
Endocrine and Local IGF-I in the Bony Fish Immune System

{kind=link}
Abstract
:1. Introduction: The GH/IGF-System and the Immune System
2. GH and IGF-I Actions in Bony Fish Immune Responses
- pro-opio-melanocortin hormones;
- thyroid system hormones;
- GH and prolactin;
- melatonin, histamine, serotonin and catecholamines;
- gonadotropin-releasing hormones; and
- their respective receptors.
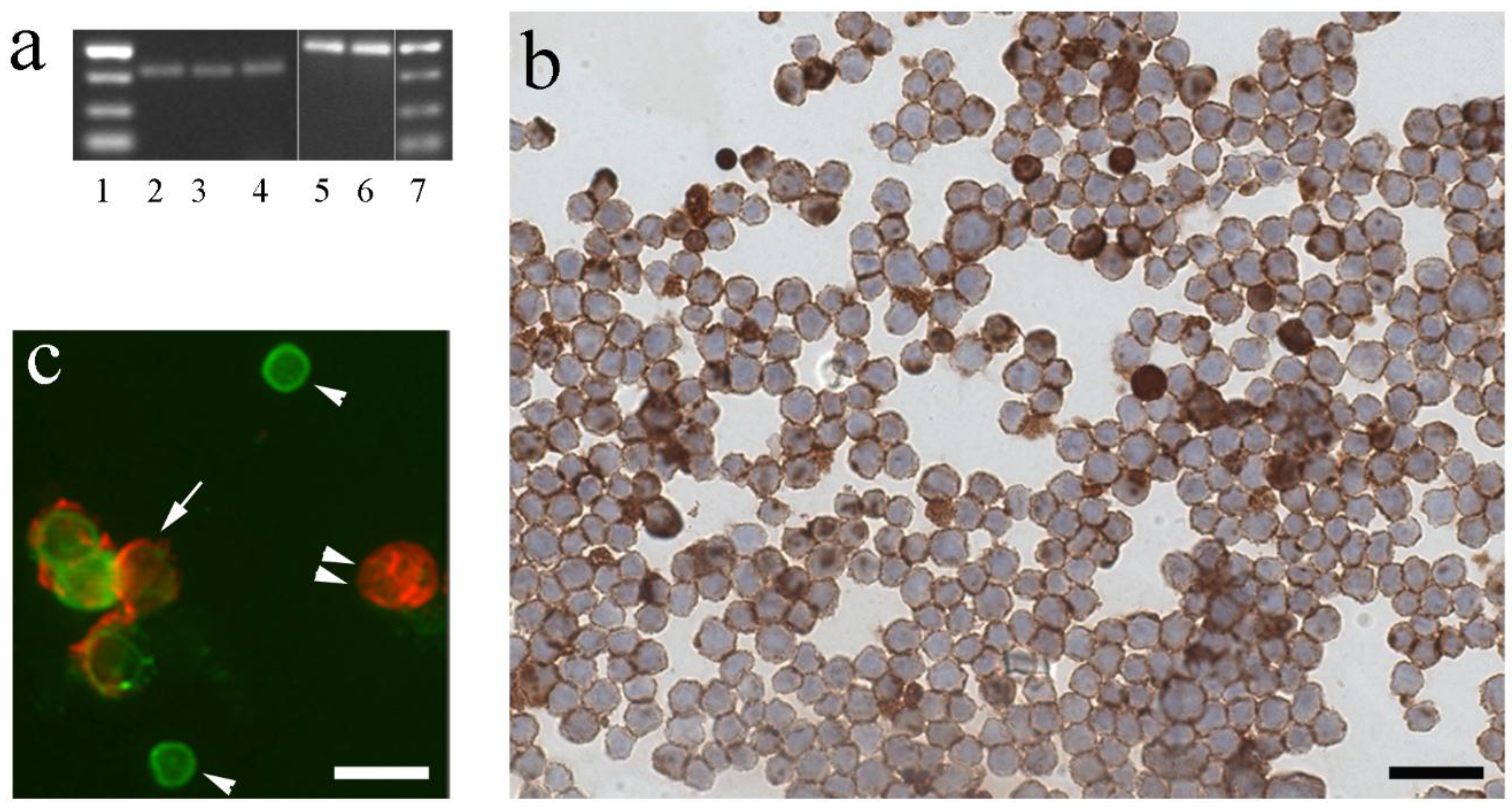
3. Immune Cells of Fish as Site of Local IGF Synthesis
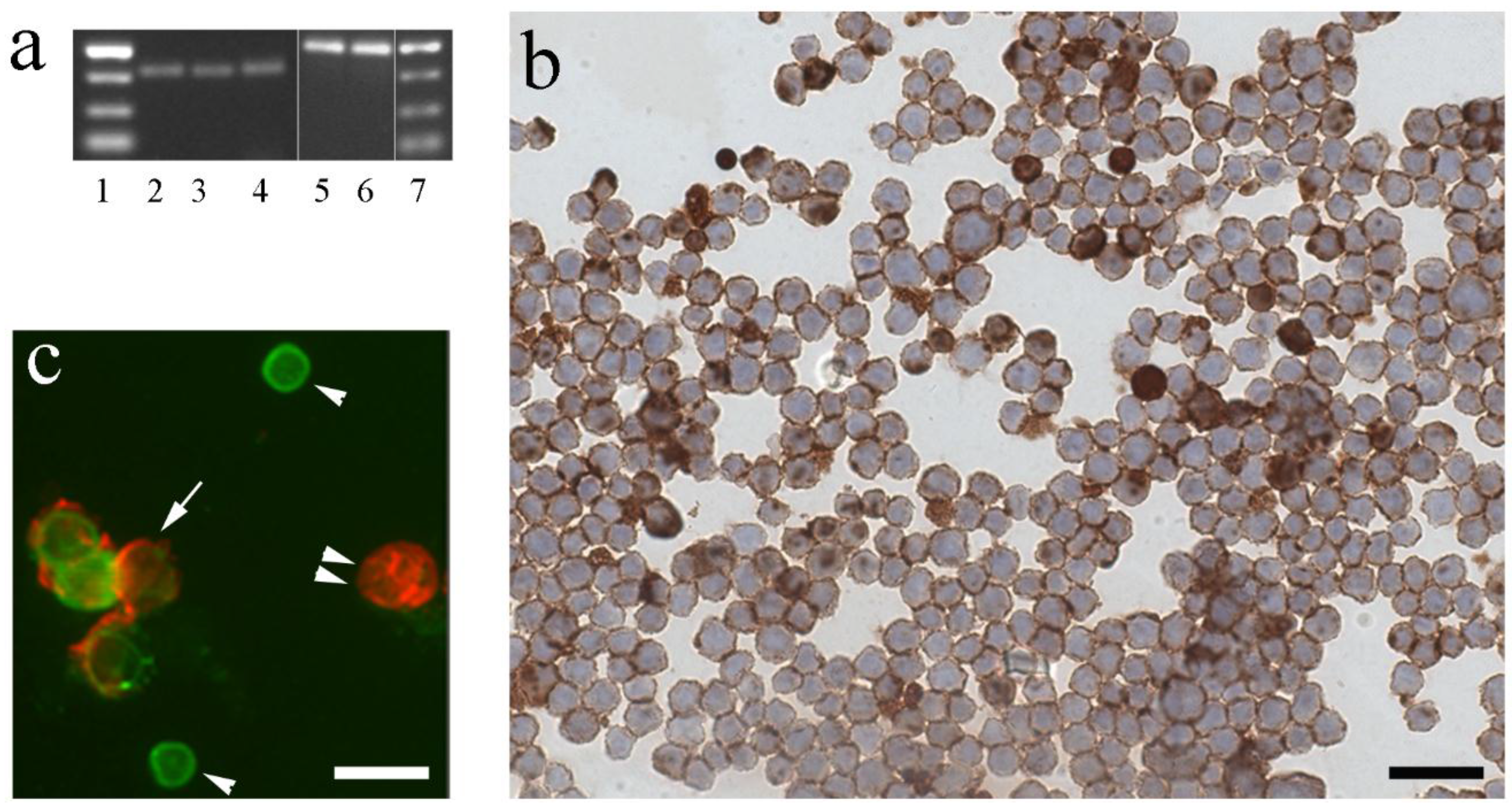
4. Regulation of IGF-I in Fish Immune Organs and Cells by GH and Other Physiological Factors
5. IGF-I and TNF-α during Pathogenic Infection of Fish
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Clark, R. The somatogenic hormones and insulin-like growth factor-1: Stimulators of lymphopoiesis and immune function. Endocr. Rev. 1997, 18, 157–179. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N. Expression, regulation and biological actions of growth hormone (GH) and ghrelin in the immune system. Growth Horm. IGF Res. 2009, 19, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J. Insulin-like growth factor-I regulation of immune function: A potential therapeutic target in autoimmune diseases? Pharmacol. Rev. 2010, 62, 199–236. [Google Scholar] [CrossRef] [PubMed]
- Weigent, D.A. Lymphocyte GH-axis hormones in immunity. Review. Cell Immunol. 2013, 285, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Segner, H.; Eppler, E.; Reinecke, M. The impact of environmental hormonally active substances on the endocrine and immune system of fish. Review. In Fish Endocrinology; Reinecke, M., Zaccone, G., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2006; Volume 2, pp. 809–865. [Google Scholar]
- Yada, T. Growth hormone and fish immune system. Gen. Comp. Endocrinol. 2007, 152, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [PubMed]
- Wallenius, K.; Sjögren, K.; Peng, X.D.; Park, S.; Wallenius, V.; Liu, J.L.; Umaerus, M.; Wennbo, H.; Isaksson, O.; Frohman, L.; et al. Liver-derived IGF-I regulates GH secretion at the pituitary level in mice. Endocrinology 2001, 142, 4762–4770. [Google Scholar] [CrossRef] [PubMed]
- Honda, J.; Manabe, Y.; Matsumura, R.; Takeuchi, S.; Takahashi, S. IGF-I regulates pro-opiomelanocortin and GH gene expression in the mouse pituitary gland. J. Endocrinol. 2003, 178, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Duan, C. Nutritional and developmental regulation of insulin-like growth factors in fish. J. Nutr. 1998, 128, 306S–314S. [Google Scholar] [PubMed]
- Reinecke, M.; Björnsson, B.T.; Dickhoff, W.W.; McCormick, S.D.; Navarro, I.; Power, D.M.; Gutiérrez, J. Growth hormone and insulin-like growth factors in fish: Where we are and where to go. Gen. Comp. Endocrinol. 2005, 142, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.W.; Duan, C.; Bern, H.A. Insulin-like growth factor signaling in fish. Int. Rev. Cytol. 2005, 243, 215–285. [Google Scholar] [PubMed]
