A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria


Abstract
:1. Introduction
2. Results and Discussion
2.1. Development of a Rapid Extraction Procedure Compatible with the HPLC-APCI Analysis
2.2. Optimization of the HPLC Gradient and APCI-MS Parameters
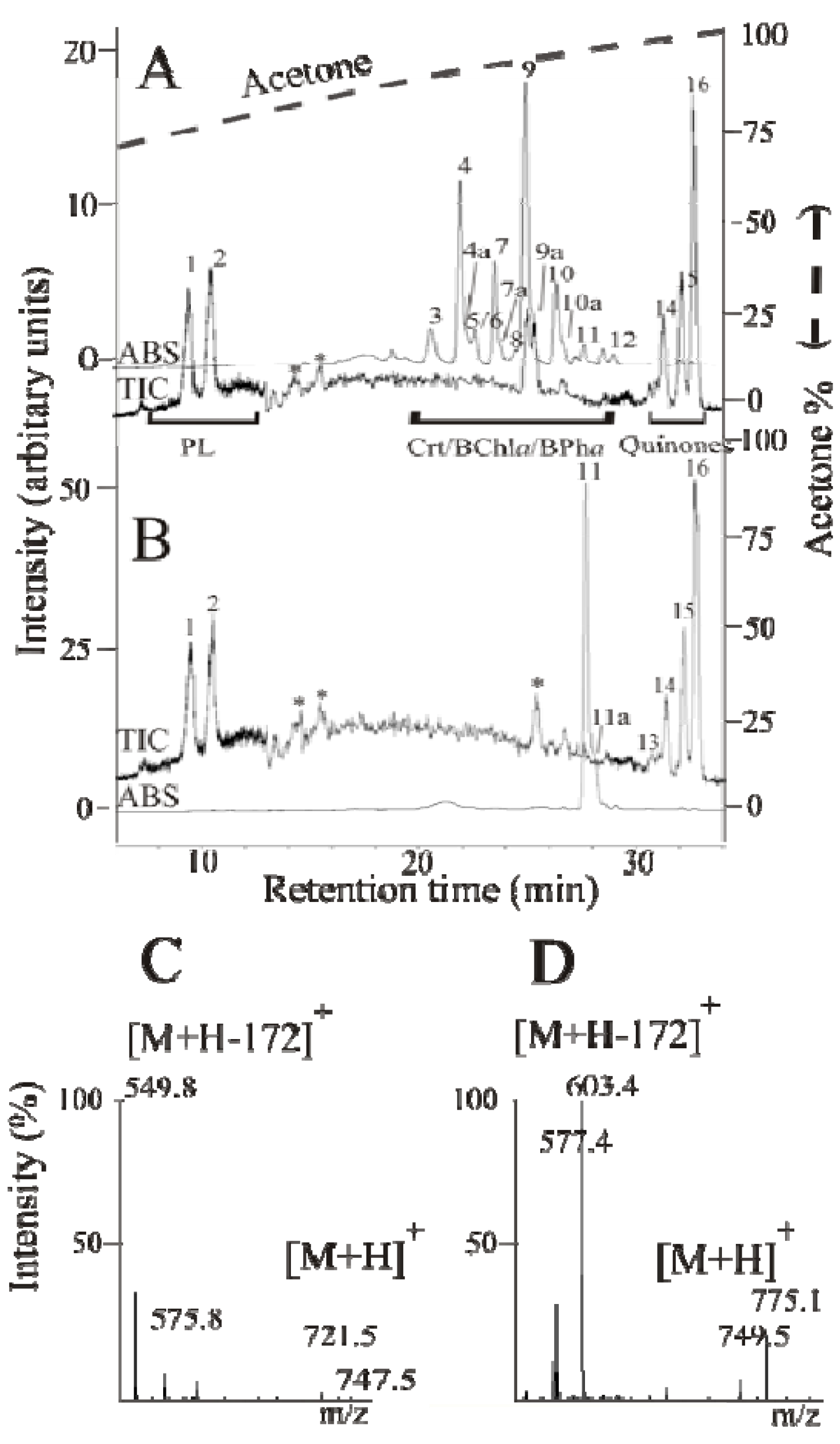
2.3. Detailed Analysis of the Carotenoids Present in the Mutant Strains ST4 and SLYC18

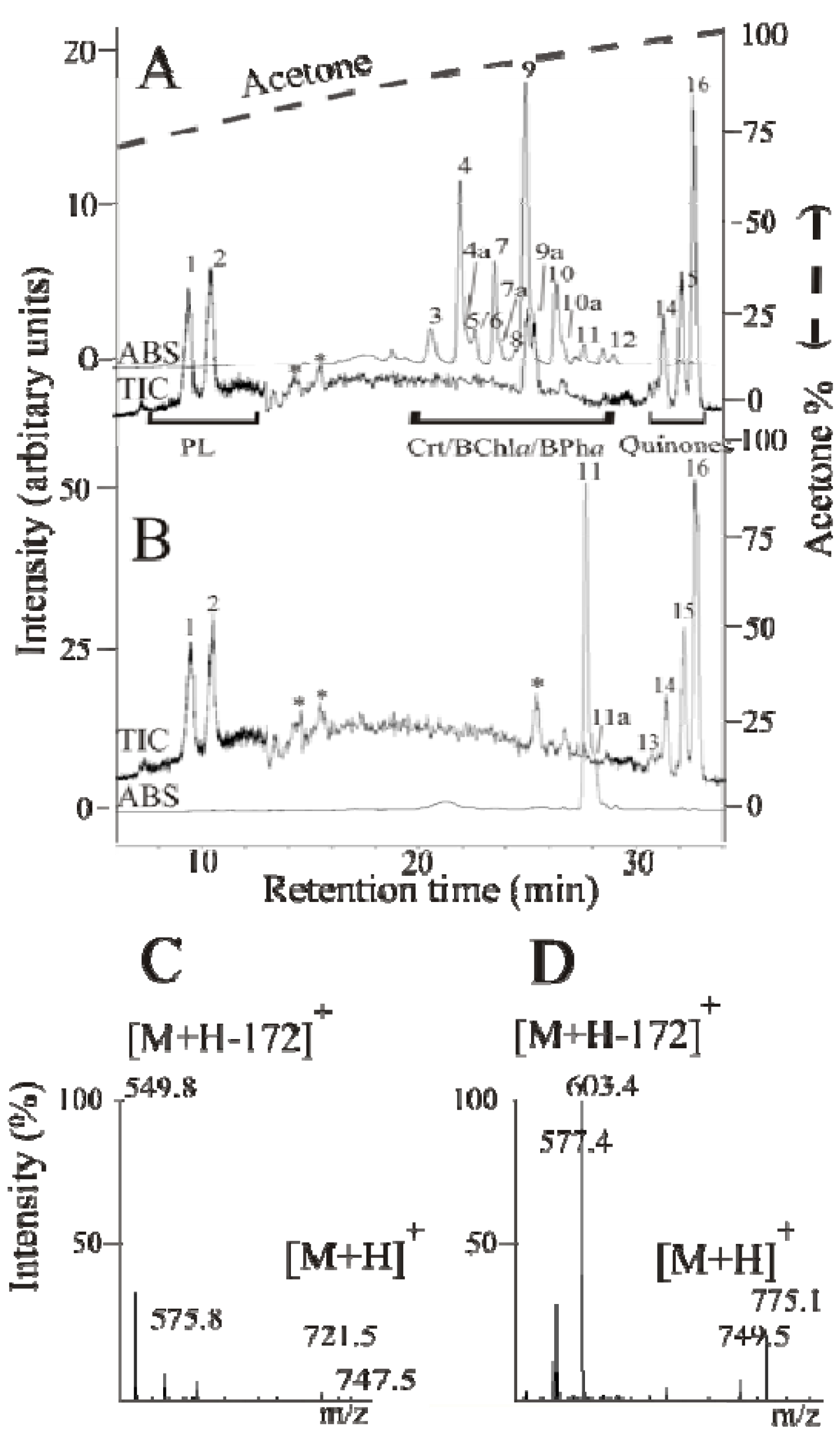{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Retention time (min) | Exact Mass (g) | Molecular ion | Molec. ion/fragment a | |
|---|---|---|---|---|---|
| Phospholipids | Palmitoyl-palmitoleyl-PG (C16:0/C16:1) (PPoPG) | 9.14 | 720.5 | [M+H-172]+ | 549.4 |
| Palmitoleyl-oleoyl-PG (C16:1/C18:1) (PoOPG) | 9.14 | 746.5 | [M+H-172]+ | 575.4 | |
| Palmitoyl-oleoyl-PG (C16:0/C18:1) (POPG) | 9.95 | 748.5 | [M+H-172]+ | 577.4 | |
