydroquinone | TER | 0.1302 |
| 2-Caprylooleomyristin | GL | −0.0796 |
| PC2 | ||
| α,α′-Trehalose 6-mycolate | GL | 0.1505 |
| 2-Caprylooleomyristin | GL | 0.1226 |
| MG(14:0/0:0/0:0) | GL | −0.1284 |
| Uralenneoside | BZD | −0.0757 |
| Abietane | TER | 0.0719 |
| Compound Name | Formula | Class | Adduct | Precursor (m/z) | Fragments (m/z) |
|---|---|---|---|---|---|
| α-campholenaldehyde | C10H16O | TER | [M + H − H2O] + | 135.1180 | 109.1021(3.7) |
| Jasmolone | C11H16O2 | JASM | [M + H − 2H2O] + | 145.1027 | 133.1026(4.0), 121.1024(3.3), 107.0864(3.5) |
| 2,4-Pentadiynylbenzene | C11H8 | BZD | [2M + NH4] + | 298.1669 | 177.0684(2.8), 145.0399(1.6), 117.0428(1.9) |
| Uralenneoside | C12H14O8 | BZD | [M + H] + | 287.0755 | 287.0741(63.6), 285.0609(0.4), 257.0637(2.6), 203.0493(0.5), 153.0300(1.45), 135.0542(0.1) |
| Synephrine acetonide | C12H17NO2 | BZD | [2M + FA − H] - | 459.2565 | 208.2805(8.9) |
| Cuscohygrine | C13H24N2O | AK | [M + H] + | 225.1977 | 197.1340(5.5), 183.1184 (5.6) |
| Acalyphin | C14H20N2O9 | GC | [M + Na] + | 383.1044 | 325.0952(1.3), 299.0774(4.6), 165.0311(0.3) |
| Pratenol B | C15H12O7 | BZD | [M + H − H2O] + | 287.0546 | 153.0195(2.8), 131.0512 (2.3) |
| Lycopodane | C15H25N | AK | [M + H − 2H2O] + | 220.3782 | 184.1841(5.8) |
| Pedalitin | C16H12O7 | FLV | [M + H − H2O] + | 299.0570 | 299.0568(7.3), 165.0197(0.5) |
| Nordihydrocapsaicin | C17H27NO3 | CAPS | [M+H] + | 294.2055 | 285.2240(3.6), 257.2282(2.8), 189.1653(3.9) |
| Nerolidyl acetate | C17H28O2 | TER | [M + H − 2H2O] + | 229.1966 | 161.134 (12.7) |
| Capsaicin | C18H27NO3 | CAPS | [M + H] + | 306.2075 | 182.1559(0.2), 137.0605(15.4), 122.0371(5.8) |
| Dihydrocapsaicin | C18H29NO3 | CAPS | [M + H] + | 308.2240 | 9137.061 (5.5) |
| Artocarbene | C19H18O4 | PPN | [M + H] + | 311.1301 | 175.0771(2.5), 169.0756(3.5), 163.0764(0.9), 160.0537(0.7), 137.0614(2.2), 131.0511(2.0) |
| 1-(4-hydroxyphenyl)-7-phenyl-(6E)-6-hepten-3-ol | C19H22O2 | PPN | [M + Cl] − | 317.1345 | 131.0808 (0.3) |
| Sterculynic acid | C19H30O2 | FAT | [M + H − H2O] + | 273.2235 | 273.2220(60.9), 255.2121(53.2), 173.1339(6.3), 163.0616(4.3), 161.1336(28.8), 147.1183(8.0) |
| Kaempferol 3-O-arabinoside | C20H18O10 | FLV | [M + H − 2H2O] + | 383.0783 | 325.0730(4.0), 299.0568(7.3), 165.0197(0.5) |
| all-trans-3,4-Didehydroretinoate | C20H26O2 | PRN | [M + H] + | 281.1929 | 181.1024(21.3), 165.0731(19.1), 157.1027(23.0), 155.0870(37.3), 145.1027(27.6), 128.0636(66.9) |
| Cinncassiol C | C20H28O7 | TER | [M + H − H2O] + | 363.1781 | 332.1368(0.6), 314.1253(0.5), 222.1141(0.6), 136.0677(2.1), 135.0456(0.2), 119.0495(0.8) |
| Isopimaric acid | C20H30O2 | TER | [M + H − 2H2O] + | 285.2239 | 284.2974(0.7), 257.2282(2.8) |
| 2′-Hydroxyisoorientin | C21H20O12 | FLV | [M + H] + | 465.1051 | 303.0512 (7.4) |
