

The Exploration of Fetal Growth Restriction Based on Metabolomics: A Systematic Review
by
Mengxin Yao
1,
Zhuoqiao Yang
1,
Xin Rong
1,
Xuan Hu
1,
Na Yao
1,
Manting Zhu
1,
Xinnan Wang
1,
Xiaoyan Zhu
2,3,4,* and
Jieyun Yin
1,* 1
Department of Epidemiology and Health Statistics, Medical College of Soochow University, Suzhou 215000, China
2
Suzhou Center for Disease Prevention and Control, Suzhou 215000, China
3
Institute of Suzhou Biobank, Suzhou 215000, China
4
School of Public Health, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China
*
Authors to whom correspondence should be addressed.
Metabolites 2022, 12(9), 860; https://doi.org/10.3390/metabo12090860
Submission received: 3 August 2022
/
Revised: 3 September 2022
/
Accepted: 4 September 2022
/
Published: 13 September 2022
(This article belongs to the Special Issue Metabolic Profiles and Biomarkers in Pregnancy)
Abstract
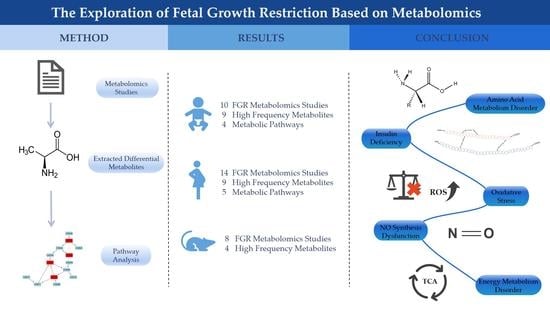
:Fetal growth restriction (FGR) is a common complication of pregnancy and a significant cause of neonatal morbidity and mortality. The adverse effects of FGR can last throughout the entire lifespan and increase the risks of various diseases in adulthood. However, the etiology and pathogenesis of FGR remain unclear. This study comprehensively reviewed metabolomics studies related with FGR in pregnancy to identify potential metabolic biomarkers and pathways. Relevant articles were searched through two online databases (PubMed and Web of Science) from January 2000 to July 2022. The reported metabolites were systematically compared. Pathway analysis was conducted through the online MetaboAnalyst 5.0 software. For humans, a total of 10 neonatal and 14 maternal studies were included in this review. Several amino acids, such as alanine, valine, and isoleucine, were high frequency metabolites in both neonatal and maternal studies. Meanwhile, several pathways were suggested to be involved in the development of FGR, such as arginine biosynthesis, arginine, and proline metabolism, glyoxylate and dicarboxylate metabolism, and alanine, aspartate, and glutamate metabolism. In addition, we also included 8 animal model studies, in which three frequently reported metabolites (glutamine, phenylalanine, and proline) were also present in human studies. In general, this study summarized several metabolites and metabolic pathways which may help us to better understand the underlying metabolic mechanisms of FGR.
1. Introduction
Fetal growth restriction (FGR) is an obstetric complication defined as the failure of a fetus to attain its pre-determined intrauterine growth potential, which is also referred to intrauterine growth restriction (IUGR) [1]. Usually, these two terms (FGR and IUGR) are considered to be synonymous and interchangeable [2]. When a newborn’s birth weight falls below a predefined threshold for its gestational age, it is considered as small for gestational age (SGA). Compared with FGR, SGA is an auxological but not an etiological definition. It is often assumed that FGR results in SGA [3].Therefore, we do not distinguish the above-mentioned terms as we aimed to discuss their etiology, and FGR is used below in this article.
Usually, FGR is classified into early-onset FGR (<32 weeks) and late-onset FGR (≥32 weeks) based on gestational age of diagnosis [4]. With an incidence of approximately 10%, FGR is second only to preterm birth as a cause of infant morbidity and mortality [5]. Not limited to the neonatal period, the adverse effects of poor fetal growth may also last throughout the entire lifespan [6]. For example, several studies have shown that FGR increases the risks of cognitive delay, obesity, cardiovascular disease, and type 2 diabetes in later life [7,8,9,10]. Currently, the main clinical diagnostic methods of FGR are symphysis-fundal height and ultrasound. The former is widely used due to its low cost and convenience [11]. However, maternal obesity, uterine leiomyomas, and polyhydramnios may limit its effectiveness in detecting FGR. Regarding ultrasound, on one hand, its relatively high cost may impede its usage in low resource areas. On the other hand, body habitus, operator experience, and fetal position may influence its accuracy [4]. Meanwhile, the phenotypes of late-onset FGR differ significantly from that of early-onset FGR. Thus, a single fetal biometric measurement is insufficient to evaluate fetal growth, except perhaps in the case of extremely small fetal size [12]. Perhaps, complementary biological tests might help us to identify FGR. In addition, despite FGR is known to be a multifactorial disease that affected by maternal, placental, fetal, and genetic factors [13,14], its etiology and pathogenesis remain unclear. Therefore, enhanced research on the etiology, pathogenesis, and metabolic drivers of FGR is urgently required for accurately monitoring fetal growth.
Metabolomics is an emerging high-throughput technique that enables the comprehensive and systematic identification and quantification of small molecules in biological systems. Previous studies have shown that metabolomics improves our understanding of the pathology of many diseases and is a promising tool for disease diagnosis [15,16]. Therefore, metabolomics may provide us with new insights into FGR.
In this study, we reviewed all metabolomics studies related with FGR in pregnancy over the last 20 years. We also comprehensively summarized and analyzed the information from these studies. The review aimed to (1) better understand the specific metabolic mechanisms of FGR, and (2) identify metabolic biomarkers that can improve diagnostic and predictive capabilities of FGR.
2. Materials and Methods
2.1. Literature Search
This systematic review was conducted in compliance with PRISMA guidelines [17], and a review protocol was entered into the Prospero database (registration number 356869). We obtained relevant publications from the PubMed and Web of Science databases from January 2000 to July 2022. The search terms were (“metabolome” or “metabolomics” or “metabolite” or “metabonomics” or “metabolic profiling” or “metabolic signature” or “metabolic biomarker” or “metabolic profile” or “metabolic portraits”) AND (“fetal growth restriction” or “intrauterine growth restriction” or “small for gestational age”). Two researchers searched the articles independently, and a third researcher made a final decision in cases of disagreement.
2.2. Inclusion and Exclusion Criteria
The inclusion criteria were (1) metabolomics studies for pregnancies, (2) full text in English, and (3) studies recorded the positive or negative relationship between metabolite markers and FGR.
The exclusion criteria were as follows: (1) review articles, (2) non-perinatal studies or FGR fetus disturbed by diet, (3) studies evaluating drug effects, (4) abstracts without full articles, and (5) studies only reported insignificant metabolites.
2.3. Data Extraction
A total of 29 studies, consisting of 21 human studies and 8 animal studies, were eligible for this systematic review (Figure 1). We extracted the following information after reading the full articles and supplementary materials: (1) basic information of included studies, including first author, published year and journal; (2) basic information of subjects, including species, sample sizes, and singleton/twins; (3) study design, diagnostic criteria, or model of FGR, biological specimen, sampling time, and analytic platform; (4) the significant metabolites with changing trends. In addition, studies by the same first or corresponding author were checked as to whether there were overlaps in the content.
2.4. Statistical Analysis
The frequencies on biological specimens, targeted/untargeted, analytic platforms, sample sizes, and repeatedly reported biomarkers were computed and charted. Pathway analysis, including enrichment analysis and topology analysis, was performed using the online MetaboAnalyst software (version 5.0, Mcgill University, Montreal, QC, Canada; https://www.metaboanalyst.ca, accessed on 10 July 2022) [18]. The significant pathways were selected based on the criteria of the false discovery rate (FDR) < 0.05.
3. Results
3.1. Human Neonatal Research
3.1.1. Study Characteristics of Human Neonatal Research
As shown in Table 1, a total of ten studies [19,20,21,22,23,24,25,26,27,28] on human neonatal samples were included. Eight studies used umbilical cord blood, and the remaining two studies collected neonatal blood and urine (Figure 2a). Seven of the nine studies were untargeted, two were targeted, and the remaining one used untargeted and targeted method simultaneously (Figure 2b). Besides, six metabolomics studies used nuclear magnetic resonance (NMR), three used mass spectrometry (MS), and one study used both (Figure 2c). For the sample sizes, the majority of the studies ranged from 50 to 100 (Figure 2d).
