One-Year Effects of Omega-3 Treatment on Fatty Acids, Oxylipins, and Related Bioactive Lipids and Their Associations with Clinical Lipid and Inflammatory Biomarkers: Findings from a Substudy of the Vitamin D and Omega-3 Trial (VITAL)

 , , , , , , add
Show full author list
, , , , , , add
Show full author list
Abstract
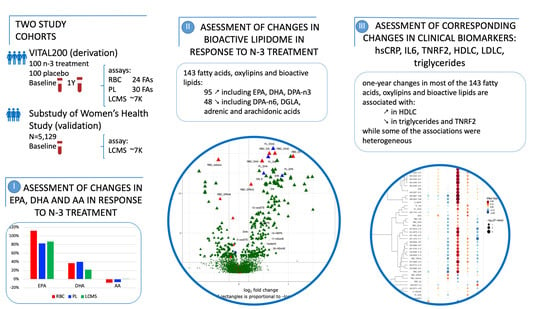
:
1. Introduction
2. Results
2.1. Baseline Characteristics
2.2. Baseline Levels of EPA, DHA and AA Were Consistent across the Three Assays
2.3. Baseline Levels of EPA, DHA and AA Were Consistent Across the Three Assays and in the n-3 Treatment and the Placebo Groups
2.4. One Year of n-3 Treatment Lead to Changes of EPA, DHA and AA Levels in the n-3 Treatment vs the Placebo Group
2.5. One Year of n-3 Treatment Lead to Significant Changes in 143 FAs, Oxylipins or Small Bioactive Lipid Features
2.6. Validation in the Women’s Health Study (WHS)
2.7. Associations of One-Year Changes in FAs, Oxylipins and Bioactive Lipids with One-year Changes in Six Downstream Clinical Biomarkers hsCRP, IL6, TNRF2, HDLC, LDLC, and Triglycerides
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Biomarker Profiling
4.3. Statistical Analysis
4.3.1. Association with n-3 Treatment
4.3.2. Associations of One-Year Randomized Changes in FAs, Oxylipins or Small Bioactive Lipid Features with One-year Changes in Downstream Clinical Lipid and Inflammatory Biomarkers
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Meaning |
| AA | arachidonic acid |
| adrenic acid | cis ω6 polyunsaturated (22:4) or 7,10,13,16-docosatetraenoic acid |
| CHD | coronary heart disease |
| CVD | cardiovascular disease |
| DGLA | dihomo gamma linolenic |
| DHA | docosahexaenoic acid |
| DPA-n6 | docosapentaenoic n-6 acid |
| EPA | eicosapentaenoic acid |
| FA | fatty acid |
| FDR | False Discovery Rate |
| WHS | Women’s Health Study |
| GC | gas chromatography |
| RBC | gas chromatography-mass spectrometry with flame ionization decoupling in red blood cells |
| LC | liquid chromatography |
| LCMS | high-throughput liquid chromatography-mass spectrometry of circulating free FAs, oxylipins and bioactive lipids from plasma |
| plasma PL | LCMS2 plasma phospholipid assay |
| MI | myocardial infarction |
| MS | mass spectrometry |
| n-3 | omega-3 |
| n-6 | omega-6 |
| o3:6i | omega-3 to omega-6 index (=[EPA + DHA]/AA) |
| o3i | omega-3 index (=EPA + DHA) |
| SD | standard deviation |
| VITAL | Vitamin D and Omega-3 Trial |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| abs. Level at bl | abs. Change Y1−bl | % Change Y1−bl | ||||
|---|---|---|---|---|---|---|
| Placebo | n−3 Treatment | Placebo | n−3 Treatment | Placebo | n−3 Treatment | |
| RBC Assay | ||||||
| N | 84 | 75 | 74 | 63 | 74 | 63 |