| 5,7,3′-trihydroxy-3,5′-dimethoxy-2′-(3′-methylbut-2-enyl)flavone | C22H22O7 | FLV | [M + H] + | 399.1472 | 381.1379 (0.9) |
| Vestitone 7-glucoside | C22H26O9 | PPN | [M + ACN + H] + | 417.1577 | 221.0831 (2.6) |
| 6-O-Acetylaustroinulin | C22H36O4 | TER | [M + ACN + Na] + | 787.5307 | 733.4879 (7.0) |
| xi-8-Acetonyldihydrosanguinarine | C23H19NO5 | AK | [M + H − H2O] + | 372.1245 | 344.1288(0.8), 149.0352(0.3) |
| Quercetin 3-(6″-malonyl-glucoside) | C24H22O15 | FLV | [M + H] + | 551.1061 | 303.0514 (23.3) |
| 12′-apo-β-carotenal | C25H34O | TER | [2M + FA − H] − | 745.5259 | 685.5227(231.2), 539.4294(47.0) |
| Kaempferol 3-xylosylglucoside | C26H28O15 | FLV | [M + H] + | 581.1525 | 341.2486(1.6), 287.0557(49.5), 153.0195(2.8), 131.0512(2.3) |
| 11′-Carboxy-α-tocopherol | C26H42O4 | TPHE | [M + H] + | 419.3222 | 177.1023 (0.5) |
| β-tocopherol | C28H48O2 | TPHE | [2M − H] - | 831.7267 | 417.6959(90.2) |
| Amarogentin | C29H30O13 | GC | [M + NH4] + | 604.2051 | 325.073(4.0), 299.0568(7.3), 165.0197(0.5) |
| Rhamnazin 3-rutinoside | C29H34O16 | FLV | [M + 2Na − H] + | 683.1485 | 303.0514 (23.3) |
| Myrciacitrin V | C30H30O13 | FLV | [M + ACN + H] + | 640.2098 | 151.0407 (5.4) |
| Bryononic acid | C30H46O3 | CBN | [M + ACN + Na] + | 518.3645 | 358.1972(0.5), 342.2300(1.5), 320.2464(1.8), 222.1338(0.3), 196.1848(0.3) |
| 3,7-Dihydroxy-25-methoxycucurbita-5,23-dien-19-al | C31H50O4 | STR | [M + Cl] − | 521.3404 | 485.7277(21.0) |
| Capsianoside I | C32H52O14 | TER | [M + Na] + | 683.3298 | 683.3291(4.4), 365.1088(1.6), 363.0929(1.6), 271.2444(7.5) |
| Diosgenin 3-O-beta-d-glucoside | C33H52O8 | TER | [M + H] + | 577.3759 | 468.2101(67.0), 441.1756(9.8), 415.3230(23.2), 397.3135(4.5), 397.1857(3.8), 271.0622(4.8) |
| Kidjoranin-3-O-β-digitoxopyranoside | C36H48O10 | SPNS | [M − H2O − H] − | 621.3029 | 621.3011(167.6), 579.2889(8.2), 285.1144(46.0), 255.0975(5.6) |
| Feruloyl-β-sitosterol | C39H58O4 | TER | [2M + Hac − H] − | 1239.9012 | 1239.893(28.0), 887.5754(11.0) |
| Ubiquinol-6 | C39H60O4 | PRN | [M + Na] + | 615.4544 | 394.3743(20.0), 322.2779(2.0), 310.3341(3.8), 134.1078(2.0) |
| Fistuloside A | C39H62O13 | SPNS | [M + H] + | 739.4309 | 577.3766(32.5), 468.2101(67.0), 441.1756(9.8), 415.3230(23.2), 397.3135 (4.5),271.0622(4.8) |
| Nigroxanthin | C40H54O2 | TER | [M + 2Na − H] + | 611.3841 | 467.2684(1.0), 449.3285(2.3), 305.2134(4.5), 287.2032(2.6), 269.1927(1.9) |
| Ursolic acid 3-[glucosyl-(1->4)-xyloside] | C41H66O12 | TER | [M + Na] + | 773.4399 | 773.4389(4.5), 686.3793(2.1), 611.3844(5.0), 449.3285(2.3), 305.2134(4.5), 287.2032(2.6) |
| Melilotoside B | C41H68O12 | TER | [M + H] + | 753.4203 | 267.1773 (0.4) |
| Licoricesaponin C2 | C42H62O15 | TER | [M + Na] + | 829.3952 | 829.3948(32.4), 723.3561(3.4),624.3786(4.6),310.1940(1.3), 250.1564(8.8), 146.0618(1.3) |
| Tuberoside L | C51H84O23 | SPNS | [M + H] + | 1065.5627 | 670.3848(1.4), 611.3847(1.6), 449.3291(1.6), 432.3226(0.5) |