- Reinecke, M. Insulin and the insulin-like growth factors. In Fish Endocrinology; Reinecke, M., Zaccone, G., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2006; Volume 1, pp. 3–13. [Google Scholar]
- Reinecke, M. Influences of the environment on the endocrine and paracrine fish growth hormone-insulin-like growth factor-I system. J. Fish Biol. 2010, 76, 1233–1254. [Google Scholar] [CrossRef] [PubMed]
- Reinecke, M.; Collet, C. The phylogeny of the insulin-like growth factors. Int. Rev. Cytol. 1998, 183, 1–94. [Google Scholar] [PubMed]
- Reinecke, M. Insulin-like growth factors and fish reproduction. Biol. Reprod. 2010, 82, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; Jiao, B.; Hu, C.; Huang, X.; Liu, Z.; Cheng, C.H. Discovery of a gonad-specific IGF subtype in teleost. Biochem. Biophys. Res. Comm. 2008, 367, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Berishvili, G.; Baroiller, J.-F.; Eppler, E.; Reinecke, M. Insulin-like growth factor-3 (IGF-3) in male and female gonads of the tilapia: Development and regulation of gene expression by growth hormone (GH) and 17alpha-ethinylestradiol (EE2). Gen. Comp. Endocrinol. 2010, 167, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Reindl, K.M.; Sheridan, M.A. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp. Biochem. Physiol. A 2012, 163, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Yada, T.; Azuma, T.; Takagi, Y. Stimulation of non-specific immune functions in seawater-acclimated rainbow trout, Oncorhynchus mykiss, with reference to the role of growth hormone. Comp. Biochem. Physiol. B 2001, 129, 695–701. [Google Scholar] [CrossRef]
- Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Alvarez-Pellitero, P.; Pérez-Sánchez, J. Evidence for a direct action of GH on haemopoietic cells of a marine fish, the gilthead sea bream (Sparus aurata). J. Endocrinol. 1995, 146, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Kajita, Y.; Sakai, M.; Kobayashi, M.; Kawauchi, H. Enhancement of non-specific cytotoxic activity of leucocytes in rainbow trout Oncorhynchus mykiss injected with growth hormone. Fish Shellfish Immunol. 1992, 2, 155–157. [Google Scholar] [CrossRef]
- Sakai, M.; Kobayashi, M.; Kawauchi, H. Enhancement of chemoluminiscent responses of phagocytic cells from rainbow trout, Oncorhynchus mykiss, by injection of growth hormone. Fish Shellfish Immunol. 1995, 5, 375–379. [Google Scholar] [CrossRef]
- Sakai, M.; Kobayashi, M.; Kawauchi, H. In vitro activation of fish phagocytic cells by GH, prolactin and somatolactin. J. Endocrinol. 1996, 151, 113–158. [Google Scholar] [CrossRef] [PubMed]
- Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Alvarez-Pellitero, P.; Pérez-Sánchez, J. Growth hormone is a phagocyte-activating factor in the gilthead sea bream (Sparus aurata). Cell Tissue Res. 1997, 287, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Kajita, Y.; Kobayashi, M.; Kawauchi, H. Immunostimulating effect of growth hormone: In vivo administration of growth hormone in rainbow trout enhances resistance to Vibrio anguillarum infection. Vet. Immunol. Immunopathol. 1997, 57, 147–152. [Google Scholar] [CrossRef]
