1. Introduction
Resource development and environmental pollution generated by industrial activities have been affected by global climate change. Climate change has influenced various environmental factors, such as temperature, humidity, precipitation, and evaporation, resulting in environmental issues, especially depletion of water resources [
1]. To overcome the depletion of water resources, one solution is to use agricultural water more efficiently. Water resources are most widely used in agriculture, but most of the water used in farming is wasted. There have been many research studies that forecast the state of water in plants by monitoring the humidity of the external surroundings [
2,
3,
4], but such studies involve a high level of uncertainty. Therefore, one needs to directly measure and analyze the various water-related conditions in plants [
5,
6].
Plants are largely constituted of water; it accounts for 80–90% of their entire weight. Water is essential for photosynthesis, nutrient absorption, transpiration, and temperature adjustment in plants [
7]. Therefore, droughts are a major abiotic stress factor that induces a significant change in the biological activity of plants. When plants are exposed to a drought condition, their related adaptive responses and biological responses over the short and long term can be observed. The stoma, found in the epidermis of leaves and stems and bordered by a pair of guard cells, exhibits the greatest change in the early stages of drought. CO
2 is absorbed and O
2 is released under normal conditions for photosynthesis with simultaneous transpiration. Under drought stress, plants suppress CO
2 absorption and water evaporation via the plant hormone abscisic acid (ABA), which leads to a decrease in turgor pressure and ultimately stoma closure [
8]. In short, a change in evaporation from a stoma is dependent on the external water conditions. Evaporation can translate the water status of a plant into an electrical signal when a sensor is attached to the backside of a leaf, where the stoma is located. The sensors should be lightweight, thin, and highly flexible, and they must remain electrically stable against changes in humidity.
Polyimide (PI) is applicable to be used as a substrate in the flexible plant drought sensor because of its outstanding properties. A PI film is formed by a spin-coating on the glass and annealing at 350 °C. By controlling the spin speed and time, PI can be very thin and light with high mechanical and chemical stability at the same time. In addition, PI can absorb water and exhibit the variation in relative permittivity (ε) with relative humidity (RH) [
9].