3.1.2. Analysis of High Frequency Biomarkers in Human Neonatal Research
Among the ten studies included, the frequencies of significant metabolites in different studies were counted (Table 2). We found several high frequency biomarkers (reported in ≥3 studies). Ranked by reported frequency, the top nine metabolites were alanine, proline, valine, phenylalanine, glutamine, isoleucine, creatine, tryptophan, and choline. Most high frequency metabolites belong to amino acids. Choline showed a consistent changing trend in FGR, and Sanz-Cortés et al. [24] reported that this biomarker was down-regulated in both early-onset and late-onset FGR.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
High Frequency Metabolic Biomarkers of Human Neonatal Studies and Maternal Studies.
| Neonatal Studies | ||||||
| No | Metabolites | Total Hits | Up | Down | ||
| Hits | Bio-Specimen | Hits | Bio-Specimen | |||
| 1 | Alanine | 5 | 3 | Cord serum [19] Dried blood [26] Cord plasma [28] | 2 | Cord serum [24] (l) Cord plasma [25] |
| 2 | Proline | 4 | 3 | Cord serum [21] Dried blood [26] Cord plasma [28] | 1 | Cord plasma [25] |
| 3 | Valine | 4 | 2 | Cord serum [19,21] | 2 | Cord serum [24] (l) Dried blood [26] |
| 4 | Phenylalanine | 4 | 2 | Cord serum [21] Cord plasma [25] | 2 | Cord serum [19,24] (e) |
| 5 | Glutamine | 4 | 2 | Cord serum [24] (e) Cord plasma [28] | 2 | Cord serum [24] (l) Cord plasma [25] |
| 6 | Isoleucine | 3 | 2 | Cord serum [19,21] | 1 | Cord plasma [20] |
| 7 | Creatine | 3 | 2 | Cord serum [24] (e) Urine [27] | 1 | Cord serum [22] |
| 8 | Tryptophan | 3 | 1 | Cord serum [21] | 2 | Cord serum [19] Cord plasma [28] |
| 9 | Choline | 3 | - | - | 3 | Cord serum [22,24] (e l) Cord plasma [25] |
| Maternal Studies | ||||||
| No | Metabolites | Total Hits | Up | Down | ||
| Hits | Bio-Specimen | Hits | Bio-Specimen | |||
| 1 | Alanine | 5 | 1 | Maternal serum [19] | 4 | Urine [29] Maternal hair [30] Maternal plasma [23] Placenta [31] |
| 2 | Citrate | 4 | 2 | Placenta [31] Human-milk [32] | 2 | Urine [29] Maternal plasma [23] |
| 3 | Valine | 4 | 1 | Maternal serum [19] | 3 | Maternal hair [30] Placenta [31] Human-milk [32] |
| 4 | Glycine | 4 | - | - | 4 | Urine [29] Maternal hair [30] Placenta [28,31] |
| 5 | Isoleucine | 3 | 1 | Maternal serum [19] | 2 | Maternal hair [30] Human-milk [32] |
| 6 | Lactate | 3 | 1 | Placenta [33] | 2 | Urine [29] Maternal hair [30] |
| 7 | Tyrosine | 3 | - | - | 3 | Urine [29] Maternal hair [30] Placenta [28] |
| 8 | Aspartate | 3 | - | - | 3 | Maternal hair [30] Placenta [28,31] |
| 9 | 3-Hydroxybutyrate | 3 | - | - | 3 | Maternal serum [19,34] Placenta [31] |
l, late-onset FGR; e, early-onset FGR.
3.1.3. Metabolic Pathway Analysis of Potential Biomarkers in Human Neonatal Research
To understand the metabolic pathways that these significant biomarkers are involved in, we imported all the reported metabolites in neonatal research into the online MetaboAnalyst 5.0 software for pathway analysis. The results showed that six pathways were significantly enriched (FDR < 0.05, shown in Table S1). Particularly, four pathways ((1) glyoxylate and dicarboxylate metabolism; (2) arginine biosynthesis; (3) arginine and proline metabolism; (4) alanine, aspartate, and glutamate metabolism) possessed relatively high impact values (Figure 3a).
3.2. Human Maternal Research
3.2.1. Study Characteristics of Human Maternal Research
A total of fourteen articles [19,23,28,29,30,31,32,33,34,35,36,37,38,39] of maternal studies were included in the final analysis (Table 3). The characteristics of human maternal and neonatal research were similar. Among these included studies, maternal blood was the most common bio-specimen. Urine, hair, human-milk, and placenta were also used (Figure 2a). Similar to neonatal studies, researchers usually conducted untargeted studies (Figure 2b). For the analytical platform, six studies were NMR, seven were MS, and the remaining one used both (Figure 2c). The number of cases among the maternal studies varied from 10 to 175, while the sample sizes of most studies were between 50 and 100 (Figure 2d).
3.2.2. Analysis of High Frequency Biomarkers in Human Maternal Research
A total of nine high frequency metabolites (reported in ≥ 3 studies) were revealed in maternal derived studies, including alanine, citrate, valine, glycine, isoleucine, lactate, tyrosine, aspartate, and 3-hydroxybutyrate (Table 2). These metabolites belong to amino acids and organic acids. Among them, glycine, tyrosine, aspartate and 3-hydroxybutyrate showed a consistent down-regulation trend in all reported human maternal articles.
3.2.3. Metabolic Pathway Analysis of Potential Biomarkers in Human Maternal Research
After inputting significant metabolites of maternal research into MetaboAnalyst, a total of seven pathways were identified (FDR < 0.05, Table S2). Five pathways had relatively high impact values, they were (1) glutathione metabolism; (2) arginine biosynthesis; (3) arginine and proline metabolism; (4) glyoxylate and dicarboxylate metabolism; (5) alanine, aspartate, and glutamate metabolism (Figure 3b).
3.3. Comparison between Human Neonatal and Maternal Research
A number of high frequency metabolites, especially amino acids (alanine, valine, and isoleucine), were simultaneously revealed from neonatal and maternal research. In addition, four pathways with FDR < 0.5 were significantly enriched in both neonatal and maternal studies. They were glyoxylate and dicarboxylate metabolism; arginine biosynthesis; arginine and proline metabolism; and alanine, aspartate, and glutamate metabolism.
3.4. Classification Potential of the Metabolic Biomarkers
Seven studies [19,22,30,31,34,36,38] have assessed the potential of metabolic biomarkers or biomarkers panels to predict or diagnose FGR (Table 4). Moros et al. [19] calculated the area under the receiver operating curve (AUC) of several metabolites one at a time, resulting in all of the AUC values greater than 0.750. Sovio et al. [36] explored the discriminating potential of various combinations of the four metabolites (1-(1-enyl-stearoyl)-2-oleoyl-GPC (P-18:0/18:1); 1,5-anhydroglucitol; 5α-androstan-3α,17α-diol disulfate; N1,N12-diacetylspermine), yielding AUC values ranging from 0.640 to 0.780. Bahado-Singh and coworkers [22,31] reported that the incorporation of metabolites with maternal factors did not improve predictive accuracy over the best biomarkers panels. Sulek et al. [30] uncovered a panel of five metabolite parameters (lactate, levulinate, 2-methyloctadecanat, tyrosine, and margarate) with an AUC value of 0.998, suggesting that this model’s ability to distinguish FGR was remarkable.
3.5. Animal Studies
3.5.1. Study Characteristics of Animal Model Research
According to the inclusion and exclusion criteria, a total of 8 studies [40,41,42,43,44,45,46,47] on animal models were included. The characteristics of these studies are presented in Table 5. The animals involved were rat, sheep, calf, and piglet. These studies were all conducted in newborn animals. Compared with human studies, the types of biological samples in animal studies were diversity, including fetal plasma, umbilical cord plasma, skeletal muscle, muscle, liver, and kidney. Untargeted method was commonly used to detect metabolites, and the analysis platform was always MS in animal studies. Most animal studies had sample size of less than 20. Only one study [47] for fetal rats had 53 cases and 57 controls.
3.5.2. Analysis of High Frequency Biomarkers in Animal Research
Four high frequency (reported in ≥ 3 studies) differential metabolites extracted from these 8 animal studies are shown in Table 6. These metabolites are arginine, reported four times, and glutamine, phenylalanine, and proline, which occurred three times. These differential metabolites in FGR animal fetus were all amino acids, which was similar to the human study. Among them, glutamine, phenylalanine, and proline were also high frequency metabolites reported in human neonatal studies.
4. Discussion
In this systematic review, we comprehensively screened and analyzed metabolomics studies on FGR around delivery. In human research, neonatal and maternal studies each identified several possible metabolic pathways and four of them were consistent, suggesting similar metabolic patterns of mothers and newborns in FGR pregnancies. Three high frequency metabolites (glutamine, phenylalanine, and proline) found in the human neonatal studies were also reported in the animal model studies. In addition, several metabolic biomarkers or biomarker panels showed classification potential for FGR.
4.1. Potential Metabolic Dysregulations of FGR
4.1.1. Amino Acid Metabolism Disorder
All of the three high frequency metabolites in both human and animal neonatal studies were amino acids. Glutamine [24,25,28,43,44,47] is the most abundant amino acid in the body [48], which is not only important for energy supply, but is also a precursor for nucleotide synthesis [49]. Phenylalanine [19,21,24,25,43,44,47] belongs to aromatic amino acids, as precursor of the monoamine neurotransmitters, serotonin and catecholamines in the brain [50]. Proline [21,25,26,28,40,43,44] plays a role in maintaining cellular homeostasis and modulating mitochondrial functions [51].