| Pentadecanoyl-octadecenoyl-PG (C15:0/C18:1) | 9.95 | 734.5 | [M+H-172]+ | 563.4 | |
| Tetradecanoyl-hexadecanoyl-PG (C14:0/C16:0) | 9.95 | 694.5 | [M+H-172]+ | 523.4 | |
| Heptadecenoyl-octadecenoyl-PG (C17:1/C18:1) | 9.95 | 760.5 | [M+H-172]+ | 589.4 | |
| Octadecenoyl-nonadecenoyl-PG (C18:1/C19:1) | 9.95 | 788.5 | [M+H-172]+ | 617.4 | |
| Tridecanoyl-octadecenoyl -PG (C13:0/C18:1) | 9.95 | 706.5 | [M+H-172]+ | 535.4 | |
| Dioctadecanoyl-PG (C18:0/C18:0) | 9.95 | 778.5 | [M+H-172]+ | 607.4 | |
| Octadecanoyl-nonadecanoyl-PG (C18:0/C19:0) | 9.95 | 792.5 | [M+H-172]+ | 621.4 | |
| Dioleoyl-PG (C18:1/C18:1) (DOPG) | 9.95 | 774.5 | [M+H-172]+ | 603.4 | |
| Carotenoids/BChla/BPha | BChla (geranylgeranyl) | 19.06 | 904.5 | [M+H]+ | 905.5 |
| Rhodovibrin | 21.43 | 584.5 | [M+H]+ | 585.5 | |
| 1,1′-Dihydroxy-3,4-didehydrolycopene | 21.43 | 570.5 | [M+H]+ | 571.5 | |