In this paper, we demonstrated a flexible PI-based plant drought sensor and its real-time monitoring system to obtain firsthand information from a plant and implement high efficiency in irrigation cultivation. A PI-based plant drought sensor could read humidity-dependent variations if it was attached well to the leaf and did not induce strain or tensile forces. The drought stress signal was measured as a form of variation in capacitance when stoma sizes were compared. Water stress responsive variation in capacitance was measured on a 24-h interval when plant watering was performed once a week. In addition, when integrated with the processing module and Bluetooth (BLE) [
10], the whole system showed great potential to be the next-generation plant monitoring sensor system.
3. Results and Discussion
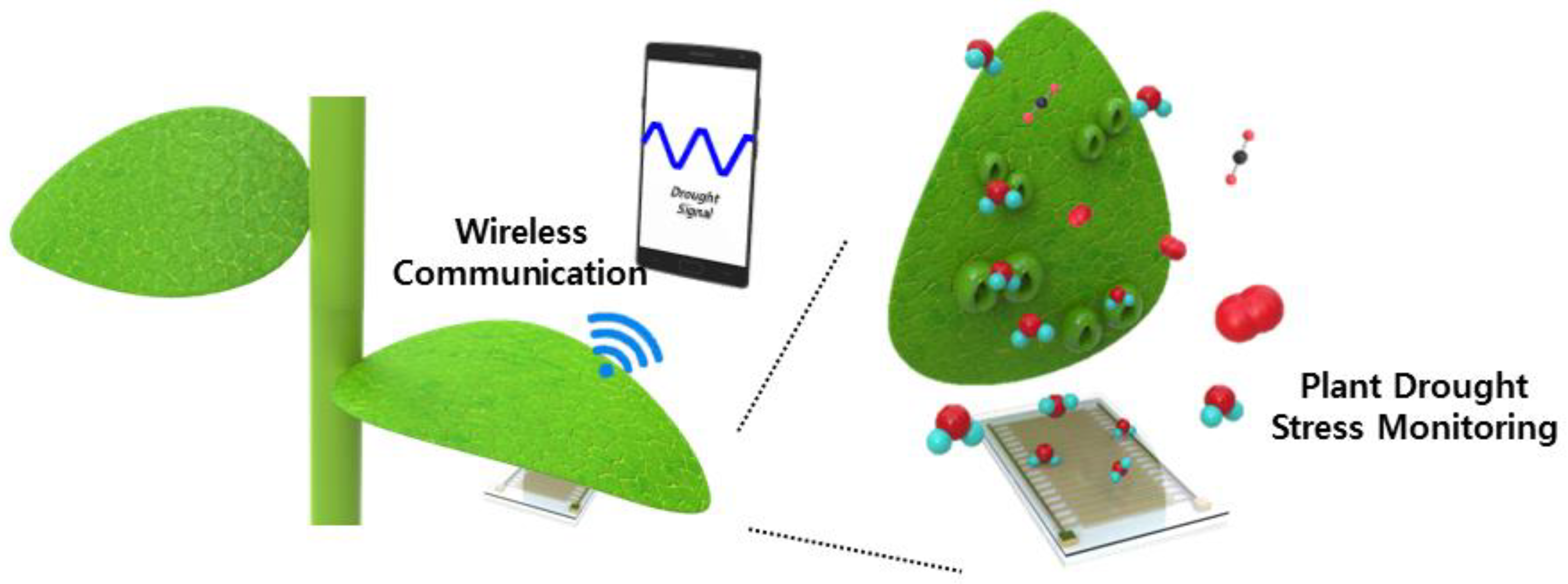
Figure 1 shows a scheme of the flexible PI-based plant drought stress sensor and its monitoring system integrated with wireless communication. The whole system is largely separated into two parts: a sensing part to measure differences and a processing part to send the processed signal to other mobile devices. For the sensing part, we focused on the stomata response to external drought stress. Stomata have evolved as a key organ of photosynthesis, nutrient uptake, cooling, in water status while controlling CO
2 uptake, and transpiration in variable environmental conditions. Stomata periodically regulate opening and closing linked to water efficiency in normal conditions. To minimize water loss and maximize photosynthesis gain, there is a typical transpiration tendency as responses to light-dependent and light-independent processes. When a water deficit occurs, the typical transpiration is no longer maintained. The stomata, which are supposed to be open, begin to close as a consequence of the short-term response to drought. As the stomata close, pore apertures formed between guard cells decrease. The decrease in pore apertures helps the plant to maintain water equilibrium for a longer period. In other words, pore aperture and transpiration from holes are in control of the water status of the plant. Though there are other responses in the leaf to drought stress including complicated and exquisite mechanisms, this paper covers the stomata response, by which the water status in a plant is translated into an electrical capacitance value that varies with respect to the water condition.
A capacitor on the flexible PI film was located on the lower (abaxial) surfaces and read the plant’s water stress as the capacitance signal. Due to the leaf curvature, the capacitor was designed to be flexible and light and should measure an accurate value when curved. The read-out circuit connected to the flexible sensor had wireless communication and was BLE integrated, so that it could effectively translate the time-varying data from the plant to the mobile devices as the plant drought stress monitoring system.