In addition, several amino acids were frequently reported to be downregulated in human maternal studies. The reported valine [19,30,31,32] and isoleucine [19,30,32] are essential branched chain amino acids (BCAAs), which can activate mTOR signaling pathway to stimulate protein synthesis and cell growth [52]. Lowered concentrations of BCAAs may result in disturbed transportation of placental amino acid to fetus and therefore impaired fetal growth [53,54]. Alanine [19,23,29,30,31] was reported several times. Alanine can be synthesized from BCAAs. As a key amino acid in nitrogen metabolism, alanine provides energy for muscle tissue and the central nervous systems via gluconeogenic pathway [55]. Glycine showed consistent down-regulated trend in the included studies [28,29,30,31]. It is involved in body’s collagen, immune response, and plays a crucial role as neurotransmitter.
In general, amino acids are the precursor of many biologically active molecules, and play a key role in regulating cell metabolism, proliferation, differentiation, and growth [56]. Our results supported the view that low concentrations of maternal amino acids play an important role in the occurrence and development of fetal abnormal growth.
4.1.2. Insulin Deficiency
Insulin is an important hormone for intrauterine growth. Pathway analysis suggested that pathways of alanine, aspartate, and glutamate metabolism, which may affect insulin secretion, are related to FGR. Alanine [19,23,24,25,26,28,29,30,31,41,45] was the most frequently reported metabolite in the included studies. A previous study suggested alanine metabolism provides key stimulus-secretion coupling factors that are critical for promoting insulin secretion [57]. Glutamate [21,30,34], which was revealed to be an important mediator in the amplification of insulin secretion [58], is the major precursor of the repeatedly reported glutamine [24,25,28,33,34,43,44,47]. Fetal insulin deficiency can not only decrease fetal tissues to uptake and utilize nutrient, but also lower circulating concentrations of insulin-like growth factors (IGFs) [59]. Animal studies showed that maternal plasma IGFs correlate positively with fetal growth and birth weight [60]. In addition to suppressing fetal growth during pregnancy, the effects of insulin deficiency may persist into postpartum and even adulthood. To compensate for intrauterine growth restriction, the neonates usually are more insulin-sensitive and then undergo a period of accelerated postnatal growth [61], which is associated with increased risk of developing insulin resistance and eventually type 2 diabetes [62].
4.1.3. Oxidative Stress
Oxidative stress refers to the imbalance between reactive oxygen species (ROS) and protective antioxidants. Overproduction of ROS can destroy normal placental functions. In the current study, mothers gave birth to FGR infants showed a disorder of glutathione metabolism. Glutathione, a natural body antioxidant, is a tripeptide composed of glycine, cysteine, and glutamate. Disturbance of glutathione metabolism may result in elevated oxidative products and increased oxidative stress [63], which could induce placental vascular lesions and lead to fetal compromise [9].
4.1.4. NO Synthesis Dysfunction
Nitric oxide (NO) is the main vasodilatory agent of placenta and is involved in implantation, fetoplacental vascular reactivity, and placental perfusion [64]. This gas is produced from arginine [22,28,40,41,43,47] through activation of nitric oxide synthases [65]. Arginine and proline metabolism and arginine biosynthesis were identified by pathway analysis, supporting NO synthesis dysfunction in FGR.
4.1.5. Energy Metabolism Disorder
The growth of the placenta and fetus during pregnancy increases the demand for energy [66]. The main source of cellular energy is the tricarboxylic acid cycle (TCA cycle) [67]. Glyoxylate and dicarboxylate metabolism can regulate TCA cycle via metabolites [68], such as the reported citrate [23,27,29,31,32,41] and glycine [28,29,30,31]. The identified glyoxylate and dicarboxylate metabolism pathway suggested disruptions in energy metabolism in FGR.
4.2. Reliability of Metabolic Biomarkers/Metabolite Panels
We identified some frequently reported metabolites, which strengthened our confidence in the exploration for metabolic biomarkers of FGR. Unfortunately, the change trends of some metabolites were not consistent across studies. Some of the inconsistencies may be explained by differences in biological samples, study populations, subject characteristics, analysis platforms, and other aspects. Therefore, more studies are needed to identify the metabolites involved in FGR.
Several metabolomics studies used single metabolites [19,34] or panels [22,30,31,36,38] to predict or diagnose FGR, showing decent discriminative ability. However, the application of these biomarkers in clinical practice is still insufficient. Further studies, such as cohorts with large sample sizes, are needed to obtain reliable conclusions. In addition, the combination of metabolic biomarkers with traditional predictive biomarkers or clinical parameters also needs to be explored to obtain more effective clinical models.
4.3. Limitations of Current Metabolomics Studies on FGR
Several limitations should be addressed. First, differences in study designs, diagnostic criteria of FGR, metabolomic testing platforms, and statistic methods may lead to inconsistent results, creating challenges in identifying potential metabolic biomarkers. For example, due to FGR or IUGR usually corresponding with SGA, studies on SGA were also included. Pathological differences between them may influence the results. Second, the majority of studies had a relatively small sample size, which may limit the statistical power and reliability of their results. In addition, many studies did not have validations. Before translating the results into clinical practice, multiple independent validations, including in vitro and in vivo studies, are necessary. Third, profiling of metabolites may be greatly dynamic and influenced by diet, immune status, lifestyle, and environmental factors [69]. Therefore, strict quality control is necessary for metabolomics studies. Finally, metabolomics only provides insights into the mechanisms of FGR, and it is necessary to combine other omics (e.g., genomics, transcriptomics, and proteomics) to make deeper and more comprehensive exploration of FGR.
5. Conclusions
In summary, this study presents a systematic review and analysis of metabolomics research. We identified a series of small-molecule metabolites, mainly amino acids, that were altered in FGR. The reported metabolic biomarkers and pathways suggest underlying metabolic mechanisms in the development and progression of FGR. However, current metabolomics studies on FGR are still in the preliminary development stage and more comprehensive metabolomic studies should be encouraged.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/metabo12090860/s1, Table S1: Results of the Pathway Analysis of Neonatal Studies; Table S2: Results of the Pathway Analysis of Maternal Studies.
Author Contributions
Conceptualization, J.Y.; software, M.Y. and Z.Y.; validation, X.H.; M.Z. and X.W.; data curation, X.R. and M.Y.; writing—original draft preparation, M.Y.; writing—review and editing, Z.Y., J.Y., and X.Z.; visualization, N.Y.; supervision, X.Z.; project administration, J.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Youth Program of Reinvigorating the Health through Science and Education in Suzhou, China (granted number KJXW2020054), Jiangsu Province Medical Innovation Team Program (K201105), the Natural Science Foundation of the Jiangsu Higher Education Institutions of China, the National Natural Science Foundation of China (grant: 82273635), and a Project Funded by Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nardozza, L.M.; Caetano, A.C.; Zamarian, A.C.; Mazzola, J.B.; Silva, C.P.; Marcal, V.M.; Lobo, T.F.; Peixoto, A.B.; Araujo Junior, E. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obs. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Infants with Fetal (Intrauterine) Growth Restriction. Available online: https://www.uptodate.com/contents/infants-with-fetal-intrauterine-growth-restriction#:~:text=Infants%20with%20fetal%20growth%20rstriction,with%20normal%20in%20utero%20growth (accessed on 1 September 2022).