| EPA | 0.5 (0.4:0.7) ‡ | 0.5 (0.4:0.7) | 0 (−0.1:0.1) | 0.7 (0.2:1.1) | −0.9 (−17.4:19) *** | 111.8 (45.4:237.7) *** |
| DHA | 4.7 (3.7:5.5) | 4.6 (4.1:5.2) | −0.1 (−0.4:0.3) | 1.7 (0.9:2.4) | −1.2 (−9.6:5.9) *** | 36.8 (17.2:56.3) *** |
| AA | 17.4 (16.2:18.3) | 17.3 (16.2:18.4) | 0 (−0.6:0.6) | −1.3 (−2.4:−0.2) | 0.3 (−3.4:3.6) *** | −7.7 (−12.6:−1.4) *** |
| o3i | 5.3 (4.2:6) | 5.1 (4.5:5.9) | −0.1 (−0.5:0.3) | 2.3 (1.2:3.2) | −1.7 (−10.4:5.5) *** | 50.7 (22.9:67.2) *** |
| o3:6i | 0.3 (0.2:0.4) | 0.3 (0.3:0.4) | 0 (−0.02−0.02) | 0.2 (0.1:0.2) | −1.7 (−8.7:7.4) *** | 62.8 (26.8:92.1) *** |
| Plasma PL Assay | ||||||
| N | 95 | 95 | 94 | 94 | 94 | 94 |
| EPA | 0.5 (0.4:0.6) | 0.5 (0.4:0.8) | 0 (−0.1:0.1) | 0.5 (0.1:1) | 0 (−20:33.3) *** | 83.3 (25:220) |
| DHA | 2.2 (1.8:2.6) | 2.1 (1.7:2.7) | 0 (−0.4:0.2) | 0.8 (0.3:1.2) | −1.4 (−16.7:13.3) *** | 39.6 (14:62.8) *** |
| AA | 10.2 (8.4:12.2) | 10.3 (8.6:11.9) | −0.2 (−1.1:1) | −0.7 (−1.6:0.2) | −2.5 (−10.1:10.2) ** | −7 (−15.1:2.7) ** |
| o3i | 2.8 (2.2:3.2) | 2.7 (2.2:3.6) | 0 (−0.4:0.3) | 1.3 (0.4:2.1) | 0 (−14.8:15.6) *** | 51 (18.8:80.9) *** |
| o3:6i | 0.2 (0.2:0.3) | 0.3 (0.2:0.3) | 0 (−0.02−0.03) | 0.2 (0:0.3) | 1.3 (−10.9:10.7) *** | 63.9 (15.9:110.7) *** |
| LCMS Assay | ||||||
| N | 100 | 99 | 100 | 98 | 100 | 98 |
| EPA | 0.5 (0.3:0.7) | 0.5 (0.3:0.8) | 0 (−0.2:0.2) | 0.4 (0.2:0.7) | −2.3 (−36.9:34) *** | 86.5 (31.5:182) *** |
| DHA | 2.2 (1.8:2.5) | 2.1 (1.8:2.7) | 0 (−0.3:0.4) | 0.5 (0:1) | 2.6 (−14.8:17.5) *** | 21.7 (1.4:44.4) *** |
| AA | 10.4 (9.1:12.1) | 10 (8.4:12.1) | 0.1 (−1.4:1.2) | 0.1 (−1.2:1) | 0.9 (−12.9:13.5) | 1.4 (−10.5:11.5) |
| o3i | 2.7 (2.2:3.1) | 2.6 (2.2:3.4) | 0 (−0.5:0.5) | 1 (0.3:1.7) | 0.6 (−17.7:21.2) *** | 36.6 (12:78.1) *** |
| o3:6i | 0.3 (0.2:0.3) | 0.3 (0.2:0.3) | 0 (−0.04−0.04) | 0.1 (0:0.2) | 1.5 (−12.5:15) *** | 39.6 (16.1:74.3) *** |
| Annotation or m/z RT | Assay | Median % Change | IQR |
|---|---|---|---|
| 528.2974 5.38 | LCMS | −29 | (−52–20) |
| 475.2657 4.13 | LCMS | −26 | (−47–18) |
| 474.2627 4.13 | LCMS | −26 | (−47–19) |
| RBC_DPAn6 | RBC | −25 | (−39−10) |
| RBC_Adrenic | RBC | −24 | (−32–7) |
| 502.2943 5.62 | LCMS | −24 | (−44–5) |
| 451.2623 4.42 | LCMS | −20 | (−47–6) |
| 450.2628 4.43 | LCMS | −20 | (−47–7) |
| 532.2654 4.43 | LCMS | −19 | (−46–9) |
| 604.3627 5.07 | LCMS | −17 | (−40–2) |
| 561.3002 5.73 | LCMS | −13 | (−32–5) |
| 502.2874 4.59 | LCMS | −12 | (−24–5) |
| 500.2787 4.62 | LCMS | −12 | (−25–5) |
| 600.2039 4.8 | LCMS | −11 | (−22–5) |
| 501.2818 4.61 | LCMS | −11 | (−25–4) |
| 503.2859 4.8 | LCMS | −10 | (−28–2) |
| 502.2846 4.8 | LCMS | −10 | (−26–0) |
| 500.2794 4.8 | LCMS | −10 | (−24–0) |
| 501.2821 4.8 | LCMS | −9 | (−23–1) |
| 441.2109 2.3 | LCMS | −9 | (−25–5) |