| 1′-Hydroxyspirilloxanthin | 20.85 | 582.5 | [M+H]+ | 583.5 | |
| 1-Methoxy-1′-OH-3,4-didehydrolycopene | 20.67 | 584.5 | [M+H]+ | 585.5 | |
| 3,4-Dihydrorhodovibrin | 22.07 | 586.5 | [M+H]+ | 587.5 | |
| 1-Hydroxy-3,4-didehydrolycopene | 22.24 | 552.5 | [M+H]+ | 553.5 | |
| 3,4-Didehydrorhodopin | 22.07 | 552.5 | [M+H]+ | 553.4 | |
| Chloroxanthin (1-OH-neurosporene)) | 24.61 | 556.5 | [M+H]+ | 557.5 | |
| BPha (geranylgeranyl) | 23.60 | 882.5 | [M+H]+ | 883.5 | |
| Rhodopin | 23.63 | 554.4 | [M+H]+ | 555.4 | |
| Spirilloxanthin | 23.87 | 596.4 | [M+H]+ | 597.4 | |
| 3,4-Dihydrospirilloxanthin | 24.46 | 598.5 | [M+H]+ | 599.5 | |
| 3,4,3′,4′-Tetrahydrospirilloxanthin | 25.05 | 601.4 | [M+H]+ | 602.4 | |
| BPha (phytyl) | 26.75 | 888.5 | [M+H]+ | 889.5 | |
| 1′-Hydroxy-3,4-dihydrospheroidene | 22.72 | 588.5 | [M+H]+ | 589.5 | |
