Metabolites from Microbes Isolated from the Skin of the Panamanian Rocket Frog Colostethus panamansis (Anura: Dendrobatidae)

 ,
,  ,
,
Abstract
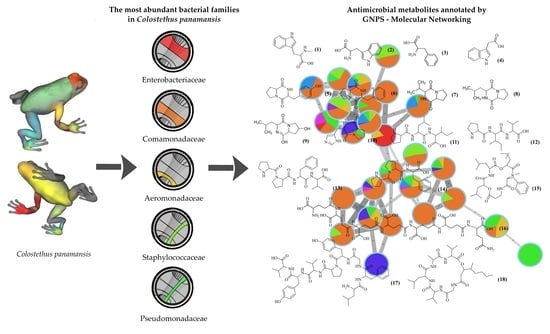
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. 16S rRNA Amplicon Sequencing and Molecular Cartography
2.2. Molecular Networking and Molecular Cartography
2.3. Comparison between 16S rRNA Sequences from C. panamansis and Bd-Inhibitory Public Datasets
3. Discussion
4. Materials and Methods
4.1. Sampling Specimens
4.2. 16S rRNA Amplicon Sequencing of Cultivable Bacteria
4.3. Fermentation and Crude Extracts Preparation
4.4. LC-MS/MS Analysis
4.5. MS/MS Data Analysis and Molecular Networking
4.6. 3D Modeling and Data Picturing
4.7. Bacterial Correlation for Inhibitory Properties Based on 16S rRNA Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feller, A.E.; Hedges, S.B. Molecular evidence for the early history of living amphibians. Mol. Phylogenet. Evol. 1998, 9, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Haddad, C.F.B.; Prado, C.P.A. Reproductive Modes in Frogs and Their Unexpected Diversity in the Atlantic Forest of Brazil. Bioscience 2005, 55, 207. [Google Scholar] [CrossRef] [Green Version]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lips, K.R.; Reeve, J.D.; Witters, L.R. Ecological Traits Predicting Amphibian Population Declines in Central America. Conserv. Biol. 2003, 17, 1078–1088. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A.; Carey, C.; Longcore, J.; Doersam, J.K.; Boutte, A.; Bruzgal, J.E.; Conlon, J.M. Activity of antimicrobial skin peptides from ranid frogs against Batrachochytrium dendrobatidis, the chytrid fungus associated with global amphibian declines. Dev. Comp. Immunol. 2002, 26, 471–479. [Google Scholar] [CrossRef]
- La Marca, E.; Lips, K.R.; Lotters, S.; Puschendorf, R.; Ibanez, R.; Rueda-Almonacid, J.V.; Schulte, R.; Marty, C.; Castro, F.; Manzanilla-Puppo, J.; et al. Catastrophic Population Declines and Extinctions in Neotropical Harlequin Frogs (Bufonidae: Atelopus)1. Biotropica 2005, 37, 190–201. [Google Scholar] [CrossRef]
- Mendelson, J.R., 3rd; Lips, K.R.; Gagliardo, R.W.; Rabb, G.B.; Collins, J.P.; Diffendorfer, J.E.; Daszak, P.; Ibáñez, D.R.; Zippel, K.C.; Lawson, D.P.; et al. Biodiversity. Confronting amphibian declines and extinctions. Science 2006, 313, 48. [Google Scholar] [CrossRef] [Green Version]
- Lips, K.R. Overview of chytrid emergence and impacts on amphibians. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [Green Version]
- Van Rooij, P.; Martel, A.; Haesebrouck, F.; Pasmans, F. Amphibian chytridiomycosis: A review with focus on fungus-host interactions. Vet. Res. 2015, 46, 137. [Google Scholar]
- Voyles, J.; Woodhams, D.C.; Saenz, V.; Byrne, A.Q.; Perez, R.; Rios-Sotelo, G.; Ryan, M.J.; Bletz, M.C.; Sobell, F.A.; McLetchie, S.; et al. Shifts in disease dynamics in a tropical amphibian assemblage are not due to pathogen attenuation. Science 2018, 359, 1517–1519. [Google Scholar]
- Grant, T.; Solís, F.; Ibáñez, R.; Jaramillo, C.; Fuenmayor, Q. IUCN Colostethus panamansis. In IUCN Red List of Threatened Species 2004; IUCN: Gland, Switzerland, 2004. [Google Scholar]
- Crawford, A.J.; Lips, K.R.; Bermingham, E. Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proc. Natl. Acad. Sci. USA 2010, 107, 13777–13782. [Google Scholar] [PubMed] [Green Version]