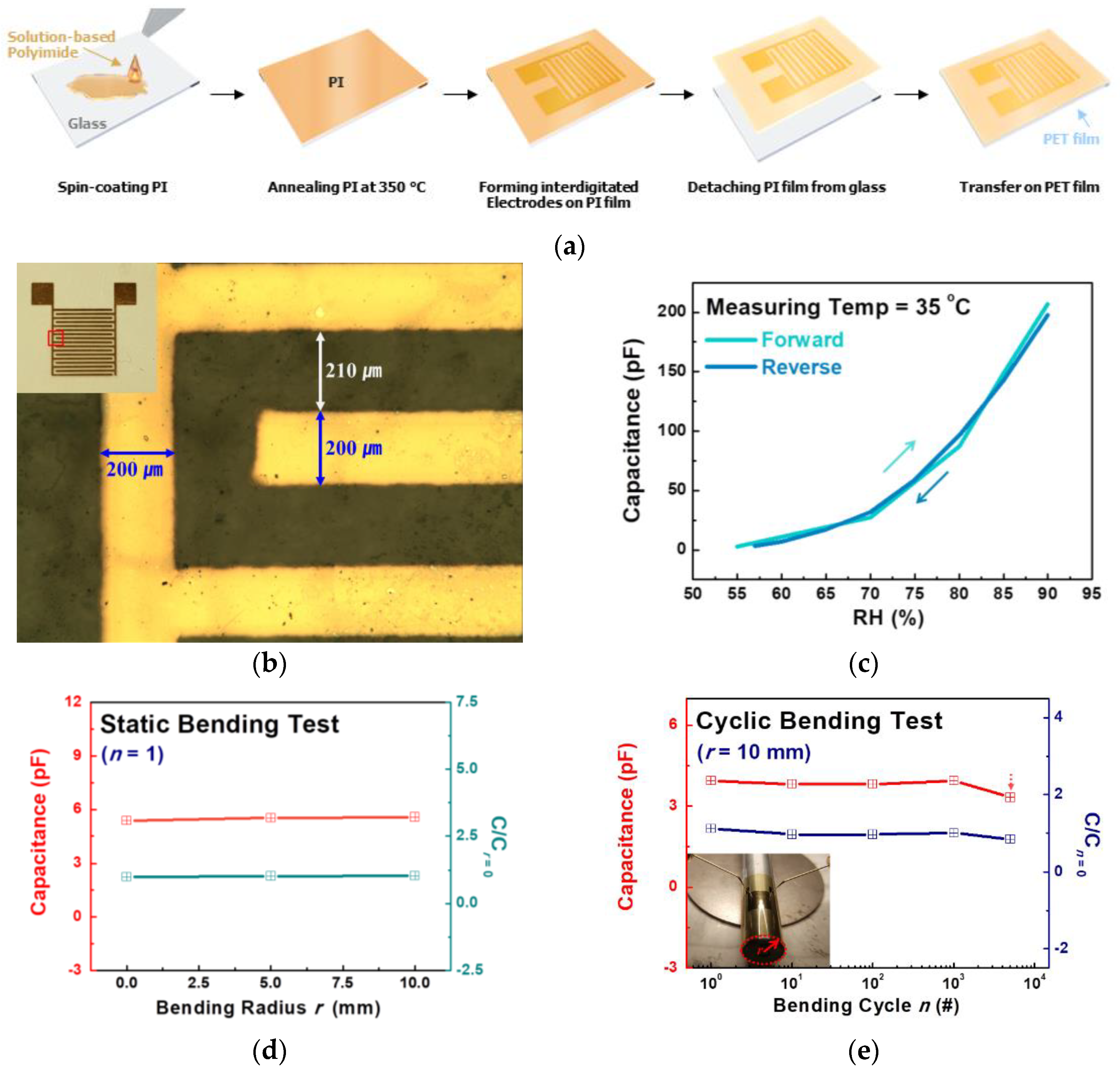
Figure 2a presents the fabrication process of the flexible PI-based plant drought sensor. The real image of the as-fabricated sensor and its optical image are shown in
Figure 2b and its inset. The electrode had fingers with a width of 200 μm and a length of 600 μm and a 210-μm spacing between fingers.
When photosynthesis occurs under normal conditions, a plant transpires through its stomata. Therefore, the RH under leaves reveals a status of transpiration. In general, the RH at the bottom of leaves is 50% and the RH enhances under the transpiration.
Figure 2c reveals the responses of the RH ranging from 50 to 90% at 35 °C in the flexible PI-based capacitive-type sensor. The RH response test was demonstrated by the thermos-hygrostat, while temperature and RH were adjusted at the same time to simulate the sensor behavior with variable RH prior to measuring the plant’s transpiration under drought stress. Forward and reverse mean the direction of pH variations. For example, the forward measure was to measure the change in the capacitance while the RH in the chamber was increased from 50%. In contrast, the reverse measure was obtained while the RH in the chamber was decreased from 90%. When the hydrophobic PI film was exposed to water vapor, the vapor diffused and was absorbed into the matrix, and the PI film underwent change in its relative permittivity (ε) with respect to surrounding RH [
9]. Pristine capacitance at 55% RH was 2.72 pF in the forward measure. According to the increase in RH, the capacitance of the sensor increased to 206 pF at 90% RH. The variation before and after the increase of the RH extended nearly up to 10
2 due to exponential growth in capacitance. Depending on the adjusted change direction of the RH, there was a slight hysteresis. However, the difference in cap at a certain RH was less than 5%, so we could ignore the hysteresis in the sensor.