| Anhydrorhodovibrin | 25.91 | 566.5 | [M+H]+ | 567.5 | |
| 3,4-Dihydroanhydrorhodovibrin | 26.74 | 568.5 | [M+H]+ | 569.5 | |
| Phytoene | 31.25 | 544.5 | [M+H]+ | 545.5 | |
| Lycopene | 27.75 | 536.5 | [M+H]+ | 537.5 | |
| Neurosporene | 29.11 | 538.5 | [M+H]+ | 539.5 | |
| Rhodoquinone-9 (RQ-9) | 30.77 | 779.6 | [M+H]+ | 780.6 | |
| Quinones | Ubiquinol-10 (UQH2-10) | 31.08 | 864.7 | [M+H]+ | 865.7 |
| Ubiquinone-9 (UQ-9) | 31.43 | 794.6 | [M+H]+ | 795.6 | |
| Rhodoquinone-10 (RQ-10) | 32.24 | 847.7 | [M+H]+ | 848.7 | |
| Ubiquinone-10 (UQ-10) | 32.81 | 862.7 | [M+H]+ | 863.7 |
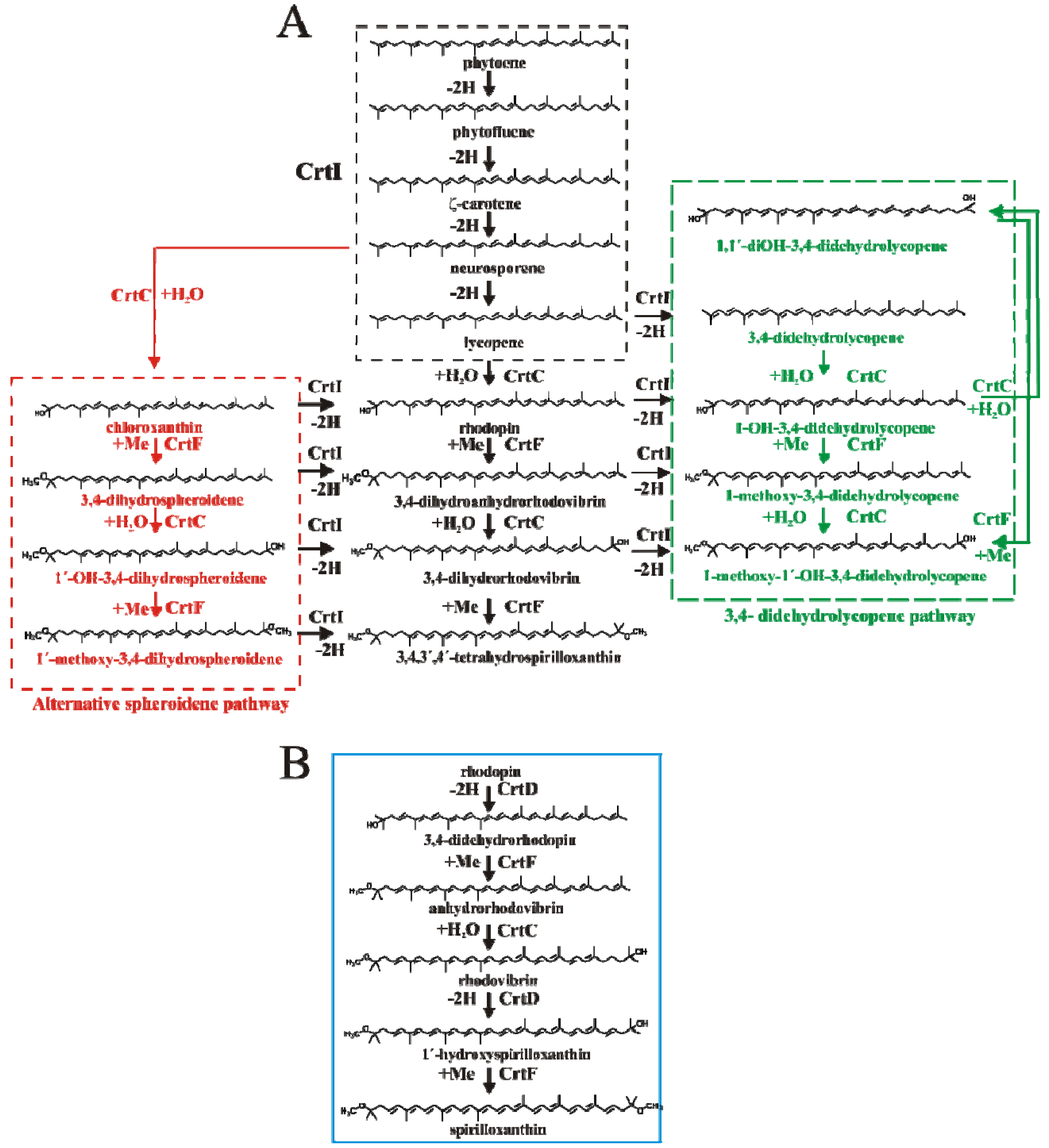
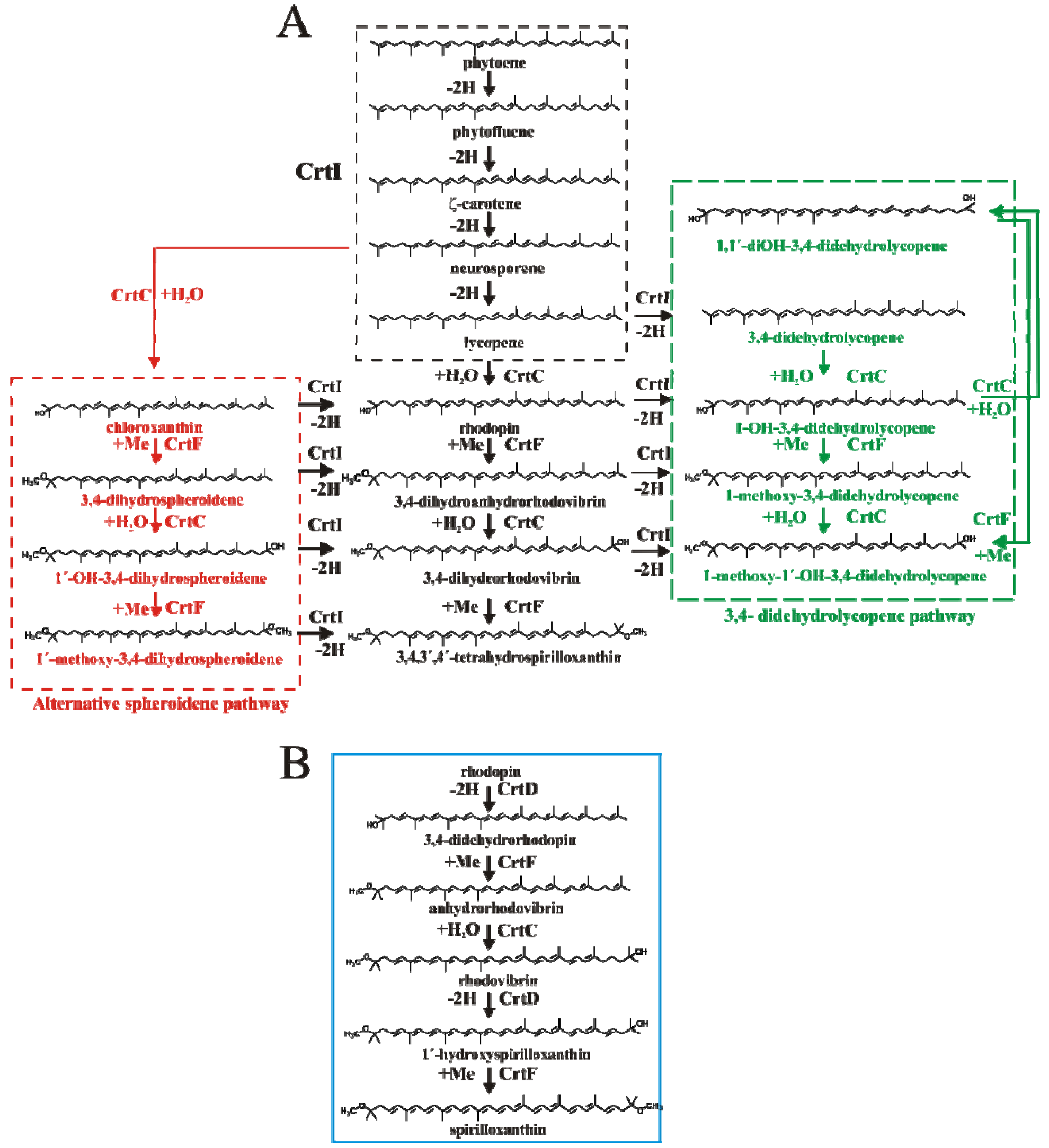
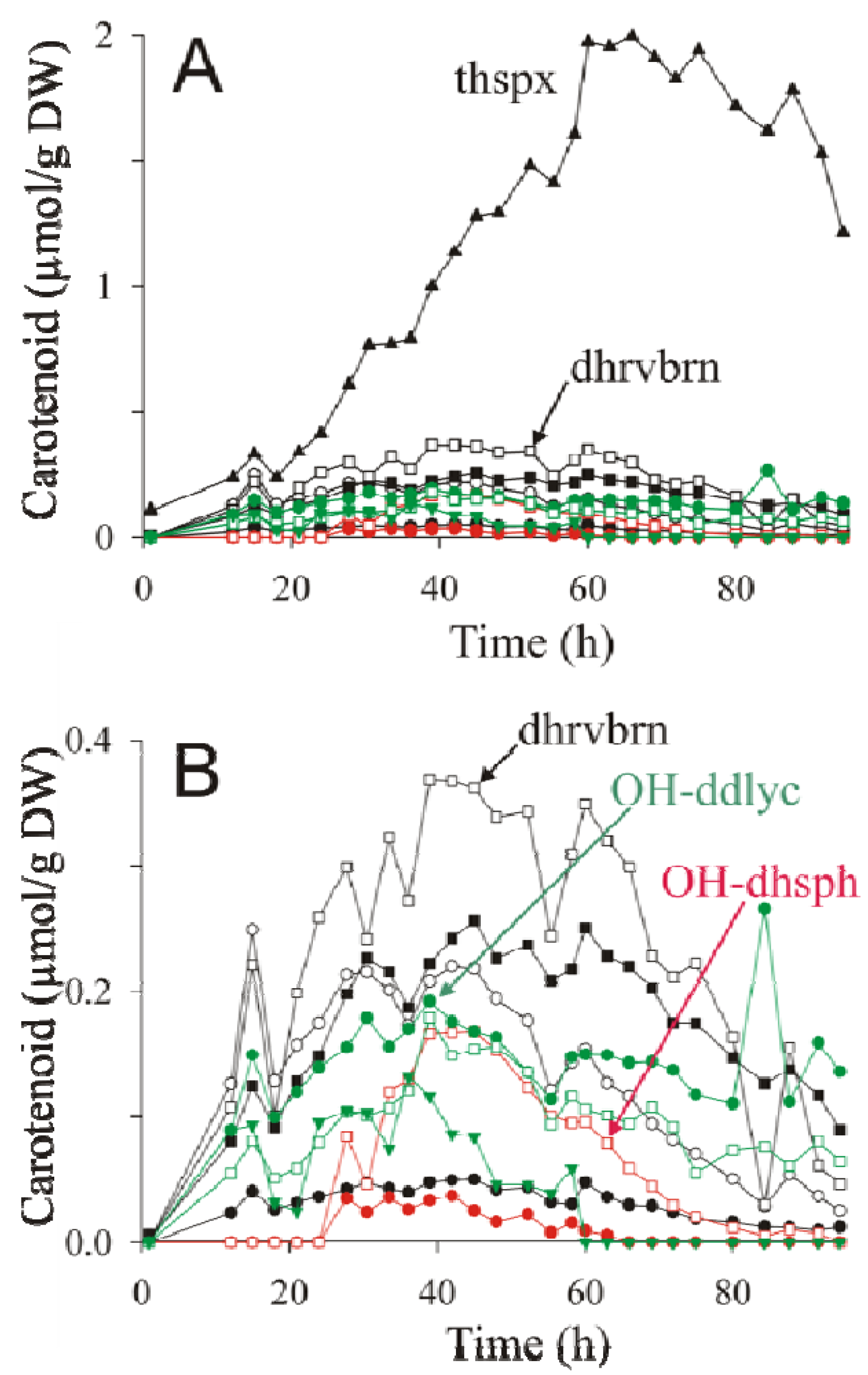
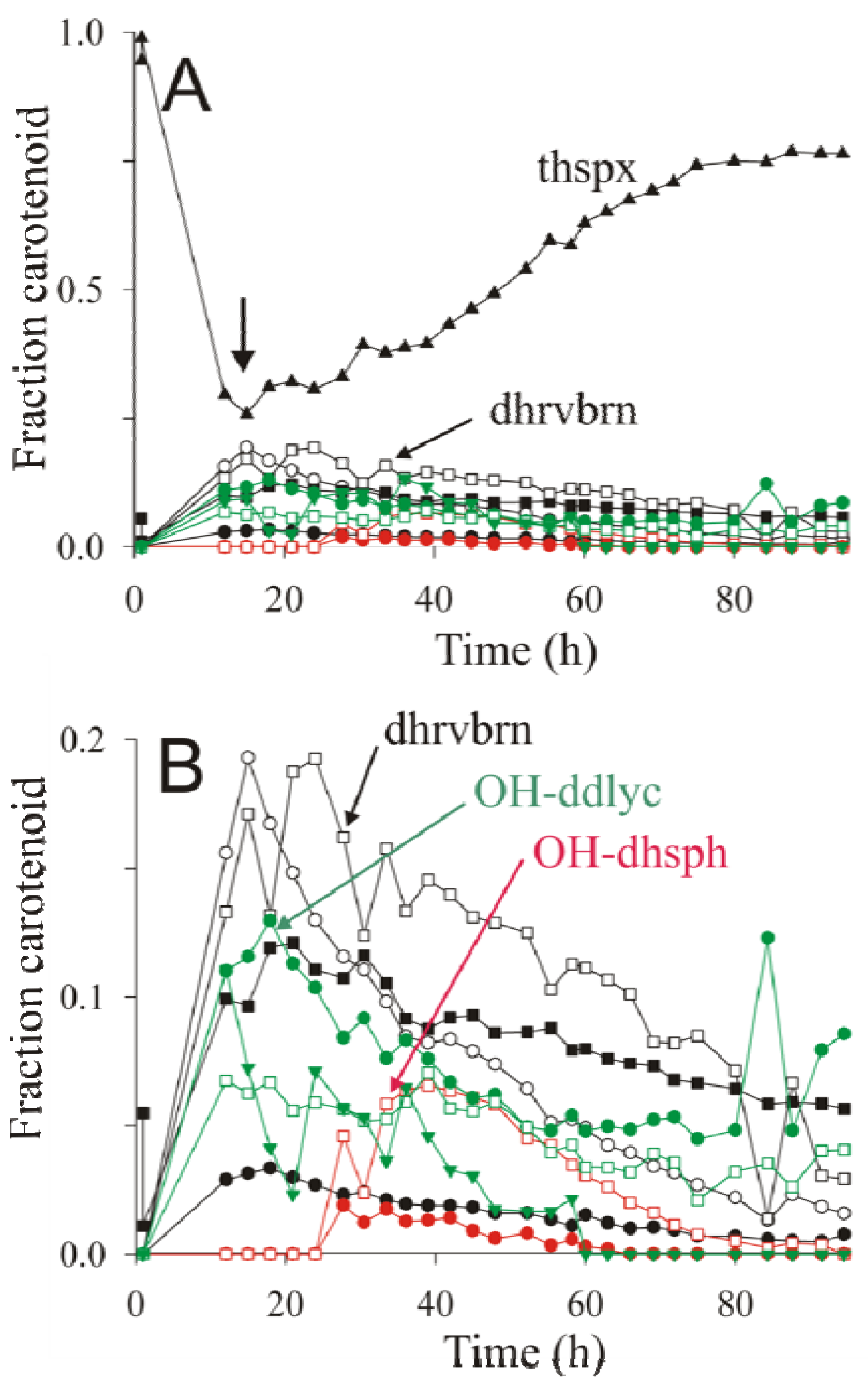
2.4. HPLC-MS Analysis of Carotenoid Pathway Dynamics
 ); 1′-OH-3,4-dihydrospheroidene (OH-dhsph) (