- Deter, R.L.; Lee, W.; Sangi-Haghpeykar, H.; Tarca, A.L.; Yeo, L.; Romero, R. Fetal growth cessation in late pregnancy: Its impact on predicted size parameters used to classify small for gestational age neonates. J. Matern. Fetal Neonatal Med. 2015, 28, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Melamed, N.; Baschat, A.; Yinon, Y.; Athanasiadis, A.; Mecacci, F.; Figueras, F.; Berghella, V.; Nazareth, A.; Tahlak, M.; McIntyre, H.D.; et al. FIGO (international Federation of Gynecology and obstetrics) initiative on fetal growth: Best practice advice for screening, diagnosis, and management of fetal growth restriction. Int. J. Gynaecol. Obs. 2021, 152 (Suppl. 1), 3–57. [Google Scholar] [CrossRef]
- Society for Maternal-Fetal Medicine; Martins, J.G.; Biggio, J.R.; Abuhamad, A. Society for Maternal-Fetal Medicine Consult Series #52: Diagnosis and management of fetal growth restriction: (Replaces Clinical Guideline Number 3, April 2012). Am. J. Obs. Gynecol. 2020, 223, B2–B17. [Google Scholar] [CrossRef]
- Dessi, A.; Ottonello, G.; Fanos, V. Physiopathology of intrauterine growth retardation: From classic data to metabolomics. J. Matern.Fetal Neonatal Med. 2012, 25, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Fung, C.; Zinkhan, E. Short- and Long-Term Implications of Small for Gestational Age. Obs. Gynecol. Clin. N. Am. 2021, 48, 311–323. [Google Scholar] [CrossRef]
- Crispi, F.; Miranda, J.; Gratacos, E. Long-term cardiovascular consequences of fetal growth restriction: Biology, clinical implications, and opportunities for prevention of adult disease. Am. J. Obs. Gynecol. 2018, 218, S869–S879. [Google Scholar] [CrossRef]
- Valsamakis, G.; Kanaka-Gantenbein, C.; Malamitsi-Puchner, A.; Mastorakos, G. Causes of intrauterine growth restriction and the postnatal development of the metabolic syndrome. Ann. N. Y. Acad. Sci. 2006, 1092, 138–147. [Google Scholar] [CrossRef]
- Pallotto, E.K.; Kilbride, H.W. Perinatal outcome and later implications of intrauterine growth restriction. Clin. Obstet. Gynecol. 2006, 49, 257–269. [Google Scholar] [CrossRef]
- Robert Peter, J.; Ho, J.J.; Valliapan, J.; Sivasangari, S. Symphysial fundal height (SFH) measurement in pregnancy for detecting abnormal fetal growth. Cochrane Database Syst. Rev. 2015, CD008136. [Google Scholar] [CrossRef]
- Lees, C.C.; Stampalija, T.; Baschat, A.; da Silva Costa, F.; Ferrazzi, E.; Figueras, F.; Hecher, K.; Kingdom, J.; Poon, L.C.; Salomon, L.J.; et al. ISUOG Practice Guidelines: Diagnosis and management of small-for-gestational-age fetus and fetal growth restriction. Ultrasound Obstet. Gynecol. 2020, 56, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Shastri, S.; Farahbakhsh, N.; Sharma, P. Intrauterine growth restriction—Part 1. J. Matern. Fetal Neonatal Med. 2016, 29, 3977–3987. [Google Scholar] [CrossRef] [PubMed]
- ACOG Practice Bulletin No. 204: Fetal Growth Restriction. Obstet. Gynecol. 2019, 133, e97–e109. [CrossRef] [PubMed]
- Bujak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Kaliszan, R. Metabolomics for laboratory diagnostics. J. Pharm. Biomed. Anal. 2015, 113, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Metabolomics for Investigating Physiological and Pathophysiological Processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ Clin. Res. Ed. 2009, 339, b2700. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Moros, G.; Boutsikou, T.; Fotakis, C.; Iliodromiti, Z.; Sokou, R.; Katsila, T.; Xanthos, T.; Iacovidou, N.; Zoumpoulakis, P. Insights into intrauterine growth restriction based on maternal and umbilical cord blood metabolomics. Sci. Rep. 2021, 11, 7824. [Google Scholar] [CrossRef]
- Youssef, L.; Simoes, R.V.; Miranda, J.; Garcia-Martin, M.L.; Paules, C.; Crovetto, F.; Amigo, N.; Canellas, N.; Gratacos, E.; Crispi, F. Paired maternal and fetal metabolomics reveal a differential fingerprint in preeclampsia versus fetal growth restriction. Sci. Rep. 2021, 11, 14422. [Google Scholar] [CrossRef]
- Favretto, D.; Cosmi, E.; Ragazzi, E.; Visentin, S.; Tucci, M.; Fais, P.; Cecchetto, G.; Zanardo, V.; Viel, G.; Ferrara, S.D. Cord blood metabolomic profiling in intrauterine growth restriction. Anal. Bioanal. Chem. 2012, 402, 1109–1121. [Google Scholar] [CrossRef]
- Bahado-Singh, R.O.; Yilmaz, A.; Bisgin, H.; Turkoglu, O.; Kumar, P.; Sherman, E.; Mrazik, A.; Odibo, A.; Graham, S.F. Artificial intelligence and the analysis of multi-platform metabolomics data for the detection of intrauterine growth restriction. PLoS ONE 2019, 14, e0214121. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Simoes, R.V.; Paules, C.; Canueto, D.; Pardo-Cea, M.A.; Garcia-Martin, M.L.; Crovetto, F.; Fuertes-Martin, R.; Domenech, M.; Gomez-Roig, M.D.; et al. Metabolic profiling and targeted lipidomics reveals a disturbed lipid profile in mothers and fetuses with intrauterine growth restriction. Sci. Rep. 2018, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Cortes, M.; Carbajo, R.J.; Crispi, F.; Figueras, F.; Pineda-Lucena, A.; Gratacos, E. Metabolomic Profile of Umbilical Cord Blood Plasma from Early and Late Intrauterine Growth Restricted (IUGR) Neonates with and without Signs of Brain Vasodilation. PLoS ONE 2013, 8, e80121. [Google Scholar] [CrossRef]
- Ivorra, C.; García-Vicent, C.; Chaves, F.J.; Monleón, D.; Morales, J.M.; Lurbe, E. Metabolomic profiling in blood from umbilical cords of low birth weight newborns. J. Trans. Med. 2012, 10, 142. [Google Scholar] [CrossRef]
- Schupper, A.; Almashanu, S.; Coster, D.; Keidar, R.; Betser, M.; Sagiv, N.; Bassan, H. Metabolic biomarkers of small and large for gestational age newborns. Early Hum. Dev. 2021, 160, 105422. [Google Scholar] [CrossRef] [PubMed]
- Dessì, A.; Marincola, F.C.; Pattumelli, M.G.; Ciccarelli, S.; Corbu, S.; Ossicini, C.; Fanos, V.; Agostino, R. Investigation of the ¹H-NMR based urine metabolomic profiles of IUGR, LGA and AGA newborns on the first day of life. J. Matern. Fetal Neonatal Med. 2014, 27 (Suppl. 2), 13–19. [Google Scholar] [CrossRef]
- Chao de la Barca, J.M.; Chabrun, F.; Lefebvre, T.; Roche, O.; Huetz, N.; Blanchet, O.; Legendre, G.; Simard, G.; Reynier, P.; Gascoin, G. A Metabolomic Profiling of Intra-Uterine Growth Restriction in Placenta and Cord Blood Points to an Impairment of Lipid and Energetic Metabolism. Biomedicines 2022, 10, 1411. [Google Scholar] [CrossRef]
- Maitre, L.; Fthenou, E.; Athersuch, T.; Coen, M.; Toledano, M.B.; Holmes, E.; Kogevinas, M.; Chatzi, L.; Keun, H.C. Urinary metabolic profiles in early pregnancy are associated with preterm birth and fetal growth restriction in the Rhea mother-child cohort study. BMC Med. 2014, 12, 14. [Google Scholar] [CrossRef]
- Sulek, K.; Han, T.L.; Villas-Boas, S.G.; Wishart, D.S.; Soh, S.E.; Kwek, K.; Gluckman, P.D.; Chong, Y.S.; Kenny, L.C.; Baker, P.N. Hair Metabolomics: Identification of Fetal Compromise Provides Proof of Concept for Biomarker Discovery. Theranostics 2014, 4, 953–959. [Google Scholar] [CrossRef]
- Bahado-Singh, R.O.; Turkoglu, O.; Yilmaz, A.; Kumar, P.; Zeb, A.; Konda, S.; Sherman, E.; Kirma, J.; Allos, M.; Odibo, A.; et al. Metabolomic identification of placental alterations in fetal growth restriction. J. Matern. Fetal Neonatal Med. 2022, 35, 447–456. [Google Scholar] [CrossRef]
- Briana, D.D.; Fotakis, C.; Kontogeorgou, A.; Gavrili, S.; Georgatzi, S.; Zoumpoulakis, P.; Malamitsi-Puchner, A. Early Human-Milk Metabolome in Cases of Intrauterine Growth-Restricted and Macrosomic Infants. JPEN J. Parenter. Enter. Nutr. 2020, 44, 1510–1518. [Google Scholar] [CrossRef] [PubMed]
- Karaer, A.; Mumcu, A.; Arda, S.; Tuncay, G.; Dogan, B. Metabolomics analysis of placental tissue obtained from patients with fetal growth restriction. J. Obstet. Gynaecol. Res. 2022, 48, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Powell, K.L.; Carrozzi, A.; Stephens, A.S.; Tasevski, V.; Morris, J.M.; Ashton, A.W.; Dona, A.C. Utility of metabolic profiling of serum in the diagnosis of pregnancy complications. Placenta 2018, 66, 65–73. [Google Scholar] [CrossRef]
- Clinton, C.M.; Bain, J.R.; Muehlbauer, M.J.; Li, Y.Y.; Li, L.P.; O’Neal, S.K.; Hughes, B.L.; Cantonwine, D.E.; McElrath, T.F.; Ferguson, K.K. Non-targeted urinary metabolomics in pregnancy and associations with fetal growth restriction. Sci. Rep. 2020, 10, 5307. [Google Scholar] [CrossRef]