- Kitlen, J.W.; Hejbol, E.K.; Zinck, T.; Varming, K.; Byatt, J.C.; McLean, E. Growth performance and respiratory burst activity in rainbow trout treated with growth hormone and vaccine. Fish Shellfish Immunol. 1997, 7, 297–304. [Google Scholar] [CrossRef]
- Peterson, B.C.; Small, B.C.; Bilodeau, L. Effects of GH on immune and endocrine responses of channel catfish challenged with Edwardsiella ictaluri. Comp. Biochem. Physiol. A 2007, 146, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Yada, T. Effects of insulin-like growth factor-I on non-specific immune functions in rainbow trout. Zool. Sci. 2009, 26, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, A.L.; Peterson, B.C.; Bosworth, B.G. Response of toll-like receptors, lysozyme, and IGF-I in back-cross hybrid (F1 male (blue × channel) × female channel) catfish challenged with virulent Edwardsiella ictaluri. Fish Shellfish Immunol. 2006, 20, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Berishvili, G.; Shved, N.; Eppler, E.; Clota, F.; Baroiller, J.F.; Reinecke, M. Organspecific expression of IGF-1 during early development of bony fish as revealed in the tilapia, Oreochromis niloticus, by in situ hybridization and immunohistochemistry: Indication for the particular importance of local IGF-I. Cell Tissue Res. 2006, 325, 287–301. [Google Scholar] [CrossRef] [PubMed]
- De Groof, F.; Joosten, K.F.; Janssen, J.A.; de Kleijn, E.D.; Hazelzet, J.A.; Hop, W.C.; Uitterlinden, P.; van Doorn, J.; Hokken-Koelega, A.C. Acute stress response in children with meningococcal sepsis: Important differences in the growth hormone/insulin-like growth factor I axis between nonsurvivors and survivors. J. Clin. Endocrinol. Metab. 2002, 87, 3118–3124. [Google Scholar] [CrossRef] [PubMed]
- Onenli-Mungan, N.; Yildizdas, D.; Yapicioglu, H.; Topaloglu, A.K.; Yüksel, B.; Ozer, G. Growth hormone and insulin-like growth factor 1 levels and their relation to survival in children with bacterial sepsis and septic shock. J. Paediatr. Child Health 2004, 40, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Mohan, S.; Sjögren, K.; Tivesten, A.; Isgaard, J.; Isaksson, O.; Jansson, J.O.; Svensson, J. The role of liver-derived insulin-like growth factor-I. Endocr. Rev. 2009, 30, 494–535. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, L.A.; Schmidt, D.; Gotway, M.B.; Ameli, N.; Filbert, E.L.; Ng, M.M.; Clor, J.L.; Epling, L.; Sinclair, E.; Baum, P.D.; et al. Growth hormone enhances thymic function in HIV-1-infected adults. J. Clin. Investig. 