| Yamogenintetroside B | C52H86O22 | SPNS | [M + 2Na − H] + | 1107.5319 | 854.4602(7.0), 762.4256(11.1), 559.4917(1.7), 541.4820(2.5), 426.3396(5.1), 309.1197(1.9) |
| Oleanolic acid 3-O-[O-β-d-glucopyranosyl-(1->4)-O-β-d-glucopyranosyl-(1->3)-O-α-l-rhamnopyranosyl-(1->2)-α-l-arabinopyranoside] | C53H86O21 | TER | [M + Na] + | 1081.5464 | 773.4401(1.3), 611.3846(3.4), 449.3298(3.2), 153.0195(6.5) |
| Tragopogonsaponin F | C56H80O21 | SPNS | [M + CH3OH + H] + | 1121.5515 | 786.4322 (43.4) |
| Trigofoenoside G | C56H92O27 | SPNS | [M + H] + | 1197.5995 | 1197.5994(31.3), 829.3948(32.4), 723.3561(3.4), 624.3786(4.6), 338.1889(1.5), 250.1564(8.8) |
| Hovenoside D | C57H92O26 | TER | [M + CH3OH + H] + | 1225.6028 | 1210.6307(124.9), 1064.5708(55.2), 870.4542(25.2), 442.3347(11.2), 325.1173(14.4), 301.0726(19.0) |
| Capsicosin | C57H94O29 | TER | [M + H] + | 1243.6144 | 595.3883(18.9), 433.3333(18.6), 415.3237(9.2), 289.2185(18.0), 271.2091(10.8), 161.1340(12.7) |
| Eleutheroside L | C59H96O25 | SPNS | [M + Na] + | 1227.6129 | 932.493(12.4), 399.3288(8.4), 285.2599(2.2) |
| β-l-arabinose 1-phosphate(2-) | C5H9O8P-2 | GC | [M + Cl] − | 262.9693 | 262.9688(2.6), 218.9505(4.6) |
| Capsicoside A | C63H106O35 | SPNS | [M + H − 2H2O] + | 1387.6618 | 901.4882(5.1), 739.4320(24.5), 577.3766(32.5), 468.2101(67.0), 441.1756(9.8), 415.3230(23.2) |
| Matesaponin 5 | C65H106O31 | SPNS | [M + Na] + | 1405.6713 | 757.4389(195.6), 595.3839(200.6), 451.2716(56.9), 289.2162(151.2), 271.2075(66.5), 253.1970(56.8) |
| Pyridoxamine | C8H12N2O2 | PYR | [M+H−H2O] + | 151.0872 | 135.0247 (0.3) |
| 3-[3,4-Dihydroxy-2-(hydroxymethyl)-1-pyrrolidinyl]propanamide | C8H16N2O4 | AK | [M + H − H2O] + | 187.1093 | 175.1117(1.3), 155.0443(0.2), 116.0711(1.3), 112.0767(0.5), 109.0296(0.9) |
| 2,4-Nonadienal | C9H14O | CBN | [M + K] + | 177.0683 | 169.1144(11.7), 157.1133(15.3), 155.0982(17.7), 153.0822(13.1), 142.0889(25.6), 128.0724(24.6) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cervantes-Hernández, F.; Alcalá-González, P.; Martínez, O.; Ordaz-Ortiz, J.J. Placenta, Pericarp, and Seeds of Tabasco Chili Pepper Fruits Show a Contrasting Diversity of Bioactive Metabolites. Metabolites 2019, 9, 206. https://doi.org/10.3390/metabo9100206
Cervantes-Hernández F, Alcalá-González P, Martínez O, Ordaz-Ortiz JJ. Placenta, Pericarp, and Seeds of Tabasco Chili Pepper Fruits Show a Contrasting Diversity of Bioactive Metabolites. Metabolites. 2019; 9(10):206. https://doi.org/10.3390/metabo9100206
Chicago/Turabian StyleCervantes-Hernández, Felipe, Paul Alcalá-González, Octavio Martínez, and José Juan Ordaz-Ortiz. 2019. "Placenta, Pericarp, and Seeds of Tabasco Chili Pepper Fruits Show a Contrasting Diversity of Bioactive Metabolites" Metabolites 9, no. 10: 206. https://doi.org/10.3390/metabo9100206