- Lips, K.R.; Brem, F.; Brenes, R.; Reeve, J.D.; Alford, R.A.; Voyles, J.; Carey, C.; Livo, L.; Pessier, A.P.; Collins, J.P. Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. Proc. Natl. Acad. Sci. USA 2006, 103, 3165–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, J.W.; Gusovsky, F.; Myers, C.W.; Yotsu-Yamashita, M.; Yasumoto, T. First occurrence of tetrodotoxin in a dendrobatid frog (Colostethus inguinalis), with further reports for the bufonid genus Atelopus. Toxicon 1994, 32, 279–285. [Google Scholar] [CrossRef]
- Wang, J.; Fan, Y.; Yao, Z. Isolation of a Lysinibacillus fusiformis strain with tetrodotoxin-producing ability from puffer fish Fugu obscurus and the characterization of this strain. Toxicon 2010, 56, 640–643. [Google Scholar] [CrossRef]
- Küng, D.; Bigler, L.; Davis, L.R.; Gratwicke, B.; Griffith, E.; Woodhams, D.C. Stability of microbiota facilitated by host immune regulation: Informing probiotic strategies to manage amphibian disease. PLoS ONE 2014, 9, e87101. [Google Scholar]
- Lauer, A.; Simon, M.A.; Banning, J.L.; Lam, B.A.; Harris, R.N. Diversity of cutaneous bacteria with antifungal activity isolated from female four-toed salamanders. ISME J. 2008, 2, 145–157. [Google Scholar] [CrossRef]
- Lauer, A.; Simon, M.A.; Banning, J.L.; André, E.; Duncan, K.; Harris, R.N. Common Cutaneous Bacteria from the Eastern Red-Backed Salamander Can Inhibit Pathogenic Fungi. Copeia 2007, 2007, 630–640. [Google Scholar] [CrossRef]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 2019, 363, 1459–1463. [Google Scholar]
- Brucker, R.M.; Baylor, C.M.; Walters, R.L.; Lauer, A.; Harris, R.N.; Minbiole, K.P.C. The identification of 2,4-diacetylphloroglucinol as an antifungal metabolite produced by cutaneous bacteria of the salamander Plethodon cinereus. J. Chem. Ecol. 2008, 34, 39–43. [Google Scholar] [CrossRef]
- Woodhams, D.C.; LaBumbard, B.C.; Barnhart, K.L.; Becker, M.H.; Bletz, M.C.; Escobar, L.A.; Flechas, S.V.; Forman, M.E.; Iannetta, A.A.; Joyce, M.D.; et al. Prodigiosin, Violacein, and Volatile Organic Compounds Produced by Widespread Cutaneous Bacteria of Amphibians Can Inhibit Two Batrachochytrium Fungal Pathogens. Microb. Ecol. 2018, 75, 1049–1062. [Google Scholar]
- Martin, H.C.; Ibáñez, R.; Nothias, L.-F.; Boya, P.C.A.; Reinert, L.K.; Rollins-Smith, L.A.; Dorrestein, P.C.; Gutiérrez, M. Viscosin-like lipopeptides from frog skin bacteria inhibit Aspergillus fumigatus and Batrachochytrium dendrobatidis detected by imaging mass spectrometry and molecular networking. Sci. Rep. 2019, 9, 3019. [Google Scholar]
- Woodhams, D.C.; Brandt, H.; Baumgartner, S.; Kielgast, J.; Küpfer, E.; Tobler, U.; Davis, L.R.; Schmidt, B.R.; Bel, C.; Hodel, S.; et al. Interacting symbionts and immunity in the amphibian skin mucosome predict disease risk and probiotic effectiveness. PLoS ONE 2014, 9, e96375. [Google Scholar]
- Da Silva, R.R.; Lopes, N.P.; Silva, D.B. CHAPTER 3: Metabolomics. In Mass Spectrometry in Chemical Biology; Royal Society of Chemistry: London, UK, 2017; pp. 57–81. [Google Scholar]
- Brunetti, A.E.; Lyra, M.L.; Melo, W.G.P.; Andrade, L.E.; Palacios-Rodríguez, P.; Prado, B.M.; Haddad, C.F.B.; Pupo, M.T.; Lopes, N.P. Symbiotic skin bacteria as a source for sex-specific scents in frogs. Proc. Natl. Acad. Sci. USA 2019, 116, 2124–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, N.; Kapono, C.; Lim, Y.W.; Koyama, N.; Vermeij, M.J.A.; Conrad, D.; Rohwer, F.; Dorrestein, P.C. Mass spectral similarity for untargeted metabolomics data analysis of complex mixtures. Int. J. Mass Spectrom. 2015, 377, 719–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar]