| RBC_AA | RBC | −8 | (−13−1) |
| 583.2847 4.8 | LCMS | −6 | (−16–0) |
| RBC_DGLA | RBC | −6 | (−11–2) |
| 582.2814 4.8 | LCMS | −5 | (−16–2) |
| RBC_DPAn3 | RBC | 17 | (5–34) |
| oxylipin1 | LCMS | 17 | (−3–59) |
| 527.286 4.69 | LCMS | 17 | (−3–53) |
| 326.2216 6.28 | LCMS | 20 | (−10–82) |
| 5-oxoETE | LCMS | 23 | (−11–66) |
| DHA | LCMS | 24 | (2–49) |
| 364.2088 6.45 | LCMS | 28 | (2–77) |
| 363.209 6.44 | LCMS | 30 | (1–78) |
| 17-HDoHE | LCMS | 30 | (−6–113) |
| 444.1595 2.2 | LCMS | 30 | (−8–85) |
| 365.2064 6.44 | LCMS | 33 | (−1–87) |
| RBC_DHA | RBC | 37 | (17–56) |
| 329.1724 4.91 | LCMS | 37 | (−6–96) |
| 325.218 6.28 | LCMS | 39 | (−5–99) |
| PL_DHA | PL | 40 | (14–63) |
| 12-HEPE | LCMS | 43 | (4–127) |
| oxylipin2 | LCMS | 45 | (−5–130) |
| 247.1704 5 | LCMS | 47 | (−2–139) |
| 5-HEPE | LCMS | 50 | (9–130) |
| 247.1705 4.91 | LCMS | 50 | (0–150) |
| RBC_O3i | RBC | 51 | (23–67) |
| PL_O3i | PL | 51 | (19–81) |
| EPA | LCMS | 61 | (24–131) |
| RBC_O3:6i | RBC | 63 | (27–92) |
| PL_O3:6i | PL | 64 | (16–111) |
| PL_EPA | PL | 83 | (25–220) |
| RBC_EPA | RBC | 112 | (45–238) |
| Palmitic Acid (Acetate Adduct) | LCMS | 200 | (23–456) |


Appendix B
Appendix B.1. The Vitamin D and Omega-3 Trial (VITAL)
Appendix B.2. The Women’s Health Study (WHS)
Appendix B.3. Blood Sample Collection
Appendix B.4. Biomarker Profiling
Appendix B.4.1. Targeted Plasma Phospholipid Fatty Acids Measured with Plasma PL Assay
Appendix B.4.2. Fatty Acids Measured with RBC Assay
Appendix B.4.3. Non-targeted Metabolomic Measurements with LCMS Assay
Appendix B.4.4. Novel Oxylipins 1,2,3 Measured on LCMS Assay
References
- Hu, Y.; Hu, F.B.; Manson, J.E. Marine omega-3 supplementation and cardiovascular disease: An updated meta-analysis of 13 randomized controlled trials involving 127 477 participants. J. Am. Heart Assoc. 2019, 8, e013543. [Google Scholar] [CrossRef] [PubMed]
- Aung, T.; Halsey, J.; Kromhout, D.; Gerstein, H.C.; Marchioli, R.; Tavazzi, L.; Geleijnse, J.M.; Rauch, B.; Ness, A.; Galan, P. Associations of omega-3 fatty acid supplement use with cardiovascular disease risks: Meta-analysis of 10 trials involving 77 917 individuals. JAMA Cardiol. 2018, 3, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; Summerbell, C.D.; Worthington, H.V.; Song, F. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Bassuk, S.S.; Lee, I.M.; Cook, N.R.; Albert, M.A.; Gordon, D.; Zaharris, E.; Macfadyen, J.G.; Danielson, E.; Lin, J.; et al. The VITamin D and OmegA-3 TriaL (VITAL): Rationale and design of a large randomized controlled trial of vitamin D and marine omega-3 fatty acid supplements for the primary prevention of cancer and cardiovascular disease. Contemp. Clin. Trials 2012, 33, 159–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine n-3 Fatty Acids and Prevention of Cardiovascular Disease and Cancer. N. Engl. J. Med. 2019, 380, 23–32. [Google Scholar] [CrossRef]