- Sovio, U.; Goulding, N.; McBride, N.; Cook, E.; Gaccioli, F.; Charnock-Jones, D.S.; Lawlor, D.A.; Smith, G.C.S. A maternal serum metabolite ratio predicts fetal growth restriction at term. Nat. Med. 2020, 26, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Delplancke, T.D.J.; de Seymour, J.V.; Tong, C.; Sulek, K.; Xia, Y.Y.; Zhang, H.; Han, T.L.; Baker, P.N. Analysis of sequential hair segments reflects changes in the metabolome across the trimesters of pregnancy. Sci. Rep. 2018, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Lee, S.M.; Byun, D.J.; Kim, S.Y.; Kim, H.I.; Lee, D.Y.; Jung, Y.M.; Park, C.W.; Park, J.S.; Choi, M.H. Maternal Signatures of Cortisol in First Trimester Small-for-Gestational Age. Reprod. Sci. 2022, 29, 1498–1505. [Google Scholar] [CrossRef]
- Morillon, A.C.; Leite, D.F.B.; Yakkundi, S.; Gethings, L.A.; Thomas, G.; Baker, P.N.; Kenny, L.C.; English, J.A.; McCarthy, F.P. Glycerophospholipid and detoxification pathways associated with small for gestation age pathophysiology: Discovery metabolomics analysis in the SCOPE cohort. Metabolomics 2021, 17, 5. [Google Scholar] [CrossRef]
- Alexandre-Gouabau, M.C.; Courant, F.; Le Gall, G.; Moyon, T.; Darmaun, D.; Parnet, P.; Coupe, B.; Antignac, J.P. Offspring Metabolomic Response to Maternal Protein Restriction in a Rat Model of Intrauterine Growth Restriction (IUGR). J. Proteome Res. 2011, 10, 3292–3302. [Google Scholar] [CrossRef]
- Chang, E.I.; Wesolowski, S.R.; Gilje, E.A.; Baker, P.R.; Reisz, J.A.; D’Alessandro, A.; Hay, W.W.; Rozance, P.J.; Brown, L.D. Skeletal muscle amino acid uptake is lower and alanine production is greater in late gestation intrauterine growth-restricted fetal sheep hindlimb. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R615–R629. [Google Scholar] [CrossRef]
- Huang, S.M.; Li, N.; Liu, C.; Li, T.T.; Wang, W.; Jiang, L.L.; Li, Z.; Han, D.D.; Tao, S.Y.; Wang, J.J. Characteristics of the gut microbiota colonization, inflammatory profile, and plasma metabolome in intrauterine growth restricted piglets during the first 12 hours after birth. J. Microbiol. 2019, 57, 748–758. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Liu, C.; Feng, C.; Fan, Z.; Dai, Z.; Lai, C.; Li, Z.; Wu, G.; Wang, J. Metabolomic analysis reveals differences in umbilical vein plasma metabolites between normal and growth-restricted fetal pigs during late gestation. J. Nutr. 2012, 142, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Zhang, Y.; Kinoshita, A.; Otomaru, K.; Oshima, K.; Gotoh, Y.; Oshima, I.; Sano, M.; Roh, S.; Oe, M.; et al. Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle. Metabolites 2021, 11, 582. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Zhang, Y.; Otomaru, K.; Oshima, K.; Oshima, I.; Sano, M.; Roh, S.; Ojima, K.; Gotoh, T. Maternal Nutrient Restriction Disrupts Gene Expression and Metabolites Associated with Urea Cycle, Steroid Synthesis, Glucose Homeostasis, and Glucuronidation in Fetal Calf Liver. Metabolites 2022, 12, 203. [Google Scholar] [CrossRef]
- Nissen, P.M.; Nebel, C.; Oksbjerg, N.; Bertram, H.C. Metabolomics Reveals Relationship between Plasma Inositols and Birth Weight: Possible Markers for Fetal Programming of Type 2 Diabetes. J. Biomed. Biotechnol. 2011, 2011, 378268. [Google Scholar] [CrossRef]
- Wang, Q.E.; Yue, J.; Zhou, X.; Zheng, M.H.; Cao, B.; Li, J. Ouabain regulates kidney metabolic profiling in rat offspring of intrauterine growth restriction induced by low-protein diet. Life Sci. 2020, 259, 118281. [Google Scholar] [CrossRef]
- Kim, H. Glutamine as an immunonutrient. Yonsei Med. J. 2011, 52, 892–897. [Google Scholar] [CrossRef]
- Wu, X.; Xie, C.; Zhang, Y.; Fan, Z.; Yin, Y.; Blachier, F. Glutamate-glutamine cycle and exchange in the placenta-fetus unit during late pregnancy. Amino Acids 2015, 47, 45–53. [Google Scholar] [CrossRef]
- Fernstrom, J.D.; Fernstrom, M.H. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J. Nutr. 2007, 137, 1539S–1547S; discussion 1548S. [Google Scholar] [CrossRef]
- Saibi, W.; Feki, K.; Yacoubi, I.; Brini, F. Bridging Between Proline Structure, Functions, Metabolism, and Involvement in Organism Physiology. Appl. Biochem. Biotechnol. 2015, 176, 2107–2119. [Google Scholar] [CrossRef]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched Chain Amino Acids: Beyond Nutrition Metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Kim, J.; Ko, J.W.; Choi, A.; Kwon, Y.H. Effects of maternal branched-chain amino acid and alanine supplementation on growth and biomarkers of protein metabolism in dams fed a low-protein diet and their offspring. Amino Acids 2022, 54, 977–988. [Google Scholar] [CrossRef]
- Litwack, G. Chapter 8—Glycolysis and Gluconeogenesis. In Human Biochemistry; Litwack, G., Ed.; Academic Press: Boston, MA, USA, 2018; pp. 183–198. [Google Scholar]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious Beneficial Effect of Nonessential Amino Acid, Glycine: A Review. Oxidative Med. Cell. Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Shine, A.; Hewage, C.; Malthouse, J.P.; Brindle, K.M.; McClenaghan, N.; Flatt, P.R.; Newsholme, P. A nuclear magnetic resonance-based demonstration of substantial oxidative L-alanine metabolism and L-alanine-enhanced glucose metabolism in a clonal pancreatic beta-cell line: Metabolism of L-alanine is important to the regulation of insulin secretion. Diabetes 2002, 51, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Takahashi, H.; Murao, N.; Gheni, G.; Yokoi, N.; Hamamoto, Y.; Asahara, S.I.; Seino, Y.; Kido, Y.; Seino, S. Glutamate is an essential mediator in glutamine-amplified insulin secretion. J. Diabetes Investig. 2021, 12, 920–930. [Google Scholar] [CrossRef]
- Fowden, A.L. Insulin deficiency: Effects on fetal growth and development. J. Paediatr. Child Health 1993, 29, 6–11. [Google Scholar] [CrossRef]
- Sferruzzi-Perri, A.N.; Owens, J.A.; Pringle, K.G.; Roberts, C.T. The neglected role of insulin-like growth factors in the maternal circulation regulating fetal growth. J. Physiol. 2011, 589, 7–20. [Google Scholar] [CrossRef]
- Eleftheriades, M.; Creatsas, G.; Nicolaides, K. Fetal growth restriction and postnatal development. Ann. N. Y. Acad. Sci. 2006, 1092, 319–330. [Google Scholar] [CrossRef]
- Berends, L.M.; Dearden, L.; Tung, Y.C.L.; Voshol, P.; Fernandez-Twinn, D.S.; Ozanne, S.E. Programming of central and peripheral insulin resistance by low birthweight and postnatal catch-up growth in male mice. Diabetologia 2018, 61, 2225–2234. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Xu, Z.; Cao, J.; Shao, P.; Zhou, M.; Qin, Z.; Liu, Y.; Yu, F.; Zhou, X.; Ji, W.; et al. Proteomics analysis of human placenta reveals glutathione metabolism dysfunction as the underlying pathogenesis for preeclampsia. Biochim. Biophys. Acta. Proteins Proteom. 2017, 1865, 1207–1214. [Google Scholar] [CrossRef]
- De Pace, V.; Chiossi, G.; Facchinetti, F. Clinical use of nitric oxide donors and L-arginine in obstetrics. J. Matern. Fetal Neonatal Med. 2007, 20, 569–579. [Google Scholar] [CrossRef] [PubMed]
- van Eijk, H.M.; Luiking, Y.C.; Deutz, N.E. Methods using stable isotopes to measure nitric oxide (NO) synthesis in the L-arginine/NO pathway in health and disease. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 851, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Duggleby, S.L.; Jackson, A.A. Protein, amino acid and nitrogen metabolism during pregnancy: How might the mother meet the needs of her fetus? Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Suzuki, M.; Saito, T.; Miyado, K. Emerging Role of TCA Cycle-Related Enzymes in Human Diseases. Int. J. Mol. Sci. 2021, 22, 13057. [Google Scholar] [CrossRef]
- Ma, W.; Song, J.; Wang, H.; Shi, F.; Zhou, N.; Jiang, J.; Xu, Y.; Zhang, L.; Yang, L.; Zhou, M. Chronic paradoxical sleep deprivation-induced depression-like behavior, energy metabolism and microbial changes in rats. Life Sci. 2019, 225, 88–97. [Google Scholar] [CrossRef]
- Souza, R.T.; Mayrink, J.; Leite, D.F.; Costa, M.L.; Calderon, I.M.; Rocha Filho, E.A.; Vettorazzi, J.; Feitosa, F.E.; Cecatti, J.G. Metabolomics applied to maternal and perinatal health: A review of new frontiers with a translation potential. Clinics 2019, 74, e894. [Google Scholar] [CrossRef]
Figure 1.