- Quinn, R.A.; Nothias, L.-F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular Networking As a Drug Discovery, Drug Metabolism, and Precision Medicine Strategy. Trends Pharmacol. Sci. 2017, 38, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based Molecular Networking in the GNPS Analysis Environment. bioRxiv 2019, 812404. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Bouslimani, A.; Porto, C.; Rath, C.M.; Wang, M.; Guo, Y.; Gonzalez, A.; Berg-Lyon, D.; Ackermann, G.; Moeller Christensen, G.J.; Nakatsuji, T.; et al. Molecular cartography of the human skin surface in 3D. Proc. Natl. Acad. Sci. USA 2015, 112, E2120–E2129. [Google Scholar] [CrossRef] [Green Version]
- Protsyuk, I.; Melnik, A.V.; Nothias, L.-F.; Rappez, L.; Phapale, P.; Aksenov, A.A.; Bouslimani, A.; Ryazanov, S.; Dorrestein, P.C.; Alexandrov, T. 3D molecular cartography using LC-MS facilitated by Optimus and ′ili software. Nat. Protoc. 2018, 13, 134–154. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.-F.; Nothias-Esposito, M.; da Silva, R.; Wang, M.; Protsyuk, I.; Zhang, Z.; Sarvepalli, A.; Leyssen, P.; Touboul, D.; Costa, J.; et al. Bioactivity-Based Molecular Networking for the Discovery of Drug Leads in Natural Product Bioassay-Guided Fractionation. J. Nat. Prod. 2018, 81, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Floros, D.J.; Petras, D.; Kapono, C.A.; Melnik, A.V.; Ling, T.-J.; Knight, R.; Dorrestein, P.C. Mass Spectrometry Based Molecular 3D-Cartography of Plant Metabolites. Front. Plant Sci. 2017, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Kapono, C.A.; Morton, J.T.; Bouslimani, A.; Melnik, A.V.; Orlinsky, K.; Knaan, T.L.; Garg, N.; Vázquez-Baeza, Y.; Protsyuk, I.; Janssen, S.; et al. Creating a 3D microbial and chemical snapshot of a human habitat. Sci. Rep. 2018, 8, 3669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, N.; Wang, M.; Hyde, E.; da Silva, R.R.; Melnik, A.V.; Protsyuk, I.; Bouslimani, A.; Lim, Y.W.; Wong, R.; Humphrey, G.; et al. Three-Dimensional Microbiome and Metabolome Cartography of a Diseased Human Lung. Cell Host Microbe 2017, 22, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Parrot, D.; Blümel, M.; Utermann, C.; Chianese, G.; Krause, S.; Kovalev, A.; Gorb, S.N.; Tasdemir, D. Mapping the Surface Microbiome and Metabolome of Brown Seaweed Fucus vesiculosus by Amplicon Sequencing, Integrated Metabolomics and Imaging Techniques. Sci. Rep. 2019, 9, 1061. [Google Scholar] [PubMed]
- Ross, A.A.; Hoffmann, A.R.; Neufeld, J.D. The skin microbiome of vertebrates. Microbiome 2019, 7, 79. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar]
- Woodhams, D.C.; Alford, R.A.; Antwis, R.E.; Archer, H.; Becker, M.H.; Belden, L.K.; Bell, S.C.; Bletz, M.; Daskin, J.H.; Davis, L.R.; et al. Antifungal isolates database of amphibian skin-associated bacteria and function against emerging fungal pathogens. Ecology 2015, 96, 595. [Google Scholar] [CrossRef] [Green Version]
- Elizabeth, A.; Grice, J.A.S. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244. [Google Scholar]
- Austin, R.M. Cutaneous Microbial Flora and Antibiosis in Plethodon Ventralis. In The Biology of Plethodontid Salamanders; Springer: Boston, MA, USA, 2000; pp. 451–462. [Google Scholar]
- Culp, C.E.; Falkinham, J.O.; Belden, L.K. Identification of the natural bacterial microflora on the skin of eastern newts, bullfrog tadpoles and redback salamanders. Herpetologica 2007, 63, 66–71. [Google Scholar] [CrossRef]