- Bhatt, D.L.; Steg, P.G.; Miller, M.; Brinton, E.A.; Jacobson, T.A.; Ketchum, S.B.; Doyle, R.T., Jr.; Juliano, R.A.; Jiao, L.; Granowitz, C.; et al. Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia. N. Engl. J. Med. 2019, 380, 11–22. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr. 2000, 71, 343S–348S. [Google Scholar] [CrossRef]
- Gordon, W.C.; Bazan, N.G. Mediator lipidomics in ophthalmology: Targets for modulation in inflammation, neuroprotection and nerve regeneration. Curr. Eye Res. 2013, 38, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capra, V.; Bäck, M.; Barbieri, S.S.; Camera, M.; Tremoli, E.; Rovati, G.E. Eicosanoids and their drugs in cardiovascular diseases: Focus on atherosclerosis and stroke. Med. Res. Rev. 2013, 33, 364–438. [Google Scholar] [CrossRef]
- Watrous, J.D.; Niiranen, T.J.; Lagerborg, K.A.; Henglin, M.; Xu, Y.J.; Rong, J.; Sharma, S.; Vasan, R.S.; Larson, M.G.; Armando, A.; et al. Directed Non-targeted Mass Spectrometry and Chemical Networking for Discovery of Eicosanoids and Related Oxylipins. Cell Chem. Biol. 2019, 26, 433–442. [Google Scholar] [CrossRef]
- Cook, N.R.; Lee, I.M.; Gaziano, J.M.; Gordon, D.; Ridker, P.M.; Manson, J.E.; Hennekens, C.H.; Buring, J.E. Low-dose aspirin in the primary prevention of cancer: The Women’s Health Study: A randomized controlled trial. JAMA 2005, 294, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-M.; Cook, N.R.; Gaziano, J.M.; Gordon, D.; Ridker, P.M.; Manson, J.E.; Hennekens, C.H.; Buring, J.E. Vitamin E in the primary prevention of cardiovascular disease and cancer: The Women’s Health Study: A randomized controlled trial. JAMA 2005, 294, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-M.; Cook, N.R.; Manson, J.E.; Buring, J.E.; Hennekens, C.H. β-Carotene supplementation and incidence of cancer and cardiovascular disease: The Women’s Health Study. J. Natl. Cancer Inst. 1999, 91, 2102–2106. [Google Scholar] [CrossRef] [Green Version]
- Ridker, P.M.; Cook, N.R.; Lee, I.M.; Gordon, D.; Gaziano, J.M.; Manson, J.E.; Hennekens, C.H.; Buring, J.E. A randomized trial of low-dose aspirin in the primary prevention of cardiovascular disease in women. N. Engl. J. Med. 2005, 352, 1293–1304. [Google Scholar] [CrossRef]