2008, 118, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G. Hormones in the immune system and their possible role. A critical review. Acta Microbiol. Immunol. Hung. 2014, 61, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Körbel, S.; Bittorf, T.; Siegl, E.; Köllner, B. Recombinant human erythropoietin induces proliferation and Ca(2+)-influx in specific leukocyte subpopulations of rainbow trout (Oncorhynchus mykiss) blood and head kidney cells. J. Comp. Physiol. B 2004, 174, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Verburg-van Kemenade, L.; Stolte, E.H.; Metz, J.R.; Chadzinska, M. Neuroendocrine-immune interactions in teleost fish. In Fish Neuroendocrinology, Fish Physiology; Bernier, N.J., van der Kraak, G., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Burlington, MA, USA, 2009; Volume 28, pp. 313–364. [Google Scholar]
- Eppler, E. The insulin-like growth factor I (IGF-I) within the bony fish pituitary: New morphofunctional and phylogenetic aspects. Open Neuroendocrinol. J. 2011, 4, 43–50. [Google Scholar] [CrossRef]
- Reinecke, M.; Schmid, A.; Ermatinger, R.; Loffing-Cueni, D. Insulin-like growth factor I in the teleost Oreochromis mossambicus, the tilapia: Gene sequence, tissue expression, and cellular localization. Endocrinology 1997, 138, 3613–3619. [Google Scholar] [CrossRef] [PubMed]
- Sitjà-Bobadilla, A.; Calduch-Giner, J.; Saera-Vila, A.; Palenzuela, O.; Álvarez-Pellitero, P.; Pérez-Sanchez, J. Chronic exposure to the parasite Enteromyxum leei (Myxozoa: Myxosporea) modulates the immune response and the expression of growth, redox and immune relevant genes in gilthead sea bream, Sparus aurata L. Fish Shellfish Immunol. 2008, 24, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Shved, N.; Berishvili, G.; Häusermann, E.; D’Cotta, H.; Baroiller, J.F.; Eppler, E. Challenge with EE2 during early development persistently impairs growth, differentiation and local expression of IGF-1 and-2 in immune organs of tilapia. Fish Shellfish Immunol. 2009, 26, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Shved, N.; Berishvili, G.; Mazel, P.; Baroiller, J.F.; Eppler, E. Growth hormone treatment acts on the endocrine and autocrine/paracrine GH/IGF-axis and on TNF-α expression in bony fish pituitary and immune organs. Fish Shellfish Immunol. 2011, 31, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Wenger, M.; Shved, N.; Akgül, G.; Caelers, A.; Casanova, A.; Segner, H.; Eppler, E. Developmental oestrogen exposure differentially modulates IGF-I and TNF-α expression levels in immune organs of Yersinia ruckeri-challenged young adult rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2014, 205, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Eppler, E.; Berishvili, G.; Mazel, P.; Caelers, A.; Hwang, G.; Maclean, N.; Reinecke, M. Distinct organ-specific up- and downregulation of IGF-I and IGF-II mRNA in various organs of a GH-overexpressing transgenic Nile tilapia. Transgenic Res. 2010, 19, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Lynn, S.G.; Shepherd, B.S. Molecular characterization and sex-specific tissue expression of prolactin, somatolactin and insulin-like growth factor-I in yellow perch (Perca flavescens). Comp. Biochem. Physiol. B 2007, 147, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Caelers, A.; Berishvili, G.; Meli, M.L.; Eppler, E.; Reinecke, M. Establishment of a real-time RT-PCR for the determination of absolute amounts of IGF-1 and IGF-2 gene expression in liver and extrahepatic sites of the tilapia. Gen. Comp. Endocrinol. 