Estrogenic Compounds or Adiponectin Inhibit Cyclic AMP Response to Human Luteinizing Hormone in Mouse Leydig Tumor Cells



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Hormones
2.2. Chemicals
2.3. Cell Culture
2.4. Plasmids, Transfections
2.5. cAMP Quantitation
2.6. Area Under Curve (AUC) Calculations and Statistical Analyses
3. Results
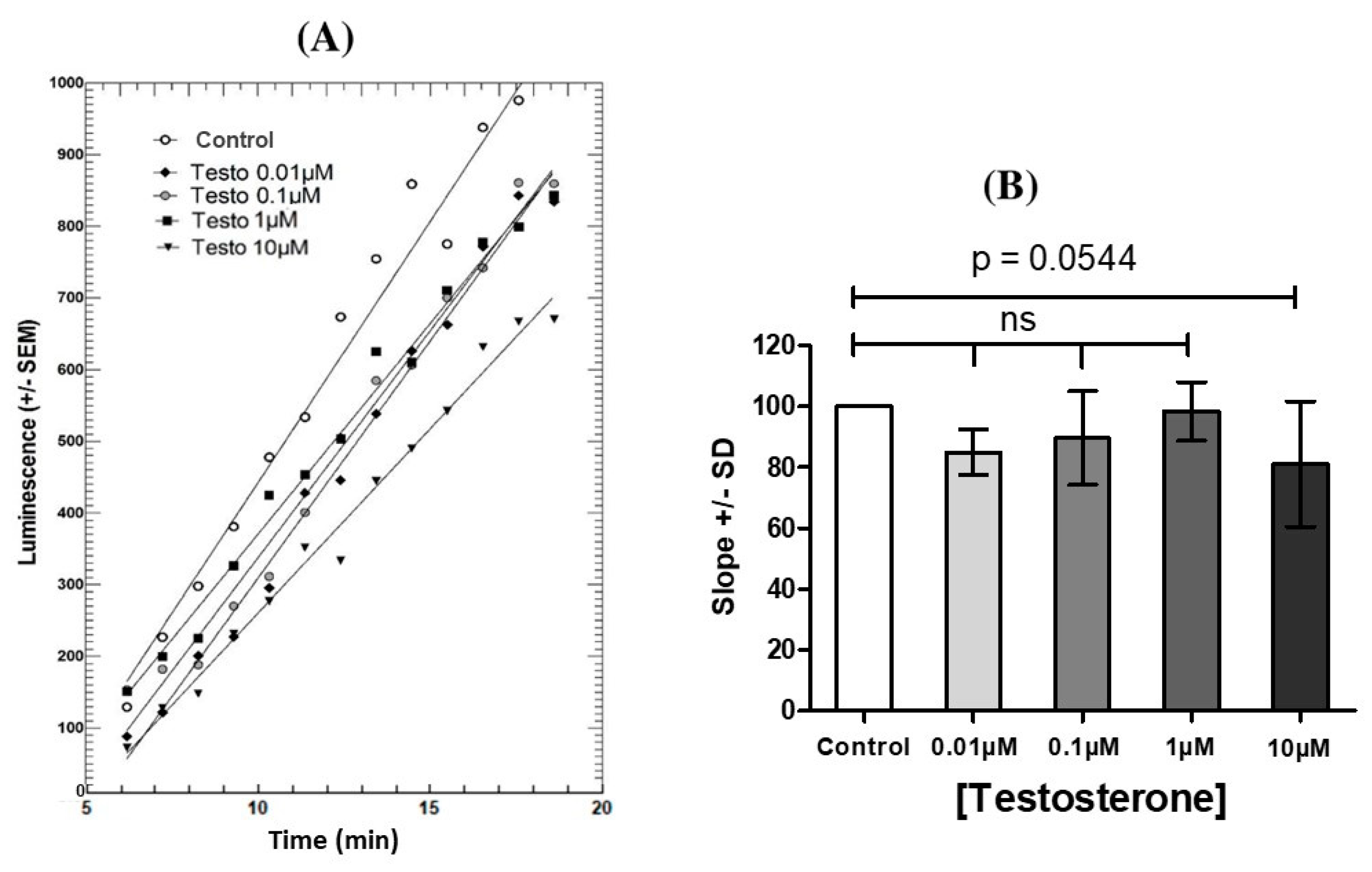
3.1. Effects of Steroid Hormones on LH-Stimulated cAMP Response
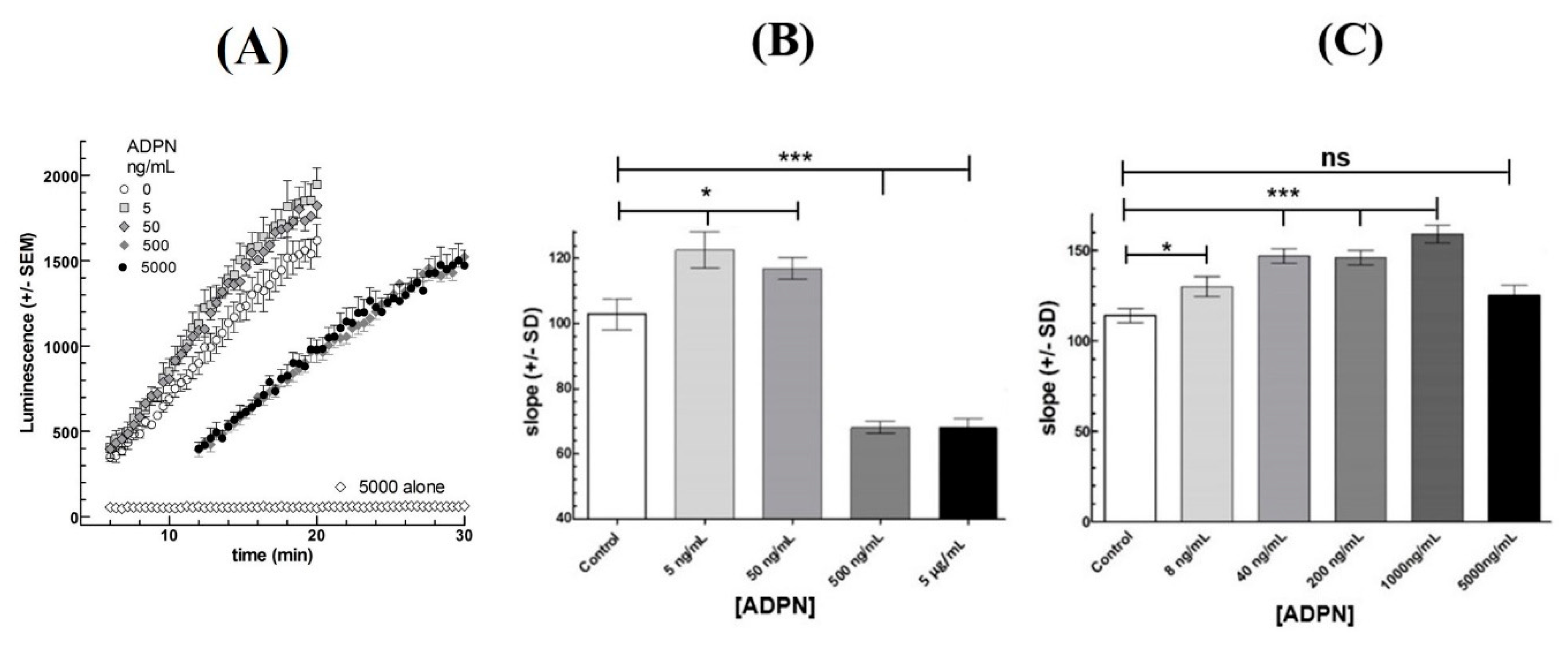
3.2. Effects of ADPN on LH-Stimulated cAMP Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: Formation, function, and regulation. Biol. Reprod. 2018, 99, 101–111. [Google Scholar] [CrossRef]
- Dimitriadis, F.; Tsiampali, C.; Chaliasos, N.; Tsounapi, P.; Takenaka, A.; Sofikitis, N. The Sertoli cell as the orchestra conductor of spermatogenesis: Spermatogenic cells dance to the tune of testosterone. Hormones 2015, 14, 479–503. [Google Scholar] [CrossRef]
- Rebois, R.V. Establishment of gonadotropin-responsive murine leydig tumor cell line. J. Cell Biol. 1982, 94, 70–76. [Google Scholar] [CrossRef]
- O’Hara, L.; McInnes, K.; Simitsidellis, I.; Morgan, S.; Atanassova, N.; Slowikowska-Hilczer, J.; Kula, K.; Szarras-Czapnik, M.; Milne, L.; Mitchell, R.T.; et al. Autocrine androgen action is essential for Leydig cell maturation and function, and protects against late-onset Leydig cell apoptosis in both mice and men. FASEB J. 2015, 29, 894–910. [Google Scholar] [CrossRef]
- Akingbemi, B.T.; Ge, R.; Rosenfeld, C.S.; Newton, L.G.; Hardy, D.O.; Catterall, J.F.; Lubahn, D.B.; Korach, K.S.; Hardy, M.P. Estrogen receptor-alpha gene deficiency enhances androgen biosynthesis in the mouse Leydig cell. Endocrinology 2003, 144, 84–93. [Google Scholar] [CrossRef]
- Ahima, R.S. Metabolic actions of adipocyte hormones: Focus on adiponectin. Obesity 2006, 14 (Suppl. 1), 9S–15S. [Google Scholar] [CrossRef]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef]
- Kos, K.; Harte, A.L.; da Silva, N.F.; Tonchev, A.; Chaldakov, G.; James, S.; Snead, D.R.; Hoggart, B.; O’Hare, J.P.; McTernan, P.G.; et al. Adiponectin and resistin in human cerebrospinal fluid and expression of adiponectin receptors in the human hypothalamus. J. Clin. Endocrinol. Metab. 2007, 92, 1129–1136. [Google Scholar] [CrossRef]