- Woodhams, D.C.; Vredenburg, V.T.; Simon, M.-A.; Billheimer, D.; Shakhtour, B.; Shyr, Y.; Briggs, C.J.; Rollins-Smith, L.A.; Harris, R.N. Symbiotic bacteria contribute to innate immune defenses of the threatened mountain yellow-legged frog, Rana muscosa. Biol. Conserv. 2007, 138, 390–398. [Google Scholar] [CrossRef]
- Rebollar, E.A.; Hughey, M.C.; Medina, D.; Harris, R.N.; Ibáñez, R.; Belden, L.K. Skin bacterial diversity of Panamanian frogs is associated with host susceptibility and presence of Batrachochytrium dendrobatidis. ISME J. 2016, 10, 1682–1695. [Google Scholar] [PubMed] [Green Version]
- Varela, B.J.; Lesbarrères, D.; Ibáñez, R.; Green, D.M. Environmental and Host Effects on Skin Bacterial Community Composition in Panamanian Frogs. Front. Microbiol. 2018, 9, 298. [Google Scholar] [PubMed]
- Grant, T. A new, toxic species of Colostethus (Anura: Dendrobatidae: Colostethinae) from the Cordillera Central of Colombia. Zootaxa 2007, 1555, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.H.; Walke, J.B.; Murrill, L.; Woodhams, D.C.; Reinert, L.K.; Rollins-Smith, L.A.; Burzynski, E.A.; Umile, T.P.; Minbiole, K.P.C.; Belden, L.K. Phylogenetic distribution of symbiotic bacteria from Panamanian amphibians that inhibit growth of the lethal fungal pathogen Batrachochytrium dendrobatidis. Mol. Ecol. 2015, 24, 1628–1641. [Google Scholar]
- Belden, L.K.; Hughey, M.C.; Rebollar, E.A.; Umile, T.P.; Loftus, S.C.; Burzynski, E.A.; Minbiole, K.P.C.; House, L.L.; Jensen, R.V.; Becker, M.H.; et al. Panamanian frog species host unique skin bacterial communities. Front. Microbiol. 2015, 6, 1171. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.H.; Richards-Zawacki, C.L.; Gratwicke, B.; Belden, L.K. The effect of captivity on the cutaneous bacterial community of the critically endangered Panamanian golden frog (Atelopus zeteki). Biol. Conserv. 2014, 176, 199–206. [Google Scholar]
- Belden, L.K.; Harris, R.N. Infectious diseases in wildlife: The community ecology context. Front. Ecol. Environ. 2007, 5, 533–539. [Google Scholar]
- Wolfender, J.L.; Allard, P.M. Integration of molecular networking and in-silico MS/MS fragmentation for sensitive high throughput natural products dereplication. Planta Med. 2016, 81, PCS7. [Google Scholar] [CrossRef]
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62, 5–16. [Google Scholar]
- Demori, I.; El Rashed, Z.; Corradino, V.; Catalano, A.; Rovegno, L.; Queirolo, L.; Salvidio, S.; Biggi, E.; Zanotti-Russo, M.; Canesi, L.; et al. Peptides for Skin Protection and Healing in Amphibians. Molecules 2019, 24, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirthgen, E.; Otten, W.; Tuchscherer, M.; Tuchscherer, A.; Domanska, G.; Brenmoehl, J.; Günther, J.; Ohde, D.; Weitschies, W.; Seidlitz, A.; et al. Effects of 1-Methyltryptophan on Immune Responses and the Kynurenine Pathway after Lipopolysaccharide Challenge in Pigs. Int. J. Mol. Sci. 2018, 19, 3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A.; Scott Fites, J.; Reinert, L.K.; Shiakolas, A.R.; Umile, T.P.; Minbiole, K.P.C. Immunomodulatory Metabolites Released by the Frog-Killing Fungus Batrachochytrium dendrobatidis. Infect. Immun. 2015, 83, 4565. [Google Scholar] [CrossRef] [Green Version]
- Katayama, N.; Makoto, K.; Kishida, O. An aquatic vertebrate can use amino acids from environmental water. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160996. [Google Scholar] [CrossRef]