); 1′-OH-3,4-dihydrospheroidene (OH-dhsph) (  ); 1 -OH-3,4-didehydrolycopene (OH-ddlyc) (
); 1 -OH-3,4-didehydrolycopene (OH-ddlyc) (  ); 1-methoxy-1′-OH-didehydrolycopene (
); 1-methoxy-1′-OH-didehydrolycopene (  ); 1,1′-diOH-3,4- didehydrolycopene (
); 1,1′-diOH-3,4- didehydrolycopene (  ).
); 1′-OH-3,4-dihydrospheroidene (OH-dhsph) ( ); 1 -OH-3,4-didehydrolycopene (OH-ddlyc) ( ); 1-methoxy-1′-OH-didehydrolycopene ( ); 1,1′-diOH-3,4- didehydrolycopene ( ).
).
); 1′-OH-3,4-dihydrospheroidene (OH-dhsph) ( ); 1 -OH-3,4-didehydrolycopene (OH-ddlyc) ( ); 1-methoxy-1′-OH-didehydrolycopene ( ); 1,1′-diOH-3,4- didehydrolycopene ( ).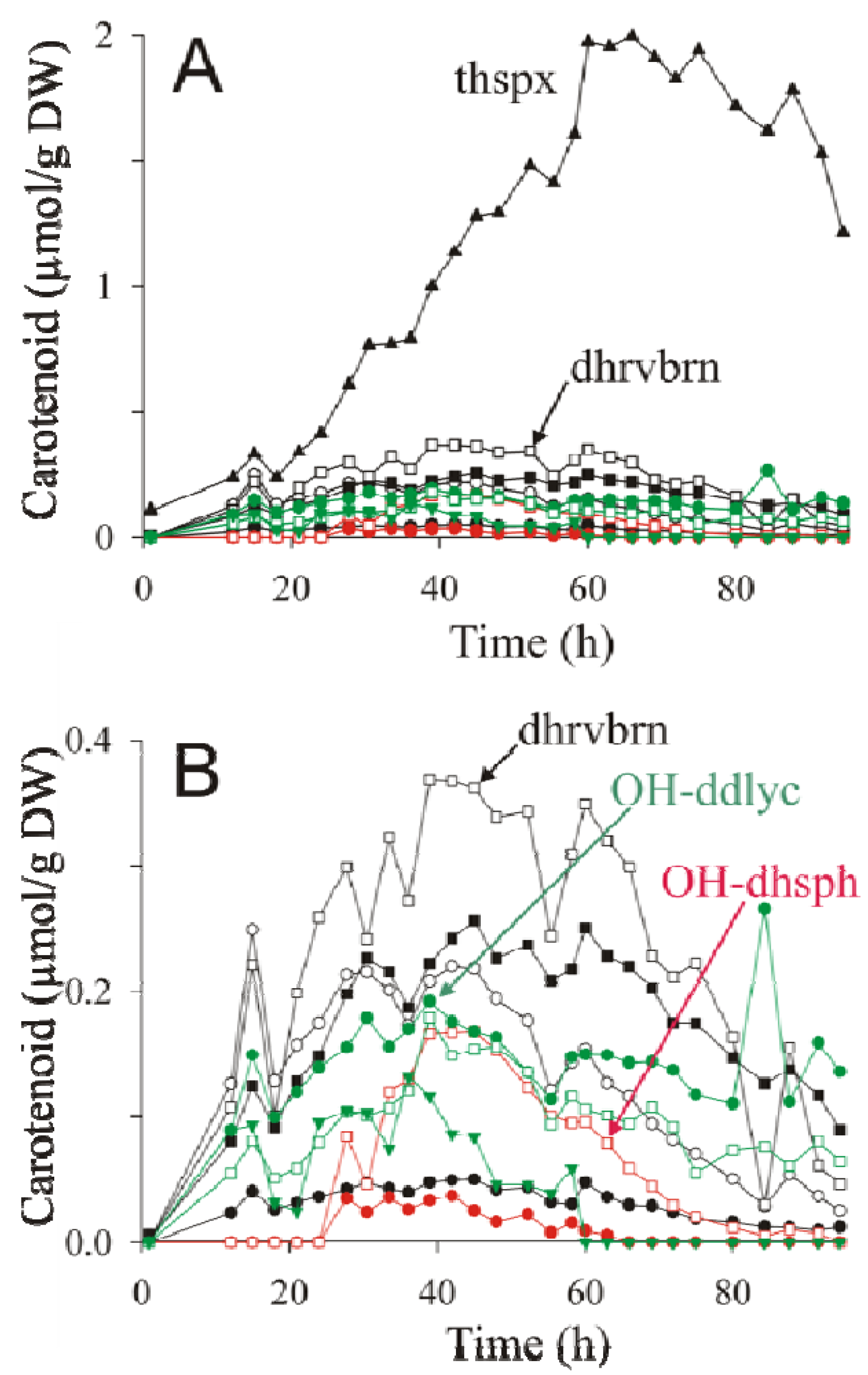
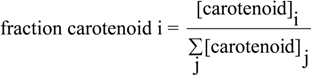
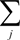 is the total concentration of all carotenoid species found at time point t.