- Takemura, Y.; Osuga, Y.; Yamauchi, T.; Kobayashi, M.; Harada, M.; Hirata, T.; Morimoto, C.; Hirota, Y.; Yoshino, O.; Koga, K.; et al. Expression of adiponectin receptors and its possible implication in the human endometrium. Endocrinology 2006, 147, 3203–3210. [Google Scholar] [CrossRef]
- Kim, S.T.; Marquard, K.; Stephens, S.; Louden, E.; Allsworth, J.; Moley, K.H. Adiponectin and adiponectin receptors in the mouse preimplantation embryo and uterus. Hum. Reprod. 2011, 26, 82–95. [Google Scholar] [CrossRef]
- Smolinska, N.; Maleszka, A.; Dobrzyn, K.; Kiezun, M.; Szeszko, K.; Kaminski, T. Expression of adiponectin and adiponectin receptors 1 and 2 in the porcine uterus, conceptus, and trophoblast during early pregnancy. Theriogenology 2014, 82, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Sarnataro, D.; Mazzarella, G.; Sofia, M.; Bianco, A.; Daniele, A. Adiponectin affects lung epithelial A549 cell viability counteracting TNFalpha and IL-1ss toxicity through AdipoR1. Int. J. Biochem. Cell Biol. 2013, 45, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, I.; Kelesidis, T.; Mantzoros, C.S. Adiponectin and cancer: A systematic review. Br. J. Cancer 2006, 94, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.J.; Scalia, R. Adiponectin: A novel adipokine linking adipocytes and vascular function. J. Clin. Endocrinol. Metab. 2004, 89, 2563–2568. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Motoshima, H.; Mahadev, K.; Stalker, T.J.; Scalia, R.; Goldstein, B.J. Involvement of AMP-activated protein kinase in glucose uptake stimulated by the globular domain of adiponectin in primary rat adipocytes. Diabetes 2003, 52, 1355–1363. [Google Scholar] [CrossRef]
- Ye, R.; Scherer, P.E. Adiponectin, driver or passenger on the road to insulin sensitivity? Mol. Metab. 2013, 2, 133–141. [Google Scholar] [CrossRef]
- Combarnous, Y.; Guillou, F.; Martinat, N. Functional states of the luteinizing hormone/choriogonadotropin-receptor complex in rat Leydig cells. J. Biol. Chem. 1986, 261, 6868–6871. [Google Scholar]
- Gronemeyer, H. Transcription activation by estrogen and progesterone receptors. Annu. Rev. Genet. 1991, 25, 89–123. [Google Scholar] [CrossRef]
- Fujimoto, N.; Katzenellenbogen, B.S. Alteration in the agonist/antagonist balance of antiestrogens by activation of protein kinase A signaling pathways in breast cancer cells: Antiestrogen selectivity and promoter dependence. Mol. Endocrinol. 1994, 8, 296–304. [Google Scholar] [PubMed]