- Walke, J.B.; Harris, R.N.; Reinert, L.K.; Rollins-Smith, L.A.; Woodhams, D.C. Social Immunity in Amphibians: Evidence for Vertical Transmission of Innate Defenses. Biotropica 2011, 43, 396–400. [Google Scholar]
- Bletz, M.C.; Loudon, A.H.; Becker, M.H.; Bell, S.C.; Woodhams, D.C.; Minbiole, K.P.C.; Harris, R.N. Mitigating amphibian chytridiomycosis with bioaugmentation: Characteristics of effective probiotics and strategies for their selection and use. Ecol. Lett. 2013, 16, 807–820. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.R.; Shilts, T.; Ertürk, U.; Timmer, L.W.; Ueng, P.P. Indole derivatives produced by the fungus Colletotrichum acutatum causing lime anthracnose and postbloom fruit drop of citrus. FEMS Microbiol. Lett. 2003, 226, 23–30. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, M.P.; Abraham, W.-R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, A.D. 2,5-Diketopiperazines: Synthesis, reactions, medicinal chemistry, and bioactive natural products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Luis, S.; Ballesteros, J.; Gutiérrez, M. Antibacterial constituents from the octocoral-associated bacterium Pseudoalteromonas sp. Rev. Latinoam. Quím 2011, 39, 75–83. [Google Scholar]
- Wang, W.-L.; Zhu, T.-J.; Tao, H.-W.; Lu, Z.-Y.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Three novel, structurally unique spirocyclic alkaloids from the halotolerant B-17 fungal strain of Aspergillus variecolor. Chem. Biodivers. 2007, 4, 2913–2919. [Google Scholar] [CrossRef]
- Wang, Y.; Mueller, U.G.; Clardy, J. Antifungal Diketopiperazines from Symbiotic Fungus of Fungus-Growing Ant Cyphomyrmex minutus. J. Chem. Ecol. 1999, 25, 935–941. [Google Scholar] [CrossRef]
- Raju, R.; Piggott, A.M.; Huang, X.-C.; Capon, R.J. Nocardioazines: A novel bridged diketopiperazine scaffold from a marine-derived bacterium inhibits P-glycoprotein. Org. Lett. 2011, 13, 2770–2773. [Google Scholar] [CrossRef]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.-H. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides. Molecules 2018, 23, 815. [Google Scholar]
- Flechas, S.V.; Acosta-González, A.; Escobar, L.A.; Kueneman, J.G.; Sánchez-Quitian, Z.A.; Parra-Giraldo, C.M.; Rollins-Smith, L.A.; Reinert, L.K.; Vredenburg, V.T.; Amézquita, A.; et al. Microbiota and skin defense peptides may facilitate coexistence of two sympatric Andean frog species with a lethal pathogen. ISME J. 2019, 13, 361–373. [Google Scholar]
- Hamano, K.; Kinoshita, M.; Furuya, K.; Miyamoto, M.; Takamatsu, Y.; Hemmi, A.; Tanzawa, K. Leualacin, a novel calcium blocker from Hapsidospora irregularis. I. Taxonomy, fermentation, isolation, physico-chemical and biological properties. J. Antibiot. 1992, 45, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Mohimani, H.; Gurevich, A.; Mikheenko, A.; Garg, N.; Nothias, L.-F.; Ninomiya, A.; Takada, K.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of peptidic natural products through database search of mass spectra. Nat. Chem. Biol. 2017, 13, 30–37. [Google Scholar] [PubMed] [Green Version]
- Ben-zur, N.; Goldman, D.M. γ-poly glutamic acid: A novel peptide for skin care. Cosmet. Toilet. 2007, 122, 65–74. [Google Scholar]