- Henglin, M.; Niiranen, T.; Watrous, J.D.; Lagerborg, K.A.; Antonelli, J.; Claggett, B.L.; Demosthenes, E.J.; von Jeinsen, B.; Demler, O.; Vasan, R.S. A Single Visualization Technique for Displaying Multiple Metabolite–Phenotype Associations. Metabolites 2019, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Itakura, H.; Yokoyama, M.; Matsuzaki, M.; Saito, Y.; Origasa, H.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Kita, T. Relationships between plasma fatty acid composition and coronary artery disease. J. Atheroscler. Thromb. 2010, 18, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Superko, H.R.; Superko, S.M.; Nasir, K.; Agatston, A.; Garrett, B.C. Omega-3 fatty acid blood levels: Clinical significance and controversy. Circulation 2013, 128, 2154–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, T.; Yoshida, M.; Arita, M. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol. 2019, 31, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2–nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costenbader, K.H.; MacFarlane, L.A.; Lee, I.M.; Buring, J.E.; Mora, S.; Bubes, V.; Kotler, G.; Camargo, C.A., Jr.; Manson, J.E.; Cook, N.R. Effects of One Year of Vitamin D and Marine Omega-3 Fatty Acid Supplementation on Biomarkers of Systemic Inflammation in Older US Adults. Clin. Chem. 2019, 65, 1508–1521. [Google Scholar] [CrossRef]
- Benjamini, Y.; Kenigsberg, E.; Reiner, A.; Yekutieli, D. FDR Adjustments of Microarray Experiments (FDR-AME). Available online: http://bioconductor.jp/packages/3.0/bioc/vignettes/fdrame/inst/doc/fdrame.pdf (accessed on 23 October 2020).
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Yekutieli, D.; Benjamini, Y. Resampling-based false discovery rate controlling multiple test procedures for correlated test statistics. J. Stat. Plan. Inference 1999, 82, 171–196. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; Available online: https://www.R-project.org/ (accessed on 24 August 2020).
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D supplements and prevention of cancer and cardiovascular disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Harris, W.S.; Pottala, J.V.; Vasan, R.S.; Larson, M.G.; Robins, S.J. Changes in erythrocyte membrane trans and marine fatty acids between 1999 and 2006 in older Americans. J. Nutr. 2012, 142, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Pottala, J.V.; Espeland, M.A.; Polreis, J.; Robinson, J.; Harris, W.S. Correcting the effects of− 20 C storage and aliquot size on erythrocyte fatty acid content in the Women’s Health Initiative. Lipids 2012, 47, 835–846. [Google Scholar] [CrossRef]






| VITAL200 Substudy | ||||
|---|---|---|---|---|
| n-3 Treatment | Placebo | Total * | VITAL | |
| N | 100 | 100 | 200 | 25,871 |
| Age, y | 65 (60–71) | 64 (60–69) | 65 (60–70) | 67 (63–71) |
| African American, n | 50 (51%) | 49 (50%) | 99 (50%) | 5106 (20%) |
| Women, n | 53 (53%) | 51 (51%) | 104 (52%) | 13,085 (51%) |
| BMI ±, kg/m2 | 29 (25–3 3) | 28 (24–33) | 28 (25–3 3) | 27 (24–31) |
| HBPmed †, n | 63 (65%) | 59 (62%) | 122 (63%) | 13166 (51%) |
| Diabetes, n | 21 (21.2%) | 23 (23%) | 44 (22%) | 3686 (14.3%) |