- Gruol, D.J.; Altschmied, J. Synergistic induction of apoptosis with glucocorticoids and 3’,5’-cyclic adenosine monophosphate reveals agonist activity by RU 486. Mol. Endocrinol. 1993, 7, 104–113. [Google Scholar] [PubMed]
- Beck, C.A.; Weigel, N.L.; Moyer, M.L.; Nordeen, S.K.; Edwards, D.P. The progesterone antagonist RU486 acquires agonist activity upon stimulation of cAMP signaling pathways. Proc. Natl. Acad. Sci. USA 1993, 90, 4441–4445. [Google Scholar] [CrossRef] [PubMed]
- Somers, J.P.; DeFranco, D.B. Effects of okadaic acid, a protein phosphatase inhibitor, on glucocorticoid receptor-mediated enhancement. Mol. Endocrinol. 1992, 6, 26–34. [Google Scholar] [PubMed]
- Ali, S.; Metzger, D.; Bornert, J.M.; Chambon, P. Modulation of transcriptional activation by ligand-dependent phosphorylation of the human oestrogen receptor A/B region. EMBO J. 1993, 12, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, P.; Montano, M.M.; Schodin, D.J.; Katzenellenbogen, B.S. Phosphorylation of the human estrogen receptor. Identification of hormone-regulated sites and examination of their influence on transcriptional activity. J. Biol. Chem. 1994, 269, 4458–4466. [Google Scholar] [PubMed]
- Catt, K.J.; Dufau, M.L.; Tsuruhara, T. Studies on a radioligand-receptor assay system for luteinizing hormone and chorionic gonadotropin. J. Clin. Endocrinol. Metab. 1971, 32, 860–863. [Google Scholar] [CrossRef]
- Catt, K.J.; Dufau, M.L.; Tsuruhara, T. Radioligand-receptor assay of luteinizing hormone and chorionic gonadotropin. J. Clin. Endocrinol. Metab. 1972, 34, 123–132. [Google Scholar] [CrossRef]
- Klett, D.; Meslin, P.; Relav, L.; Nguyen, T.M.; Mariot, J.; Jegot, G.; Cahoreau, C.; Combarnous, Y. Low reversibility of intracellular cAMP accumulation in mouse Leydig tumor cells (MLTC-1) stimulated by human Luteinizing Hormone (hLH) and Chorionic Gonadotropin (hCG). Mol. Cell. Endocrinol. 2016, 434, 144–153. [Google Scholar] [CrossRef]
- Kotula-Balak, M.; Pawlicki, P.; Milon, A.; Tworzydlo, W.; Sekula, M.; Pacwa, A.; Gorowska-Wojtowicz, E.; Bilinska, B.; Pawlicka, B.; Wiater, J.; et al. The role of G-protein-coupled membrane estrogen receptor in mouse Leydig cell function-in vivo and in vitro evaluation. Cell Tissue Res. 2018, 374, 389–412. [Google Scholar] [CrossRef]