is the total concentration of all carotenoid species found at time point t.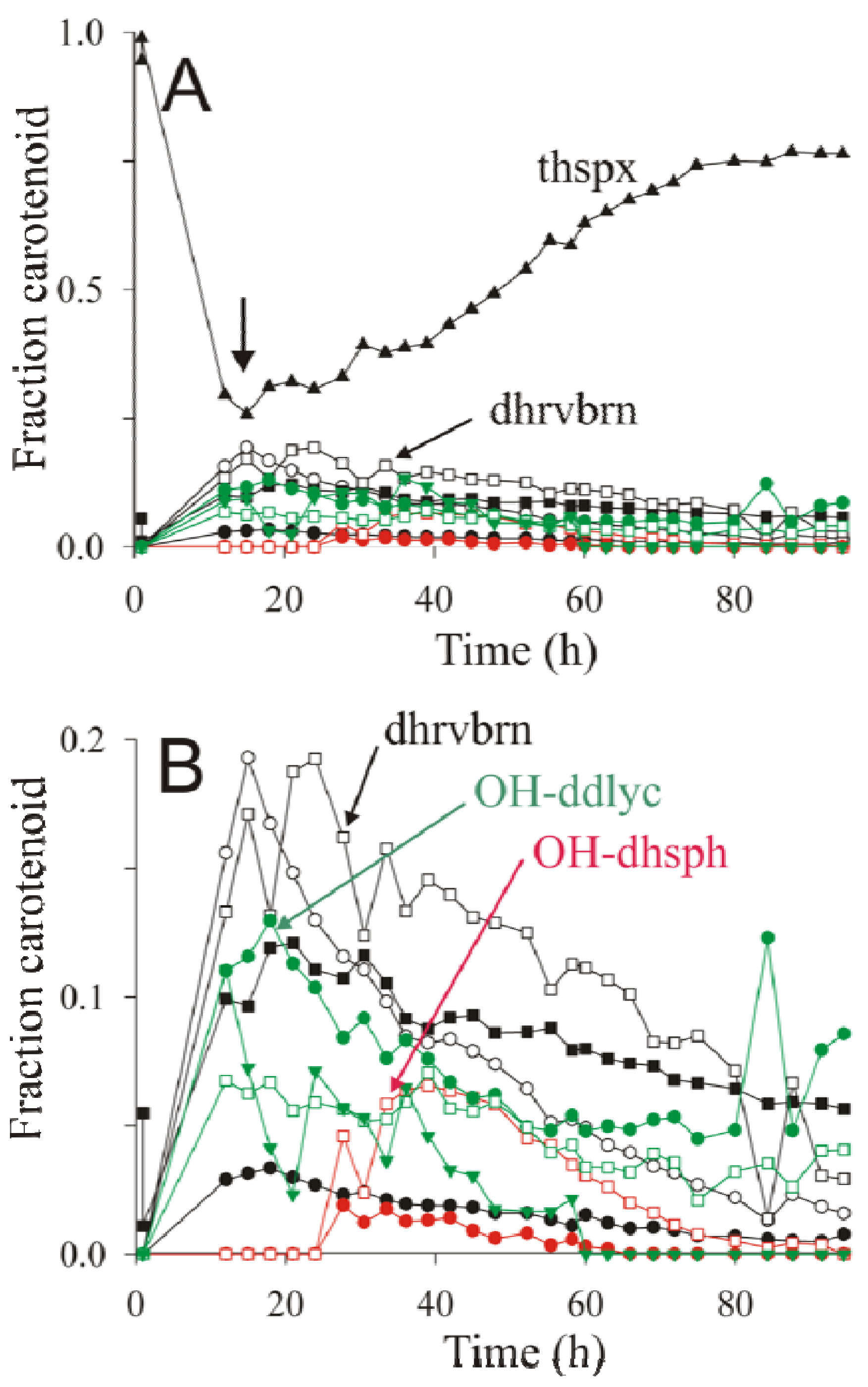
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Cell Growth of R. rubrum Strains
3.3. Extraction Method
3.4. Calibration Procedures
3.5. Instrumentation and Operating Conditions
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Kaiser, P.; Surmann, P.; Vallentin, G.; Fuhrmann, H. A small-scale method for quantitation of carotenoids in bacteria and yeasts. J. Microbiol. Meth. 2007, 70, 142–149. [Google Scholar] [CrossRef]
- Kaiser, P.; Geyer, R.; Surmann, P.; Fuhrmann, H. LC-MS method for screening unknown microbial carotenoids and isoprenoid quinones. J. Microbiol. Meth. 2012, 88, 28–34. [Google Scholar] [CrossRef]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Kolarovic, L.; Fournier, N.C.A. Comparison of extraction methods for the isolation of phospholipids from biological sources. Anal. Biochem. 1986, 156, 244–250. [Google Scholar] [CrossRef]
- Guckert, J.B.; White, D.C. Evaluation of a hexane/isopropanol lipid solvent system for analysis of bacterial analysis of bacterial phospholipids and application to chloroform-soluble Nucleopore (polycarbonate) membranes with retained bacteria. J. Microbiol. Meth. 1988, 8, 131–137. [Google Scholar] [CrossRef]
- Takada, M.; Ikenoya, S.; Yuzuriha, T.; Katayama, K. Studies on reduced and oxidized coenzyme Q (ubiquinones). II. The determination of oxidation-reduction levels of coenzyme Q in mitochondria, microsomes and plasma by high-performance liquid chromatography. Biochim. Biophys. Acta 1992, 679, 308–314. [Google Scholar]
- Cabrini, L.; Landi, L.; Stefanelli, C.; Barzanti, V.; Sechi, A.M. Extraction of lipids and lipophilic antioxidants from fish tissues: A comparison among different methods. Comp. Biochem. Phys. B 1992, 101, 383–386. [Google Scholar]
- Wang, G.-S.; Grammel, H.; Ghosh, R. High-level production of the industrial product lycopene by the photosynthetic bacterium Rhodospirillum rubrum. Appl. Environ. Microb. 2012, 78, 7205–7215. [Google Scholar] [CrossRef]
- Komori, M.; Ghosh, R.; Takaichi, S.; Hu, Y.; Mizoguchi, T.; Koyama, Y.; Kuki, M. A null lesion in the rhodopin 3,4-desaturase of Rhodospirillum rubrum unmasks a cryptic branch of the carotenoid biosynthetic pathway. Biochemistry 1998, 37, 8987–8994. [Google Scholar]
- Mercadante, A.Z.; Rodriguez-Amaya, D.B.; Britton, G. HPLC and mass spectrometric analysis of carotenoids from mango. J. Agr. Food Chem. 1997, 45, 120–123. [Google Scholar] [CrossRef]
- Careri, M.; Elviri, L.; Mangia, A. Liquid chromatography—electrospray mass spectrometry of β-carotene and xanthophylls: validation of the analytical method. J. Chromatogr. A 1999, 854, 233–244. [Google Scholar] [CrossRef]
- Breithaupt, D.E. Simultaneous HPLC determination of carotenoids used as food coloring additives: Applicability of accelerated solvent extraction. Food Chem. 2004, 86, 449–456. [Google Scholar] [CrossRef]
- Stöggl, W.; Huck, C.; Wongyai, S.; Scherz, H.; Bonn, G. Simultaneous determination of carotenoids, tocopherols, and γ-oryzanol in crude rice bran oil by liquid chromatography coupled to diode array and mass spectrometric detection employing silica C30 stationary phases. J. Sep. Sci. 2005, 28, 1712–1718. [Google Scholar] [CrossRef]
- Weber, R.W.S.; Anke, H.; Davoli, P. Simple method for the extraction and reversed-phase high-performance liquid chromatographic analysis of carotenoid pigments from red yeasts (Basidiomycota, Fungi). J. Chromatogr. A 2007, 1145, 118–122. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Lu, H.; Hung, C.F.; Wu, W.B.; Lin, C.L.; Chen, B.H. Determination of carotenoids and their esters in fruits of Lycium barbarum Linnaeus by HPLC-DAD-APCI-MS. J. Pharm. Biomed. Anal. 2008, 47, 812–818. [Google Scholar] [CrossRef]