PRISMA flowchart of study selection.

Figure 2.
Numbers of the human neonatal and maternal studies according to (a) bio-specimen, (b) untargeted/targeted, (c) analytical platform, and (d) sample size.
Figure 2.
Numbers of the human neonatal and maternal studies according to (a) bio-specimen, (b) untargeted/targeted, (c) analytical platform, and (d) sample size.

Figure 3.
Overview of pathway analysis of human (a) neonatal and (b) maternal studies. The x-axis is the pathway impact value calculated based on topology analysis, while the y-axis indicates the significance level in enrichment analysis. Each circle in the figure represents a metabolic pathway. The circle size reflects the pathway impact value. And the blue circle indicates the pathway is significantly enriched (FDR < 0.05) and has relatively high impact value.
Figure 3.
Overview of pathway analysis of human (a) neonatal and (b) maternal studies. The x-axis is the pathway impact value calculated based on topology analysis, while the y-axis indicates the significance level in enrichment analysis. Each circle in the figure represents a metabolic pathway. The circle size reflects the pathway impact value. And the blue circle indicates the pathway is significantly enriched (FDR < 0.05) and has relatively high impact value.

Table 1.
Characteristics of the Human Neonatal Studies.
| Ref | Case Sample Size | Control Sample Size | Diagnostic Criteria | Bio-Specimen | Sampling Time | Analytic Platform | Targeted/ Untargeted | Upregulated | Downregulated |
|---|---|---|---|---|---|---|---|---|---|
| Georgios Moros (2021) [19] | 41 | 36 | BW < 10th customized centile and abnormal umbilical artery RI | Umbilical cord serum | Not mentioned | 1H NMR | Untargeted | Alanine; Leucine; Isoleucine; Valine | Phenylalanine; Glycerol; Tryptophan |
| Lina Youssef (2021) [20] | 43 | 86 | EFW and BW < 10th centile and abnormal cerebroplacental ratio or abnormal uterine artery pulsatility index, or BW < 3rd centile | Umbilical cord plasma | Immediately after delivery | 1H NMR | Untargeted | Triglycerides (IDL); Cholesterol (IDL) | Cholesterol (HDL); Isoleucine |
| Donata Favretto (2012) [21] | 22 | 21 | EFW < 10th percentile | Umbilical cord serum | Immediately after delivery | LC-HRMS | Untargeted | Valine; Isoleucine; Glutamate; Methionine; Dopamine; Histidine; Proline; Phenylalanine; Uric acid; Caffeine; 5-Methyl-2-undecenoic acid; Tryptophan; Kynurenine; Leu pro; Thyronine; Oleic acid; Hexadecanedioic acid; Phe phe; Arg cys asn; Arg phe arg; Trp arg arg; 1-Hydroxyvitamin D3 3-dglucopyranoside | None |
| Ray Oliver Bahado-Singh (2019) [22] | 39 | 39 | BW < 10th percentile | Umbilical cord serum | Within 20 min of delivery | LC-MS/MS 1H NMR | Untargeted | Lysine; Threonine; Dopa; Kynurenine; LysoPC a C18:1; LysoPC a C20:3; PC aa C36:1; PC aa C36:3; PC aa C38:3; PC ae C44:5; Arginine | Creatinine; C0; C10:1; C12:1; C16:1; C2; C4; C4-OH; PC aa C24.0; PC aa C26.0; PC aa C28:1; PC aa C30:0; PC aa C32:0; PC aa C32:1; PC aa C34:2; PC aa C36:4; PC aa C36:6; PC aa C38:4; PC aa C38:5; PC aa C40:4; PC aa C40:6; PC aa C42:2; PC aa C42:4; PC aa C42:5; PC aa C42:6; PC ae C30:2; PC ae C36:0; PC ae C36:3; PC ae C36:5; PC ae C38:4; PC ae C38:6; PC ae C42:1; 2-Hydroxybutyrate; 3-Hydroxybutyrate; Acetate; Acetoacetate; Choline; Creatine; Formate |
| Jezid Miranda (2018) [23] | 27 | 28 | BW < 3rd centile and/or abnormal uterine artery doppler and/or abnormal cerebroplacental ratio | Umbilical cord plasma | Immediately after delivery | 1H NMR | Untargeted +targeted | Cholesterol (VLDL); Triglycerides (VLDL); Triglycerides (IDL); Acetate; Formate | None |
| Magdalena Sanz-Cortés (2013) [24] | 20 | 23 | BW < 10th centile and abnormal umbilical artery | Umbilical cord serum | At delivery | 1H NMR | Untargeted | Triglycerides; Creatine; Glutamine (Early-onset FGR) | Glucose; Choline; Phenylalanine (Early-onset FGR) Choline; Glutamine; Tyrosine; Valine; Leucine; Alanine (Late-onset FGR) |
| Carmen Ivorra (2012) [25] | 20 | 30 | BW < 10th percentile | Umbilical cord plasma | Immediately after delivery | NMR | Untargeted | Citrulline; Phenylalanine | Proline; Choline; Glutamine; Alanine; Glucose |
| Aviv Schupper (2021) [26] | 6380 | 61,068 | BW < 10th percentile | Neonatal dried blood | Between 36 and 72 h from birth | HPLC-MS | Targeted | Alanine; Methionine; Proline; Carnitine | Valine |
| Angelica Dessı` (2014) [27] | 12 | 17 | Ultrasonographically in the prenatal period and BW < 10th percentile | Neonatal urine | Within 8 h of delivery | 1H NMR | Untargeted | Citrate; Creatinine; Creatine; Myo-inositol; Betaine/Trimethylamine N-oxide; Glycine | None |
| Juan Manuel Chao de la Barca (2022) [28] | 15 | 15 | BW < 10th percentile and doppler abnormalities | Umbilical cord plasma | Not mentioned | LC-MS/MS FIA-MS/MS | Targeted | C0; C4; C2; Alanine; Asparagine; Tyrosine; Glutamine; Proline; Alpha-aminoadipic acid; Trans-4-hydroxyproline; Spermine; LysoPC a C26:1; PC aa C32:0; PC aa C24:0 | LysoPC a C18:0; LysoPC a C17:0; LysoPC a C18:1; LysoPC a C20:3; LysoPC a C18:2; LysoPC a C20:4; LysoPC a C16:1; LysoPC a C16:0; PC ae C40:6; PC ae C42:5; PC aa C38:6; PC aa C36:0; PC aa C42:0; PC aa C42:6; PC ae C42:4; PC ae C40:2; PC ae C38:6; PC aa C36:1; PC ae C40:3; PC ae C40:5; PC ae C40:4; PC aa C36:3; PC ae C42:2; PC ae C40:1; PC aa C40:6; PC ae C42:3; PC ae C44:5; PC aa C38:0; PC ae C38:0; PC aa C36:6; PC aa C38:3; PC ae C44:4; PC ae C38:3; PC ae C44:6; PC ae C36:3; SM(OH)C22:1; SM(OH)C24:1; SM C24:0; SM C26:0; Tryptophan |
BW, birth weight; RI, resistance indices; EFW, estimated fetal weight; IDL, intermediate density lipoprotein; HDL, high density lipoprotein; VLDL, very low density lipoprotein.
Table 3.