- Milon, A.; Pawlicki, P.; Rak, A.; Mlyczynska, E.; Płachno, B.J.; Tworzydlo, W.; Gorowska-Wojtowicz, E.; Bilinska, B.; Kotula-Balak, M. Telocytes are localized to testis of the bank vole (Myodes glareolus) and are affected by lighting conditions and G-coupled membrane estrogen receptor (GPER) signaling. Gen. Comp. Endocrinol. 2019, 271, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Yasar, P.; Ayaz, G.; User, S.D.; Gupur, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Thomas, P. Minireview: G Protein-Coupled Estrogen Receptor-1, GPER-1: Its Mechanism of Action and Role in Female Reproductive Cancer, Renal and Vascular Physiology. Endocrinology 2012, 153, 2953–2962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Guan, R.; Segaloff, D.L. Revisiting and questioning functional rescue between dimerized LH receptor mutants. Mol. Endocrinol. 2012, 26, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, M.; Guan, R.; Segaloff, D.L. Heterodimerization between the lutropin and follitropin receptors is associated with an attenuation of hormone-dependent signaling. Endocrinology 2013, 154, 3925–3930. [Google Scholar] [CrossRef] [PubMed]
- Scaling, A.L.; Prossnitz, E.R.; Hathaway, H.J. GPER mediates estrogen-induced signaling and proliferation in human breast epithelial cells and normal and malignant breast. Horm. Cancer 2014, 5, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, A.C.; Peyton, C.; Dong, J.; Thomas, P. Bisphenol A and Related Alkylphenols Exert Nongenomic Estrogenic Actions Through a G Protein-Coupled Estrogen Receptor 1 (Gper)/Epidermal Growth Factor Receptor (Egfr) Pathway to Inhibit Meiotic Maturation of Zebrafish Oocytes. Biol. Reprod. 2015, 93, 135. [Google Scholar] [CrossRef]
- Acconcia, F.; Pallottini, V.; Marino, M. Molecular Mechanisms of Action of BPA. Dose Response 2015, 13. [Google Scholar] [CrossRef]
- Song, S.; Duan, Y.; Zhang, T.; Zhang, B.; Zhao, Z.; Bai, X.; Xie, L.; He, Y.; Ouyang, J.-P.; Huang, X.; et al. Serum concentrations of bisphenol A and its alternatives in elderly population living around e-waste recycling facilities in China: Associations with fasting blood glucose. Ecotoxicol. Environ. Saf. 2019, 169, 822–828. [Google Scholar] [CrossRef]
- Caminos, J.E.; Nogueiras, R.; Gaytan, F.; Pineda, R.; Gonzalez, C.R.; Barreiro, M.L.; Castano, J.P.; Malagon, M.M.; Pinilla, L.; Toppari, J.; et al. Novel expression and direct effects of adiponectin in the rat testis. Endocrinology 2008, 149, 3390–3402. [Google Scholar] [CrossRef]