2004, 137, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Ponce, M.; Infante, C.; Funes, V.; Manchado, M. Molecular characterization and gene expression analysis of insulin-like growth factors I and II in the redbanded seabream, Pagrus auriga: Transcriptional regulation by growth hormone. Comp. Biochem. Physiol. B 2008, 150, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Hogan, R.J.; Waldbieser, G.; Goudie, C.A.; Antao, A.; Godwin, U.B.; Wilson, M.R.; Miller, N.W.; Clem, L.W.; McConnell, T.J.; Wolters, W.R.; et al. Molecular and immunological characterization of gynogenetic channel catfish (Ictalurus punctatus). Mar. Biotechnol. 1999, 1, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Secombes, C.J.; Long, S.; Miller, N.; Clem, L.W.; Chinchar, V.G. Molecular identification and expression analysis of tumor necrosis factor in channel catfish (Ictalurus punctatus). Dev. Comp. Immunol. 2003, 27, 845–858. [Google Scholar] [CrossRef]
- Clay, L.A.; Wang, S.Y.; Wolters, W.R.; Peterson, B.C.; Waldbieser, G.C. Molecular characterization of the insulin-like growth factor-I (IGF-1) gene in channel catfish (Ictalurus punctatus). Biochim. Biophys. Acta 2005, 1731, 139–148. [Google Scholar] [CrossRef] [PubMed]
- A Marca Pereira, M.L.; Eppler, E.; Thorpe, K.L.; Wheeler, J.R.; Burkhardt-Holm, P. Molecular and cellular effects of chemicals disrupting steroidogenesis during early ovarian development of brown trout (Salmo trutta fario). Environ. Toxicol. 2014, 29, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Eppler, E.; Caelers, A.; Shved, N.; Hwang, G.; Rahman, A.M.; Maclean, N.; Zapf, J.; Reinecke, M. Insulin-like growth factor I (IGF-I) in a growth-enhanced transgenic (GH-overexpressing) bony fish, the tilapia (Oreochromis niloticus): Indication for a higher impact of autocrine/paracrine than of endocrine IGF-I. Transgenic Res. 2007, 16, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, A.; Riesen, I.; Köllner, B.; Eppler, E.; Segner, H. Surface marker-defined head kidney granulocytes and B lymphocytes of rainbow trout express Benzo[a]pyrene-inducible cytochrome P4501A protein. Toxicol. Sci. 2008, 103, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Eppler, E.; Shved, N.; Moret, O.; Reinecke, M. IGF-I is distinctly located in the bony fish pituitary as revealed for Oreochromis niloticus, the Nile tilapia, using real-time RT-PCR, in situ hybridisation and immunohistochemistry. Gen. Comp. Endocrinol. 2007, 150, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, A.; Okamoto, N.; Fukuda, H. Characterization of monoclonal antibodies against antigens shared with neutrophils and macrophages in rainbow trout (Oncorhynchus mykiss). Fish Pathol. 2000, 35, 205–213. [Google Scholar] [CrossRef]