In addition, leaves usually have certain curvatures, various radii (
r), and shake with the wind. Since leaves are held by thin and weak stalks (petioles), they are easily managed by applying strong mechanical stresses. Therefore, a lightweight and high flexibility are vital for a sensor attached to the backside of a leaf. Solution-based PI substrates have great flexibility [
11,
12], but the mechanical strength is low. To enhance the mechanical stress tolerance in the sensor, the PI film was laminated with a 100-μm-thick one-sided sticky PET film. As a consequence, harsh bending tests could be conducted for sensor stability tests.
Figure 2d shows the results of the static bending test. The capacitances of sensors were measured with flat conditions and bending conditions with bending radii of 5 mm and 10 mm at 1 MHz, respectively. The bending radii dependent variation in capacitance (C/C
0, where C and C
0 are the capacitance of the bent and unbent sensors, respectively) was negligible. In particular, the re-flattened sensor with 5-mm and 10-mm bending radii showed 1.030 and 1.037 differences, respectively, which were almost the same values as those of the pristine state. To confirm the stability of a sensor when a plant swayed in the wind, the cyclic bending test (
Figure 2e) with periodically repeated flat and upward bending was conducted at a 5-mm bending radius with bending cycles (
n) of 0, 10, 100, 1000, and 5000. As the number of cycles (
n) increased, the variation in capacitance slightly rose. When the sensor underwent bending cycles of 1, 10, 100, 1000, and 5000, the values of C/C
0 were 1.117, 0.968, 0.968, 1.001, and 0.8433, respectively. After the cyclic bending of 5000 cycles, degradations in capacitance and C/C
0 were generated. Like the static bending test, the change in capacitance was also negligible. Therefore, flexible PI-based sensors could be directly attached to the leaf as a node of the plant drought stress monitoring system.