- Wu, L.; Xu, B.; Fan, W.; Zhu, X.; Wang, G.; Zhang, A. Adiponectin protects Leydig cells against proinflammatory cytokines by suppressing the nuclear factor-kappaB signaling pathway. FEBS J. 2013, 280, 3920–3927. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.; Pare, A.; Jean, S.; Martin, L.J. Adiponectin influences progesterone production from MA-10 Leydig cells in a dose-dependent manner. Endocrine 2015, 48, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Ocon-Grove, O.M.; Krzysik-Walker, S.M.; Maddineni, S.R.; Hendricks, G.L., 3rd; Ramachandran, R. Adiponectin and its receptors are expressed in the chicken testis: Influence of sexual maturation on testicular ADIPOR1 and ADIPOR2 mRNA abundance. Reproduction 2008, 136, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Hoffstedt, J.; Arvidsson, E.; Sjolin, E.; Wahlen, K.; Arner, P. Adipose tissue adiponectin production and adiponectin serum concentration in human obesity and insulin resistance. J. Clin. Endocrinol. Metab. 2004, 89, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Orio, F., Jr.; Palomba, S.; Cascella, T.; Milan, G.; Mioni, R.; Pagano, C.; Zullo, F.; Colao, A.; Lombardi, G.; Vettor, R. Adiponectin levels in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2003, 88, 2619–2623. [Google Scholar] [CrossRef] [PubMed]
- Tsao, T.S. Assembly of adiponectin oligomers. Rev. Endocr. Metab. Disord. 2014, 15, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.B.; Giron, R.M.; Malinowski, P.R.; Nunez, M.; Tsao, T.S. Role of redox environment on the oligomerization of higher molecular weight adiponectin. BMC Biochem. 2011, 12, 24. [Google Scholar] [CrossRef]
- Horakova, D.; Stepanek, L.; Nagelova, R.; Pastucha, D.; Azeem, K.; Kollarova, H. Total and high-molecular-weight adiponectin levels and prediction of insulin resistance. Endokrynol. Pol. 2018, 69, 375–380. [Google Scholar]
- Hu, J.; Cui, W.; Ding, W.; Gu, Y.; Wang, Z.; Fan, W. Globular Adiponectin Attenuated H2O2-Induced Apoptosis in Rat Chondrocytes by Inducing Autophagy Through the AMPK/ mTOR Pathway. Cell. Physiol. Biochem. 2017, 43, 367–382. [Google Scholar] [CrossRef]