- Marinova, T.T.; Kuerten, S.; Petrow, D.B.; Angelov, D.N. Thymic epithelial cells of human patients affected by myasthenia gravis overexpress IGF-I immunoreactivity. APMIS 2008, 116, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Law, H.K.; Tu, W.; Liu, E.; Lau, Y.L. Insulin-like growth factor I promotes cord blood T cell maturation through monocytes and inhibits their apoptosis in part through interleukin-6. BMC Immunol. 2008, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eppler, E.; Janas, E.; Link, K.; Weidmann, L.; Bischofberger, H.; Wenger, M.; Tinguely, M.; Schraml, P.; Moch, H.; Fellbaum, C. Insulin-like growth factor-I is expressed in classical and nodular lymphocyte-predominant Hodgkin’s lymphoma tumour and microenvironmental cells. Cell Tissue Res. 2015, 359, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Arkins, S.; Rebeiz, N.; Biragyn, A.; Reese, D.L.; Kelley, K.W. Murine macrophages express abundant insulin-like growth factor-I class I Ea and Eb transcripts. Endocrinology 1993, 133, 334–343. [Google Scholar]
- Oberlin, D.; Fellbaum, C.; Eppler, E. Insulin-like growth factor I messenger RNA and protein are expressed in the human lymph node and distinctly confined to subtypes of macrophages, antigen-presenting cells, lymphocytes and endothelial cells. Immunology 2009, 128, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Duguay, S.J.; Swanson, P.; Dickhoff, W.W. Differential expression and hormonal regulation of alternatively spliced IGF-I mRNA transcripts in salmon. J. Mol. Endocrinol. 1994, 12, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Duguay, S.J.; Lai-Zhang, J.; Steiner, D.F.; Funkenstein, B.; Chan, S.J. Developmental and tissue regulated expression of IGF-I and IGF-II mRNAs in Sparus aurata. J. Mol. Endocrinol. 1996, 16, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Otteson, D.C.; Cirenza, P.F.; Hitchcock, P.F. Persistent neurogenesis in the teleost retina: Evidence for regulation by the growth-hormone/insulin-like growth factor-I axis. Mech. Dev. 2002, 117, 137–149. [Google Scholar] [CrossRef]
- Vong, Q.P.; Chan, K.M.; Cheng, C.H. Quantification of common carp (Cyprinus carpio) IGF-I and IGF-II mRNA by real-time PCR: Differential regulation of expression by GH. J. Endocrinol. 2003, 178, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Biga, P.R.; Schelling, G.T.; Hardy, R.W.; Cain, K.D.; Overturf, K.; Ott, T.L. The effects of recombinant bovine somatotropin (rbST) on tissue IGF-I, IGF-I receptor, and GH mRNA levels in rainbow trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol. 2004, 135, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Gahr, S.A.; Vallejo, R.L.; Weber, G.M.; Shepherd, B.S.; Silverstein, J.T.; Rexroad, C.E., III. Effects of short-term growth hormone treatment on liver and muscle transcriptomes in rainbow trout (Oncorhynchus mykiss). Physiol. Genomics 2008, 32, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Fukada, H.; Ozaki, Y.; Pierce, A.L.; Adachi, S.; Yamauchi, K.; Hara, A.; Swanson, P.; Dickhoff, W.W. Salmon growth hormone receptor: Molecular cloning, ligand specificity, and response to fasting. Gen. Comp. Endocrinol. 