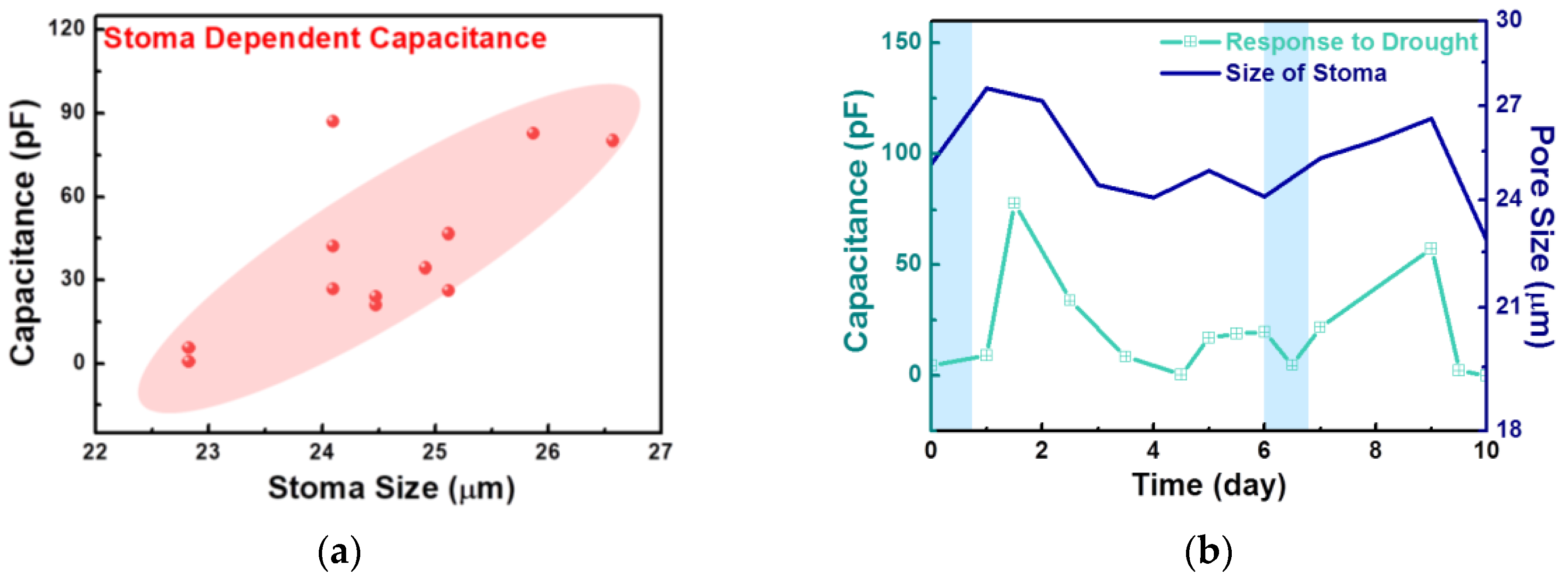
Figure 3a shows the relationship between stoma size and capacitance measured on the same plant. Capacitance obtained under various conditions is proportional to stoma size. There are many factors that influence transparency from the stoma and internal dielectrics in the leaves on certain conditions, which we did not discuss in this paper. A growth in stoma size induces an increase in the measured capacitance; therefore, we assumed that the water status in plants could be monitored as a form of electrical signal.
Figure 3b shows the time-varying capacitance and stoma size for 10 days. The opening and closure of the stoma are also significantly affected by the light and temperature, so the measurement time was fixed to illuminate variations from other factors. Watering occurred once every 6 days, as depicted with blue blocks in
Figure 3b. One day after watering, both the capacitance and stoma size increased, but as time passed, both decreased significantly, similar to in the pristine condition. As shown in
Figure 3c–e, stoma apertures differed, depending on the exposure time after watering. The stoma under proper water conditions had a larger pore, guard cells filled with water, and many ions due to osmotic pressure. The stoma under poor water conditions closed, and guard cells were shrunk to maintain homeostasis in the system. Even though stomata were located in the same leaf, there were differences in their morphologies. However, the measured data could be majorly attributed to the representative water status in the plant owing to the plant’s response to water conditions, showing a constant tendency.
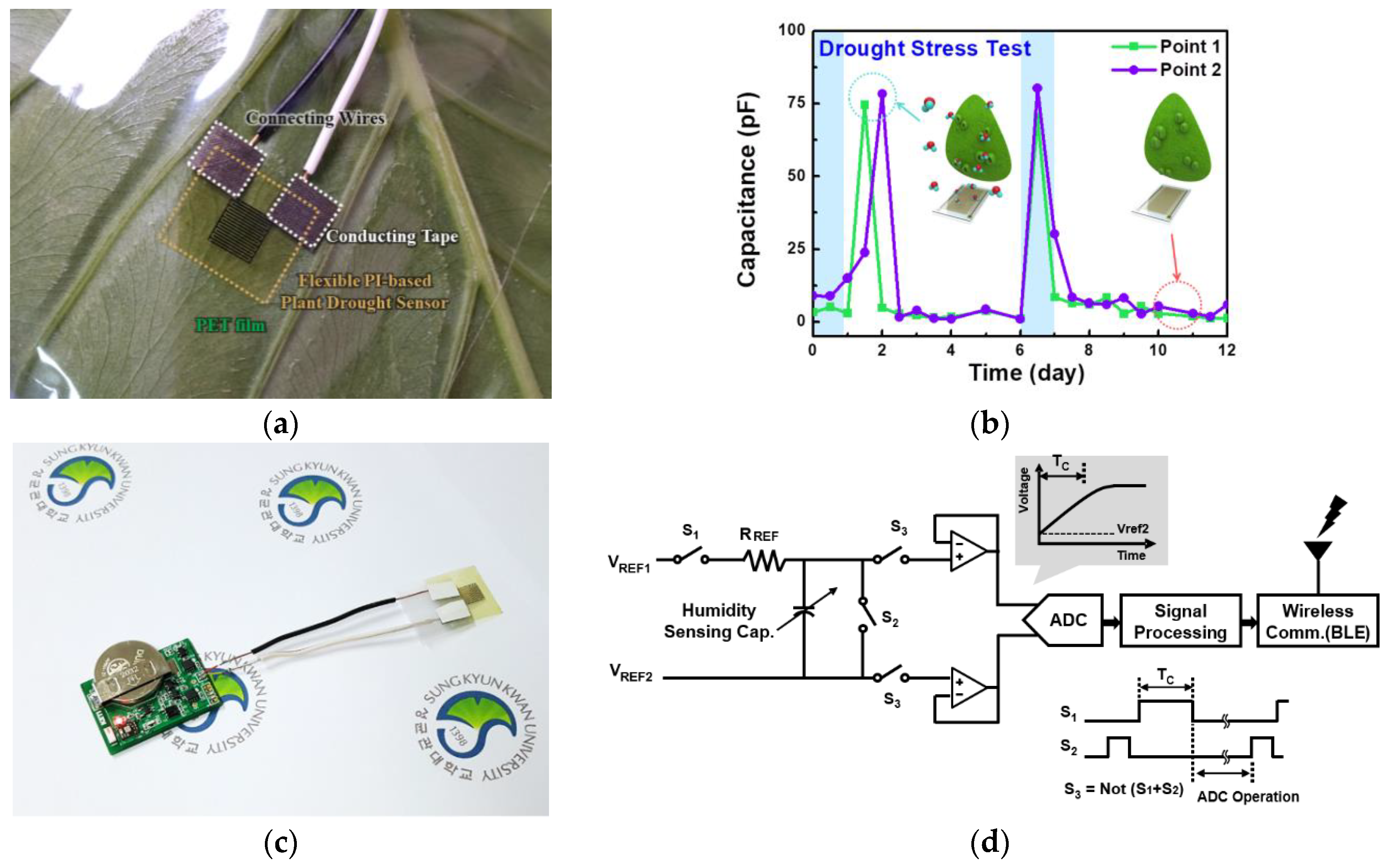
Figure 4a shows the real image of the flexible PI-based plant drought sensor attached on the lower surface of leaves. Using the one-side sticky PET film, the sensor was attached to leaves in a state. Its electrodes faced the leaves and connected with wires to measure the change in capacitance, depending on the water condition. The sensors were stuck on leaves without gap between them for accurate results.