Characteristics of the Human Maternal Studies.
| Ref | Case Sample Size | Control Sample Size | Diagnostic Criteria | Bio-Specimen | Sampling Time | Analytic Platform | Targeted/ Untargeted | Upregulated | Downregulated |
|---|---|---|---|---|---|---|---|---|---|
| Georgios Moros (2021) [19] | 41 | 36 | BW < 10th customized centile and abnormal umbilical artery RI | Maternal serum | During delivery | 1H NMR | Untargeted | Alanine; Leucine; Isoleucine; Valine | Phenylalanine; Glycerol; 3-Hydroxybutyrate |
| Chelsea M. Clinton (2020) [35] | 30 | 30 | BW < 10th percentile | Urine | 10 w | GC-MS | Untargeted | Benzoic acid; Malonic acid; 2-Ketoleucine/ketoisoleucine; 2-Ketobutyric acid; 2-Methylglutaric acid; Acetoacetate | None |
| 26 w | 1,2-Propanediol; Kynurenic acid; N-heptanoic acid; Benzoic acid | None | |||||||
| Léa Maitre (2014) [29] | 36 | 275 | BW < 10th percentile of predicted BW distribution | Urine | 11–13 w | 1H NMR | Untargeted | None | Tyrosine; Lactate; Alanine; Acetate; Citrate; Trimethylamine; Glycine; Formate |
| Ulla Sovio (2020) [36] | 175 | 299 | BW < 3rd percentile, or BW between the 3rd and 10th percentile and the lowest decile of fetal abdominal growth velocity | Maternal serum | 20/28/36 w | UPLC-MS/MS | Untargeted | 1-(1-Enyl-stearoyl)2-oleoyl-GPC (P-18:0/18:1); 1,5-Anhydroglucitol; Cotinine N-oxide; 4-Androsten-3beta,17beta-diol monosulfate; Hydroxycotinine; Acisoga; 3-Hydroxycotinine glucuronide; O-cresol sulfate; Dehydroisoandrosterone sulfate | 5Alpha-androstan-3Alpha,17alpha-diol disulfate; Estriol 3-sulfate; 4-Cholesten-3-one; Pregnanolone/allopregnanolone sulfate; 5Alpha-pregnan-3alpha,20beta-diol disulfate 1; N1,N12-diacetylspermine; 17Alpha-hydroxypregnanolone glucuronide; 5Alpha-pregnan-3beta,20beta-diol monosulfate; Progesterone; Pregnanediol-3-glucuronide |
| Karolina Sulek (2014) [30] | 41 | 42 | BW < 10th corrected birth centile | Maternal hair | 26–28 w | GC-MS | Untargeted | Palmitate; 2-Methyloctadecanoate; Myristate; Margarate; Stearate; Dodecanoate; Octanoate; Heptadecane; Nicotinamide | 3-Hydroxybenzoate; Levulinic acid; 1-Aminocyclopropane-1-carboxylate; Citraconate; Lactate; Glycine; Proline; Isoleucine; Serine; Leucine; Glutamate; Phenylalanine; Alanine; Valine; Aspartate; Threonine; Tyrosine; Methionine; Lysine; Pyroglutamate; Ornithine; Glutathione |
| Jezid Miranda (2018) [23] | 27 | 28 | BW < 3rd centile and/or abnormal uterine artery doppler and/or abnormal cerebroplacental ratio | Maternal plasma | 2–4 h after delivery | 1H NMR | Untargeted + targeted | None | Triglycerides (HDL); Alanine; Citrate; 2-oxoisovaleric acid; Pyruvate |
| Ray Oliver Bahado-Singh (2020) [31] | 19 | 30 | BW < 10th percentile | Placenta | Within 20 min of delivery | DI-LC-MS/MS 1H NMR | Untargeted | Citrate | 3-Hydroxybutyrate; Glycine; D-Glucose; t4-OH-Pro; Symmetric dimethylarginine; Sarcosine; Kynurenine; Methionine sulfoxide; Alanine; Aspartate; Spermidine; Valine; PC ae C424; Myo-inositol |
| Despina D. Briana (2020) [32] | 19 | 60 | BW ≤ 10th customized percentile | Human-milk | Within 3–4 d of delivery | 1H NMR | Untargeted | Citrate; Choline; Lactose; N-acetylglutamine; Phosphocoline | Isoleucine; Valine |
| Thibaut D. J. Delplancke (2018) [37] | 20 | 73 | BW < 10th customized centile | Hair | In the second trimester | GC-MS LC-MS | Untargeted | Myristic acid; Pentadecanoic acid; Margaric acid | None |
| Abdullah Karaer (2022) [33] | 10 | 14 | AC/EFW ratio < 3rd percentile | Placenta | Within 5 min of delivery | 1H HR-MAS NMR | Untargeted | Lactate; Glutamine; Glycerophosphocholine; Phosphocholine; Taurine; Myo-inositol | None |
| Chaelin Lee (2022) [38] | 56 | 56 | BW < 10th percentile | Maternal serum | 10–14 w | LC-MS | Targeted | Tetrahydrocortisol | 21-Deoxycortisol |
| Aude-Claire Morillon (2021) [39] | 40 | 40 | BW < 10th percentile | Urine | 20 w | UPLC-MS | Untargeted | None | Sulfolithocholic acid; Estriol-16-glucuronide; 4-Hydroxybenzaldehyde; Neuromedin N; D-Glucuronic acid; 18-Hydroxycortisol; Beta-1,4-Mannosyl-N-acetylglucosamine |
| 40 | 40 | Maternal serum | PE(P-31:0); PE(42:1); PE(36:4); PS(O-37:0); PS(41:5); PS(37:2); PS(43:6); PS(P-34:0); PC(O-42:4); PC(40:5); PC(38:6); LysoPC(16:0); PA(O-36:2); LysoPA(18:1); PI(37:1); PI(P-33:1); PGP(38:4); PGP(40:4); PG(36:6); PG(39:8); PG(38:4); DG(44:4); DG(O-34:1); N,N-dimethyl arachidonoyl amine; N-palmitoyl valine; SM(34:1) Ganglioside GA2 (40:1); Cer(34:0); Cer(39:2) | CL(72:2); TG(64:15); 8S-hydroxy-hexadecanoic acid; CE(17:0) | |||||
| Katie L. Powel (2018) [34] | 34 | 82 | EFW < 10th percentile and abnormal placental vascular resistance | Maternal serum | 26–41 w | 1H NMR | Untargeted | Glutamate; Glutamine | 3-Hydroxybutyrate |
| Juan Manuel Chao de la Barca (2022) [28] | 20 + 24 | 20 + 22 | BW < 10th percentile and doppler abnormalities | Placenta | Within 30 min after delivery | LC-MS/MS FIA-MS/MS | Targeted | C0; C2; C4; C4-OH; C16; C18; C18:1; C5; C16:1; C18:2; Tryptophan; Creatinine; PC ae C36:5; Hexose | Aspartate; Threonine; Kynurenine; Putrescine; Spermidine; Trans-4-hydroxyproline; LysoPC a C18:0; LysoPC a C20:3; LysoPC a C20:4; PC aa C36:1; PC aa C38:0; PC aa C40:3; PC ae C42:3; SM C24:0 SM C26:0; SM C24:1; SM (OH) C22:1; Glycine; Serine; Arginine; Tyrosine; Alpha-aminoadipic acid; LysoPC a C16; LysoPC a C16:1; LysoPC a C17:0; LysoPC a C18:1; LysoPC a C18:2; PC aa C32:0; PC ae C34:0; PC ae C40:2; PC ae C44:5; SM C26:1; SM(OH) C22:2; SM(OH) C24:1; Methionine; Carnosine; LysoPC a C24:0; LysoPC a C28:1; PC aa C36:3; PC aa C38:3; PC aa C42:0; PC ae C30:0; PC ae C30:2; PC ae C36:1; PC ae C36:2; PC ae C36:3; PC ae C38:2; PC ae C38:3; PC ae C40:3; PC ae C40:4; PC ae C42:4; PC ae C42:6; SM C16:1; SM(OH) C14:1 |
RI, resistance indices; BW, birth weight; HDL, high density lipoprotein; AC, abdominal circumference; EFW, estimated fetal weight.
Table 4.
The Potential of Metabolic Markers for the Prediction and Diagnosis of FGR.