- Lee, N.-R.; Go, T.-H.; Lee, S.-M.; Jeong, S.-Y.; Park, G.-T.; Hong, C.-O.; Son, H.-J. In vitro evaluation of new functional properties of poly-γ-glutamic acid produced by Bacillus subtilis D7. Saudi J. Biol. Sci. 2014, 21, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenfeld, A.; Blond, A.; Gueye, S.; Herson, P.; Nay, B.; Dupont, J.; Prado, S. Insecticidal cyclodepsipeptides from Beauveria felina. J. Nat. Prod. 2011, 74, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Hossain, E.; Khanam, S.; Wu, C.; Lostracco-Johnson, S.; Thomas, D.; Katemauswa, M.; Gosmanov, C.; Li, D.; Woelfel-Monsivais, C.; Sankaranarayanan, K.; et al. 3D mapping of host-parasite-microbiome interactions reveals metabolic determinants of tissue tropism and disease tolerance in Chagas disease. bioRxiv 2019, 727917. [Google Scholar] [CrossRef]
- León, M.; Ferreira, C.R.; Eberlin, L.S.; Jarmusch, A.K.; Pirro, V.; Rodrigues, A.C.B.; Favaron, P.O.; Miglino, M.A.; Graham Cooks, R. Metabolites and Lipids Associated with Fetal Swine Anatomy via Desorption Electrospray Ionization—Mass Spectrometry Imaging. Sci. Rep. 2019, 9, 7247. [Google Scholar] [CrossRef]
- Berger, L.; Speare, R.; Skerratt, L.F. Distribution of Batrachochytrium dendrobatidis and pathology in the skin of green tree frogs Litoria caerulea with severe chytridiomycosis. Dis. Aquat. Organ. 2005, 68, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Puschendorf, R.; Bolaños, F. Detection of Batrachochytrium dendrobatidis in Eleutherodactylus fitzingeri: Effects of skin sample location and histologic stain. J. Wildl. Dis. 2006, 42, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Colombo, B.M.; Scalvenzi, T.; Benlamara, S.; Pollet, N. Microbiota and mucosal immunity in amphibians. Front. Immunol. 2015, 6, 111. [Google Scholar]
- Boyle, D.G.; Boyle, D.B.; Olsen, V.; Morgan, J.A.T.; Hyatt, A.D. Rapid quantitative detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using real-time Taqman PCR assay. Dis. Aquat. Organ. 2004, 60, 141–148. [Google Scholar] [CrossRef]
- Waddle, A.W.; Sai, M.; Levy, J.E.; Rezaei, G.; van Breukelen, F.; Jaeger, J.R. Systematic approach to isolating Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2018, 127, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Medina, D.; Walke, J.B.; Gajewski, Z.; Becker, M.H.; Swartwout, M.C.; Belden, L.K. Culture Media and Individual Hosts Affect the Recovery of Culturable Bacterial Diversity from Amphibian Skin. Front. Microbiol. 2017, 8, 1574. [Google Scholar] [PubMed] [Green Version]
- Bast, F.; Bast, F. Sequence similarity search, Multiple Sequence Alignment, Model Selection, Distance Matrix and Phylogeny Reconstruction. Protoc. Exch. 2013. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2006, 23, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching molecular structure databases with tandem mass spectra using CSI:FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin H., C.; Ibáñez, R.; Nothias, L.-F.; Caraballo-Rodríguez, A.M.; Dorrestein, P.C.; Gutiérrez, M. Metabolites from Microbes Isolated from the Skin of the Panamanian Rocket Frog Colostethus panamansis (Anura: Dendrobatidae). Metabolites 2020, 10, 406. https://doi.org/10.3390/metabo10100406
Martin H. C, Ibáñez R, Nothias L-F, Caraballo-Rodríguez AM, Dorrestein PC, Gutiérrez M. Metabolites from Microbes Isolated from the Skin of the Panamanian Rocket Frog Colostethus panamansis (Anura: Dendrobatidae). Metabolites. 2020; 10(10):406. https://doi.org/10.3390/metabo10100406
Chicago/Turabian StyleMartin H., Christian, Roberto Ibáñez, Louis-Félix Nothias, Andrés Mauricio Caraballo-Rodríguez, Pieter C. Dorrestein, and Marcelino Gutiérrez. 2020. "Metabolites from Microbes Isolated from the Skin of the Panamanian Rocket Frog Colostethus panamansis (Anura: Dendrobatidae)" Metabolites 10, no. 10: 406. https://doi.org/10.3390/metabo10100406