Figure 4b presents the variation in capacitance measured in the same vein as responses to drought stress over time. Watering occurred once every 6 days, as depicted by the blue blocks. The measured capacitance after watering for 1 day and 7 days increased to nearly 75 pF, because the plant transpired and respired through the stomata properly. As shown in
Figure 3c and the inset in
Figure 3b, under normal conditions, most stomata in the leaf opened and transpired to activate photosynthesis and growth. Capacitance decreased and eventually saturated at a certain range, as shown in
Figure 4b, when the plant was exposed to drought. On the earlier day, some of the stomata in the leaf were still open and operated as a gas exchange path, as shown in
Figure 3d. Under serious drought conditions, most of the stomata in the leaf closed and stopped transpiration, under drought conditions, as shown in
Figure 3e.
Figure 4c is an image of the whole system for wireless and real-time flexible PI-based plant drought stress monitoring. The flexible PI-based plant drought sensor was integrated with a read-out processing circuit and a BLE module that could function in the real-time monitoring system for a smart farm to obtain firsthand information about plant response to water status.
The relation between humidity and the corresponding capacitance of the sensor was similar to a square function rather than a linear characteristic. To linearize the output signal of the sensor, a read-out circuit was designed with a simple linearized function. Because a simple resistance
capacitance (RC) time constant with a controlled timing signal showed an inverse exponential curve, a resistor and humidity sensor could be implemented to generate a linear voltage signal within a limited range.