- Kurz, C.; Carle, R.; Schieber, A. HPLC-DAD-MS characterisation of carotenoids from apricots and pumpkins for the evaluation of fruit product authenticity. Food Chem. 2008, 110, 522–530. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Isaji, M.; Harada, J.; Tamiaki, H. Identification of 3,4-didehydrorhodopin as major carotenoid in Rhodopseudomonas species. Photochem. Photobiol. Sci. 2008, 7, 492–497. [Google Scholar] [CrossRef]
- Karlsson, A.Å.; Michelsen, P.; Larsen, Å.; Odham, G. Normal-phase liquid chromatography class separation and species determination of phospholipids utilizing electrospray mass spectrometry/tandem mass spectrometry. Rapid Commun. Mass Sp. 1996, 10, 775–780. [Google Scholar] [CrossRef]
- Tolonen, A.; Lehto, T.M.; Hannuksela, M.L.; Savolainen, M.J. A method for determination of phosphatidylethanol from high density lipoproteins by reversed-phase HPLC with TOF-MS detection. Anal. Biochem. 2005, 341, 83–88. [Google Scholar] [CrossRef]
- Sommer, U.; Herscovitz, H.; Welty, F.K.; Costello, C.E. LC-MS-based method for the qualitative and quantitative analysis of complex lipid mixtures. J. Lipid Res. 2006, 47, 804–814. [Google Scholar] [CrossRef]
- Lang, J.; Gohil, K.; Packer, L. Simultaneous determination of tocopherols, ubiquinols, and ubiquinones in blood, plasma, tissue homogenates, and subcellular fractions. Anal. Biochem. 1986, 157, 106–116. [Google Scholar] [CrossRef]
- Mitchell, K.; Fallon, R.J. The determination of ubiquinone profiles by reversed-phase high-performance thin-layer chromatography as an aid to the speciation of Legionellaceae. J. Gen. Microbiol. 1990, 136, 2035–2041. [Google Scholar] [CrossRef]
- Lytle, C.; Gan, Y.D.; Salone, K.; White, D.C. Sensitive characterization of microbial ubiquinones from biofilms by electrospray/mass spectrometry. Environ. Microbiol. 2001, 3, 265–272. [Google Scholar] [CrossRef]
- Mahendra, K.; Murthy, Y.L.N.; Narasimha Rao, C.V.; Bala Murali Krishna, K. Determination of ubiquinone Q10 (coenzyme Q10) and its synthesis related impurities by high performance liquid chromatography and mass spectrometry. Int. J. PharmTech Res. 2011, 3, 1467–1477. [Google Scholar]
- Ruiz-Jiménez, J.; Priego-Capote, F.; Mata-Granados, J.M.; Quesada, J.M.; Luque de Castro, M.D. Determination of the ubiquinol-10 and ubiquinone-10 (coenzyme Q10) in human serum by liquid chromatography tandem mass spectrometry to evaluate the oxidative stress. J. Chromatogr. A 2007, 1175, 242–248. [Google Scholar] [CrossRef]
- Airs, R.L.; Atkinson, J.E.; Keely, B.J. Development and application of a high resolution liquid chromatographic method for the analysis of complex pigment distributions. J. Chromatogr. A 2001, 917, 167–177. [Google Scholar] [CrossRef]
- Airs, R.L.; Keely, B.J. A high resolution study of the chlorophyll and bacteriochlorophyll pigment distributions in a calcite/gypsum microbial mat. Org. Geochem. 2003, 34, 539–551. [Google Scholar] [CrossRef]
- Katz, J.J.; Strain, H.H.; Harkness, A.L.; Studier, M.H.; Svec, W.A.; Janson, T.R.; Cope, B.T. Esterifying alcohols in the chlorophylls of purple photosynthetic bacteria. New chlorophyll, bacteriochlorophyll (gg), all-trans-geranylgeranyl bacteriochlorophyllide a. J. Am. Chem. Soc. 1972, 94, 7938–7939. [Google Scholar]
- Walter, E.; Schreiber, J.; Zass, E.; Eschenmoser, A. Bakteriochlorophyll aGg und Bakteriophäophytin aP in den photosynthetischen Reaktionszentren von Rhodospirillum rubrum G-9+. (in German). Helv. Chim. Acta 1979, 62, 899–920. [Google Scholar] [CrossRef]
- Hirayama, O. Lipids and lipoprotein complex in photosynthetic tissues. Agric. Biol. Chem. 1968, 32, 34–41. [Google Scholar] [CrossRef]
- Snozzi, M.; Bachofen, R. Characterisation of reaction centers and their phospholipids from Rhodospirillum rubrum. Biochim. Biophys. Acta 1979, 546, 236–247. [Google Scholar] [CrossRef]
- Wood, B.; Nichols, B.; James, A. The lipids and fatty acid metabolism of photosynthetic bacteria. Biochim. Biophys. Acta 1965, 106, 261–273. [Google Scholar] [CrossRef]