| Current Smoker, n | 6 (6%) | 7 (7%) | 13 (7%) | 1836 (7%) |
| Statin Treatment, n | 35 (36%) | 40 (40%) | 75 (38%) | 8890 (35%) |
| Total Chol, mg/dL | 192 (169–210) | 187 (168–220) | 189 # (168–213) | 204 (179–232) |
| LDLC, mg/dL | 118 (100–1 36) | 112 (96–143) | 115 (96–138) | 124 (102–148) |
| HDLC, mg/dL | 47 (41–57) | 48 (37–56.2) | 47 (38–57) | 52 (42–65) |
| Trig, mg/dL | 108 (855–151) | 110 (81–141) | 109 (83–146) | 111 (83–155) |
| hsCRP, mg/L | 1.5 (0.8–4.0) | 1.7 (0.8–4.1) | 1.7 (0.8–4.1) | NA |
| IL6, pg/mL | 1.7 (1.0–2.2) | 1.7 (1.1–2.8) | 1.7 (1.1–2.5) | NA |
| TNRF2, ng/mL | 2651 (2104–3232) | 2459 (2051–3201) | 2530 (2068–3207) | NA |
| Substudy of the Women’s Health Study (Validation Cohort) | ||||
| Total | ||||
| N | 5129 | |||
| Age, y | 64 (61–68) | |||
| African American, n | 78 (2%) | |||
| Women, n | 5129 (100%) | |||
| BMI, kg/m2 | 25 (23–28) | |||
| HBPmed †, n | 1093 (21%) | |||
| Diabetes, n | 199 (4%) | |||
| Current Smoker, n | 492 (10%) | |||
| Statin Treatment, n | 314 (6%) | |||
| Total Chol, mg/dL | 220 (195–245) | |||
| LDLC, mg/dL | 130 (109–154) | |||
| HDLC, mg/dL | 52 (43–63) | |||
| Trig, mg/dL | 133 (94–190) | |||
| hsCRP, mg/L | 2.4 (1.1–4.8) | |||
| IL6, pg/mL | NA | |||
| TNRF2, ng/mL | NA | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demler, O.V.; Liu, Y.; Luttmann-Gibson, H.; Watrous, J.D.; Lagerborg, K.A.; Dashti, H.; Giulianini, F.; Heath, M.; Camargo, C.A., Jr.; Harris, W.S.; et al. One-Year Effects of Omega-3 Treatment on Fatty Acids, Oxylipins, and Related Bioactive Lipids and Their Associations with Clinical Lipid and Inflammatory Biomarkers: Findings from a Substudy of the Vitamin D and Omega-3 Trial (VITAL). Metabolites 2020, 10, 431. https://doi.org/10.3390/metabo10110431
Demler OV, Liu Y, Luttmann-Gibson H, Watrous JD, Lagerborg KA, Dashti H, Giulianini F, Heath M, Camargo CA Jr., Harris WS, et al. One-Year Effects of Omega-3 Treatment on Fatty Acids, Oxylipins, and Related Bioactive Lipids and Their Associations with Clinical Lipid and Inflammatory Biomarkers: Findings from a Substudy of the Vitamin D and Omega-3 Trial (VITAL). Metabolites. 2020; 10(11):431. https://doi.org/10.3390/metabo10110431
Chicago/Turabian StyleDemler, Olga V., Yanyan Liu, Heike Luttmann-Gibson, Jeramie D. Watrous, Kim A. Lagerborg, Hesam Dashti, Franco Giulianini, Mallory Heath, Carlos A. Camargo, Jr., William S. Harris, and et al. 2020. "One-Year Effects of Omega-3 Treatment on Fatty Acids, Oxylipins, and Related Bioactive Lipids and Their Associations with Clinical Lipid and Inflammatory Biomarkers: Findings from a Substudy of the Vitamin D and Omega-3 Trial (VITAL)" Metabolites 10, no. 11: 431. https://doi.org/10.3390/metabo10110431