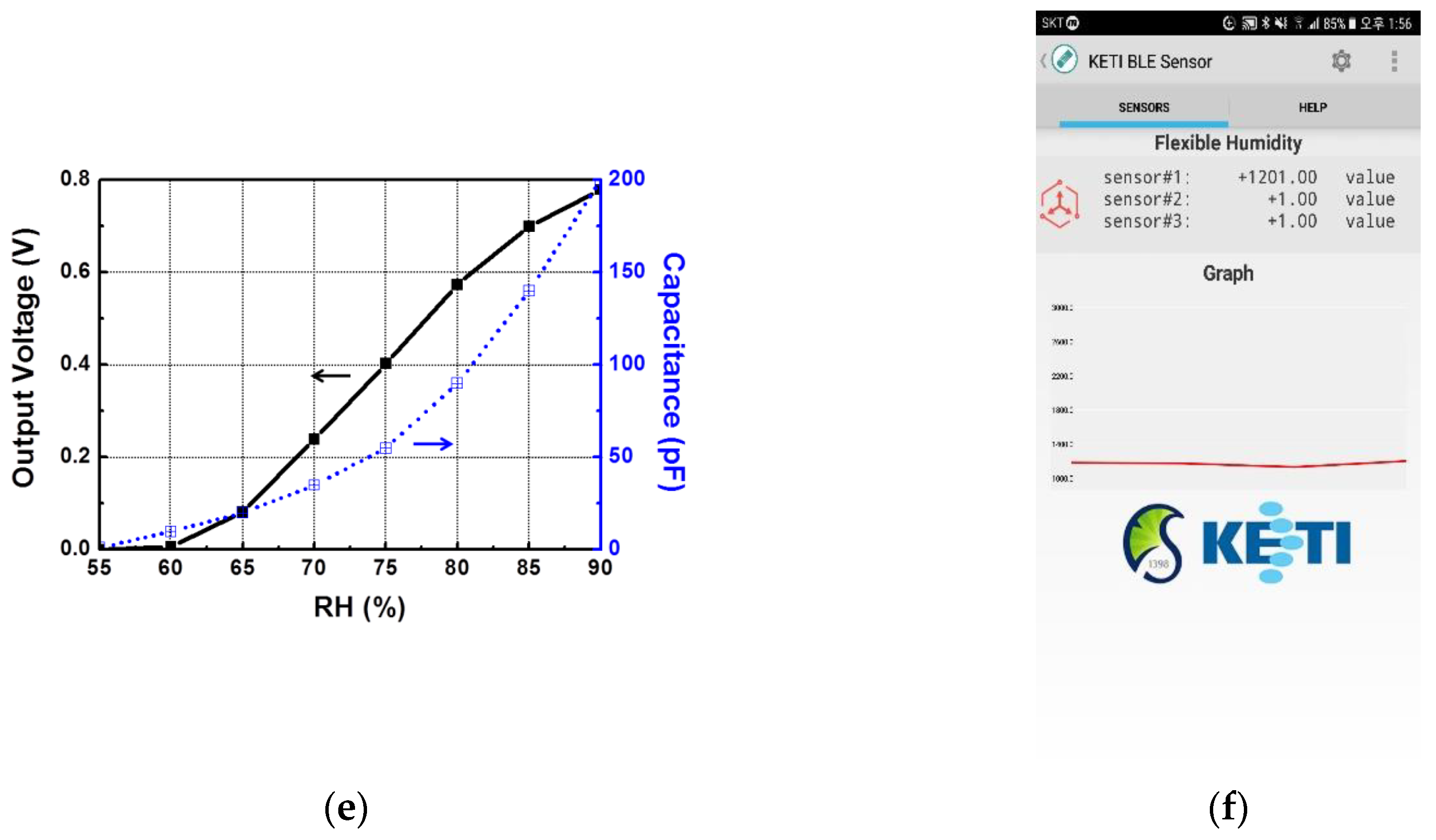
Figure 4d shows the designed read-out circuit with a resistor and a humidity sensor as an equivalent capacitor. Voltage buffers in front of an analog-to-digital convertor were employed to isolate an unwanted interaction. With a resistance (R
REF) of 1 Mohm and a time-slot (Tc) of 60 µs, the measured linear output voltage according to the RH is shown in
Figure 4e.
The measured data was analyzed at the processing circuit and wirelessly transmitted using the BLE module to a mobile device application, as shown in
Figure 4f. From these experimental results, we found great potential in the flexible plant drought sensor and its real-time monitoring system to be applied to a smart farm with advanced technologies, such as IoT, MEMS, and other wireless communications. The flexible PI-based real-time monitoring system will help gain further understanding about a plant’s physical mechanisms under various conditions and provide more direct information for highly efficient agricultural developments for water shortages.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