- Wang, S.; Miao, J.; Qu, M.; Yang, G.Y.; Shen, L. Adiponectin modulates the function of endothelial progenitor cells via AMPK/eNOS signaling pathway. Biochem. Biophys. Res. Commun. 2017, 493, 64–70. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Y.; Liu, L.; Lin, C.; Liao, C.; Xin, L.; Zhong, S.; Cheng, Q.; Zhang, L. Adiponectin Inhibits TNF-alpha-Activated PAI-1 Expression Via the cAMP-PKA-AMPK-NF-kappaB Axis in Human Umbilical Vein Endothelial Cells. Cell. Physiol. Biochem. 2017, 42, 2342–2352. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gurke, J.; Fischer, B.; Navarrete Santos, A. Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Zhang, L.; Gao, P.; Wu, X. Adiponectin attenuates high glucose-induced apoptosis through the AMPK/p38 MAPK signaling pathway in NRK-52E cells. PLoS ONE 2017, 12, e0178215. [Google Scholar]
- Abdou, H.S.; Bergeron, F.; Tremblay, J.J. A cell-autonomous molecular cascade initiated by AMP-activated protein kinase represses steroidogenesis. Mol. Cell. Biol. 2014, 34, 4257–4271. [Google Scholar] [CrossRef] [PubMed]
- Gorowska-Wojtowicz, E.; Dutka, P.; Kudrycka, M.; Pawlicki, P.; Milon, A.; Plachno, B.J.; Tworzydlo, W.; Pardyak, L.; Kaminska, A.; Hejmej, A.; et al. Regulation of steroidogenic function of mouse Leydig cells: G-coupled membrane estrogen receptor and peroxisome proliferator-activated receptor partnership. J. Physiol. Pharmacol. 2018, 69, 373–390. [Google Scholar]
- Zheng, F.; Zhang, S.; Lu, W.; Wu, F.; Yin, X.; Yu, D.; Pan, Q.; Li, H. Regulation of insulin resistance and adiponectin signaling in adipose tissue by liver X receptor activation highlights a cross-talk with PPARgamma. PLoS ONE 2014, 9, e101269. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.M.D.; Klett, D.; Combarnous, Y. Estrogenic Compounds or Adiponectin Inhibit Cyclic AMP Response to Human Luteinizing Hormone in Mouse Leydig Tumor Cells. Biology 2019, 8, 45. https://doi.org/10.3390/biology8020045
Nguyen TMD, Klett D, Combarnous Y. Estrogenic Compounds or Adiponectin Inhibit Cyclic AMP Response to Human Luteinizing Hormone in Mouse Leydig Tumor Cells. Biology. 2019; 8(2):45. https://doi.org/10.3390/biology8020045
Chicago/Turabian StyleNguyen, Thi Mong Diep, Danièle Klett, and Yves Combarnous. 2019. "Estrogenic Compounds or Adiponectin Inhibit Cyclic AMP Response to Human Luteinizing Hormone in Mouse Leydig Tumor Cells" Biology 8, no. 2: 45. https://doi.org/10.3390/biology8020045
APA StyleNguyen, T. M. D., Klett, D., & Combarnous, Y. (2019). Estrogenic Compounds or Adiponectin Inhibit Cyclic AMP Response to Human Luteinizing Hormone in Mouse Leydig Tumor Cells. Biology, 8(2), 45. https://doi.org/10.3390/biology8020045