- Schröder, J.; Drews, G. Quantitative Bestimmung der Fettsäuren von Rhodospirillum rubrum und Rhodopseudomonas capsulata während der Thylakoidmorphogenese. Arch. Mikrobiol. 1968, 64, 59–70. (in German). [Google Scholar] [CrossRef]
- Oelze, J.; Golecki, J.R.; Kleinig, H.; Weckesser, J. Characterization of two cell-envelope fractions from chemotrophically grown Rhodospirillum rubrum. Anton. Leeuw. 1975, 41, 273–286. [Google Scholar] [CrossRef]
- Collins, M.L.; Niederman, R.A. Membranes of Rhodospirillum rubrum: Isolation and physicochemical properties of membranes from aerobically grown cells. J. Bacteriol. 1976, 126, 1316–1325. [Google Scholar]
- Russell, N.J.; Harwood, J.L. Changes in the acyl lipid composition of photosynthetic bacteria grown under photosynthetic and non-photosynthetic conditions. Biochem. J. 1979, 181, 339–345. [Google Scholar]
- Jensen, S.L.; Cohen-Bazire, G.; Nakayama, T.O.M.; Stanier, R.Y. The path of carotenoid synthesis in a photosynthetic bacterium. Biochim. Biophys. Acta 1958, 29, 477–499. [Google Scholar] [CrossRef]
- Davies, B.H. A novel sequence for phytoene dehydrogenation in Rhodospirillum rubrum. Biochem. J. 1970, 116, 93–99. [Google Scholar]
- Schwerzmann, R.U.; Bachofen, R. Carotenoid profiles in pigment-protein complexes of Rhodospirillum rubrum. Plant Cell Physiol. 1989, 30, 497–504. [Google Scholar]
- Schmidt-Dannert, C.; Umeno, D.; Arnold, F.H. Molecular breeding of carotenoid biosynthetic pathways. Nat. Biotechnol. 2000, 18, 750–753. [Google Scholar] [CrossRef]
- Albrecht, M.; Takaichi, S.; Steiger, S.; Wan, Z.-Y.; Sandmann, G. Novel hydroxycarotenoids with improved antioxidative properties produced by gene combination in Escherichia coli. Nat. Biotechnol. 2000, 18, 843–846. [Google Scholar] [CrossRef]
- Yen, H.-C.; Marrs, B. Map of genes for carotenoid and bacteriochlorophyll biosynthesis in Rhodopseudomonas capsulata. J. Bacteriol. 1976, 126, 619–629. [Google Scholar]
- Grammel, H.; Gilles, E.-D.; Ghosh, R. Microaerophilic cooperation of oxidative and reductive pathways allows maximal photosynthetic membrane biosynthesis in Rhodospirillum rubrum. Appl. Environ. Microb. 2003, 69, 6577–6586. [Google Scholar] [CrossRef]
- Sistrom, W.R. A requirement for sodium in the growth of Rhodopseudomonas sphaeroides. J. Gen. Microbiol. 1960, 22, 778–785. [Google Scholar] [CrossRef]
- Ghosh, R.; Hardmeyer, A.; Thoenen, I.; Bachofen, R. Optimization of the Sistrom culture medium for large-scale batch cultivation of Rhodospirillum rubrum under semiaerobic conditions with maximal yield of photosynthetic membranes. Appl. Environ. Microb. 1994, 60, 1698–1700. [Google Scholar]
- Young, A.; Britton, G. Carotenoids in Photosynthesis; Chapman & Hall: London, UK, 1993; pp. 459–488. [Google Scholar]
- Callender, H.L.; Forrester, J.S.; Ivanova, P.; Preininger, A.; Milne, S.; Brown, H.A. Quantification of diacylglycerol species from cellular extracts by electrospray ionization mass spectrometry using a linear regression algorithm. Anal. Chem. 2007, 79, 263–272. [Google Scholar] [CrossRef]
- Olson, T.L.; van de Meene, A.M.L.; Francis, J.N.; Pierson, B.K.; Blankenship, R.E. Pigment analysis of “Candidatus Chlorothrix halophila”, a green filamentous anoxygenic phototrophic bacterium. J. Bacteriol. 2007, 189, 4187–4195. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bóna-Lovász, J.; Bóna, A.; Ederer, M.; Sawodny, O.; Ghosh, R. A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria. Metabolites 2013, 3, 912-930. https://doi.org/10.3390/metabo3040912
Bóna-Lovász J, Bóna A, Ederer M, Sawodny O, Ghosh R. A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria. Metabolites. 2013; 3(4):912-930. https://doi.org/10.3390/metabo3040912
Chicago/Turabian StyleBóna-Lovász, Judit, Aron Bóna, Michael Ederer, Oliver Sawodny, and Robin Ghosh. 2013. "A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria" Metabolites 3, no. 4: 912-930. https://doi.org/10.3390/metabo3040912