| Ref | Bio-Specimen | Potential Biomarkers | Sensitivity | Specificity | AUC |
|---|---|---|---|---|---|
| Georgios Moros (2021) [19] | Maternal and umbilical cord blood | Alanine | - | - | 0.871 (Umbilical cord blood) 0.792 (Maternal blood) |
| Isoleucine | - | - | 0.795 (Umbilical cord blood) 0.812 (Maternal blood) | ||
| Leucine | - | - | 0.816 (Umbilical cord blood) 0.773 (Maternal blood) | ||
| Valine | - | - | 0.785 (Umbilical cord blood) 0.786 (Maternal blood) | ||
| Phenylalanine | - | - | 0.779 (Umbilical cord blood) 0.75 (Maternal blood) | ||
| Glycerol | - | - | 0.853 (Umbilical cord blood) 0.751 (Maternal blood) | ||
| Tryptophan | - | - | 0.751 (Umbilical cord blood) | ||
| 3-Hydroxybutyrate | - | - | 0.774 (Maternal blood) | ||
| Ulla Sovio (2020) [36] | Maternal serum | (A)1-(1-Enyl-stearoyl)-2-oleoyl-GPC (P-18:0/18:1) | - | - | 0.640 |
| (B)1,5-Anhydroglucitol | - | - | 0.650 | ||
| (C)5α-Androstan-3α,17α-diol disulfate | - | - | 0.690 | ||
| (D)N1,N12-diacetylspermine | - | - | 0.660 | ||
| A × B | - | - | 0.700 | ||
| C × D | - | - | 0.710 | ||
| (A × B)/ (C × D) | - | - | 0.780 | ||
| Karolina Sulek (2014) [30] | Maternal hair | Lactate + levulinate + 2-methyloctadecanate + tyrosine + margarate | - | - | 0.998 |
| Ray O. Bahado-Singh (2020) [31] | Placenta | 3-Hydroxybutyric acid + glycine + PC aa C420 | 0.867 | 0.842 | 0.912 |
| LysoPC a C261 + PC ae C382 + PC ae C446 | 0.867 | 0.737 | 0.796 | ||
| 3-Hydroxybutyric acid + glycine + PC aa C420 + medical disorder + maternal age + prior FGR + race + gravidity | 0.833 | 0.789 | 0.832 | ||
| Ray Oliver Bahado-Singh (2019) [22] | Cord blood serum | Creatinine + C14 + C2 + C4 + LysoPC.a.C16:1 + LysoPC.a.C18:1 + LysoPC.a.C20:3 + LysoPC.a.C20:4 + LysoPC.a.C28:1 + PC.aa.C24:0 + PC.aa.C36:4 + PC.aa.C38:4 + PC.aa.C42:4 + EDTAca_N + creatine | 0.870 | 0.830 | 0.910 |
| LysoPC.a.C16:1 + C2 + creatinine + LysoPC.a.C18:2 + LysoPC.a.C18:1 + LysoPC.a.C20:3 + PC.aa.C24:0 + C6.C4:1.DC. + C4 + C10:1 + C16:1 + C12:1 + C12 + C0 + LysoPC.a.C28:1 | 0.830 | 0.850 | 0.870 | ||
| Creatinine + LysoPC.a.C16:1 + LysoPC.a.C20:3 + C2 + LysoPC.a.C18:2 + C4 + C12:1 + EDTAca_N + C6.C4:1.DC. + taurine + C16:2 + C0 + putrescine + PC.aa.C24:0 + LysoPC.a.C28:1 | 0.850 | 0.790 | 0.880 | ||
| Creatinine + C2 + C4 + LysoPC.a.C16:1 + LysoPC.a.C2:.3 + LysoPC.a.C28:1 + PC.aa.C24:0 | 0.830 | 0.870 | 0.880 | ||
| Creatinine + C2 + C4 + LysoPC.a.C16:1 + LysoPC.a.C2:.3 + LysoPC.a.C28:1 + PC.aa.C24:0 + prior FGR + maternal age + gravity + race | 0.700 | 0.730 | 0.690 | ||
| Chaelin Lee (2022) [38] | Maternal serum | 21-DeoxyF + F/21-deoxyF(cortisol to 21-deoxycortisol) + THF/F(tetrahydrocortisol to cortisol) | 0.734 | 0.817 | 0.824 |
| Katie L. Powel (2018) [34] | Placenta | 3-Hydroxybutyrate | 0.939 0.364 | 0.300 0.545 | 0.623 (Discovery) 0.581 (Validation) |
Table 5.
Characteristics of the Animal Studies.
| Ref | Case Sample Size | Control Sample Size | FGR Model | Bio-Specimen | Sampling Time | Analytic Platform | Targeted/ Untargeted | Upregulated | Downregulated |
|---|---|---|---|---|---|---|---|---|---|
| Alexandre-Gouabau (2011) [40] | 4 | 10 | Fetal rats of low protein diet maternal rats | Fetal plasma | Postnatal 0 d | LC-HRMS | Untargeted | None | Proline; Arginine; Histidine |
| Eileen I. Chang (2019) [41] | 10 | 8 | Fetal sheep of placental insufficiency | Fetal skeletal muscle | Late gestation | HPLC | Untargeted | 2-Oxo-7-methylthioheptanoic acid; 2-Hydroxyglutarate; N-acetylneuraminate | Cystathionine; N-acetylcitrulline; Gamma-glutamylalanine; N-carbamyl-L-glutamate; Gamma-glutamylcysteine; Arginine; Aspartate; N,N-dimethylglycine; Adenosine; 5-Hydroxylysine; 4-Hydroxyproline |
| Fetal arterial and venous plasma | 4-Pyridoxate; 2-Aminoadipate; Triacanthine; Dopamine; Uric Acid; Citrate; Adrenaline; Carnitine; 5,6-Dihydrothymine; Arabitol; Guanine; Taurine; Dehydroascorbate; Alanine | Orthophosphate; Diphosphate; 3D-(3,5/4)-trihydroxycyclohexane-1,2-dione; Fructose 1,6-bisphosphate; 5-Hydroxylysine; 2-Methyleneglutarate; N-Acyl-D-mannosaminolactone; Pyruvate; 1,4-Beta-D-xylan; Glucose; Cys gly; Ascorbate; Arginine; Sn-glycerol 3-phosphate; Succinate; Ribose | |||||||
| Shimeng Huang (2019) [42] | 6 | 6 | Piglets with a birth weight 2 sd below the mean | Fetal plasma | Immediately after birth | UPLC-MS | Untargeted | 6’-Sialyllactose; Faradiol; Neotigogenin; Zymosterol intermediate 2, 13, 14-Dihydro PGF-1 alpha; 6, 10, 14-Trimethyl-5, 9, 13-pentadecatrien-2-one | Serotonin; Dihydrowyerone |
| Gang Lin (2012) [43] | 9 | 9 | Piglets with a birth weight 2 sd below the mean | Umbilical vein plasma | 90 d of gestation | HPLC-Q-TOFMS | Untargeted | Pyroglutamic acid | Proline; Valine; Phenylalanine; Isoleucine; Leucine; Tyrosine; Glutamine; Arginine; Dodecanoylcarnitine |
| 9 | 9 | 110 d of gestation | Creatinine; Carnitine; Pyroglutamic acid | Tryptophan; Glutamine; Arginine; Dodecanoylcarnitine | |||||
| Susumu Muroya (2021) [44] | 4 | 4 | Calves of maternal undernutrition | Fetal muscle | 260 ± 8.3 d of gestation | CE-TOFMS | Untargeted | Carnosine; Glutamine; Glycerol; Creatine; N6-methyllysine; Phosphorylcholine; Phenylalanine; Proline | Myo-inositol 2-phosphate; 2-Aminoethylphosphonic acid |
| Susumu Muroya (2022) [45] | 4 | 4 | Calves of maternal undernutrition | Fetal liver | 260 ± 8.3 d of gestation | CE-TOFMS | Untargeted | Aspartate; Betaine aldehyde; Glycerol; 3-Aminopropane1,2-diol; Alanine; 6-Phosphogluconate; Ophthalmate | 4-Amino-3-hydroxybutyrate; 2-Aminoethylphosphonate; UDP-glucose/UDP-galactose; N5-Ethylglutamine; UDP-glucuronate; 2-Hydroxybutyrate; Octanoate; Gly leu |
| PiaMarlene Nissen (2011) [46] | 12 | 12 | Piglets with low birth weight | Umbilical cord plasma | 110 d of gestation | GC-MS | Untargeted | Myoinositol; D-chiro-inositol | None |
| Qien Wang (2020) [47] | 53 | 57 | Fetal rats of low protein diet maternal rats | Fetal kidney | 20 d of gestation | GC-MS LC-MS | Untargeted | Xanthine; Xanthosine; L-lysine; Oleate; Arachidonate; Phenylalanine; Glutamine; Guanine | N-acetylornithine; Arginine; GMP; UMP; Acetyl-CoA; Carbamoyl phosphate; dAMP; UDP; TMP; dTDP; Urea; Pantothenate |
Table 6.
High Frequency Metabolic Biomarkers of Animal Studies.
| No | Metabolites | Hits | Up | Down |
|---|---|---|---|---|
| 1 | Arginine | 4 | - | Fetal rats plasma [40] Fetal sheep skeletal muscle+ arterial and venous plasma [41] Piglets umbilical vein plasma [43] Fetal rats kidney [47] |
| 2 | Glutamine | 3 | Calves muscle [44] Fetal rats kidney [47] | Piglets umbilical vein plasma [43] |
| 3 | Phenylalanine | 3 | Calves muscle [44] Fetal rats kidney [47] | Piglets umbilical vein plasma [43] |
| 4 | Proline | 3 | Calves muscle [44] | Fetal rats plasma [40] Piglets umbilical vein plasma [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yao, M.; Yang, Z.; Rong, X.; Hu, X.; Yao, N.; Zhu, M.; Wang, X.; Zhu, X.; Yin, J. The Exploration of Fetal Growth Restriction Based on Metabolomics: A Systematic Review. Metabolites 2022, 12, 860. https://doi.org/10.3390/metabo12090860
AMA Style
Yao M, Yang Z, Rong X, Hu X, Yao N, Zhu M, Wang X, Zhu X, Yin J. The Exploration of Fetal Growth Restriction Based on Metabolomics: A Systematic Review. Metabolites. 2022; 12(9):860. https://doi.org/10.3390/metabo12090860
Chicago/Turabian StyleYao, Mengxin, Zhuoqiao Yang, Xin Rong, Xuan Hu, Na Yao, Manting Zhu, Xinnan Wang, Xiaoyan Zhu, and Jieyun Yin. 2022. "The Exploration of Fetal Growth Restriction Based on Metabolomics: A Systematic Review" Metabolites 12, no. 9: 860. https://doi.org/10.3390/metabo12090860
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.