2004, 139, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Very, N.M.; Kittilson, J.D.; Norbeck, L.A.; Sheridan, M.A. Isolation, characterization, and distribution of two cDNAs encoding for growth hormone receptor in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. B 2005, 140, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Jiao, B.; Huang, X.; Chan, C.B.; Zhang, L.; Wang, D.; Cheng, C.H. The co-existence of two growth hormone receptors in teleost fish and their differential signal transduction, tissue distribution and hormonal regulation of expression in seabream. J. Mol. Endocrinol. 2006, 36, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cueto, J.A.; Martínez-Barbera, J.P.; Pendón, C.; Rodríguez, R.B.; Valdivia, M.M.; Sarasquete, M.C. Autoradiographic localization of growth hormone receptors in tissues of the gilthead sea bream, Sparus aurata. In Proceedings of the 17th Conference of European Comparative Endocrinologists, From Basic to Applied Aspects, Córdoba (Spain), 5–10 September 1994. Abstract 237.
- Nagasawa, T.; Nakayasu, C.; Rieger, A.M.; Barreda, D.R.; Somamoto, T.; Nakao, M. Phagocytosis by Thrombocytes is a Conserved Innate Immune Mechanism in Lower Vertebrates. Front. Immunol. 2014, 5, 445. [Google Scholar] [CrossRef] [PubMed]
- Beckman, B.R. Perspectives on concordant and discordant relations between insulin-like growth factor 1 (IGF1) and growth in fishes. Gen. Comp. Endocrinol. 2011, 170, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Deane, E.E.; Woo, N.Y. Modulation of beta-actin, insulin-like growth factor 1 and glucose-6-phosphate dehydrogenase gene expression during vibriosis of sea bream, Sparus (=Rhabdosargus) sarba Forsskål. J. Fish Dis. 2005, 28, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Saera-Vila, A.; Calduch-Giner, J.A.; Prunet, P.; Pérez-Sánchez, J. Dynamics of liver GH/IGF axis and selected stress markers in juvenile gilthead sea bream (Sparus aurata) exposed to acute confinement: Differential stress response of growth hormone receptors. Comp. Biochem. Physiol. A 2009, 154, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Pinard, M.; Wang, Y.; Yang, L.; Lin, R.; Hiscott, J.; Su, B.; Brodt, P. Crosstalk between the TNF and IGF pathways enhances NF-κB activation and signaling in cancer cells. Growth Horm IGF Res. 2015, 25, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Noble, P.W.; Lake, F.R.; Henson, P.M.; Riches, D.W.H. Hyaluronate activation of CD44 induces insulin-like gowth factor-1 expression by a tumor necrosis factor-alpha dependent mechanism in murine macrophages. J. Clin. Investig. 1993, 91, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Fournier, T.; Riches, D.W.; Winston, B.W.; Rose, D.M.; Young, S.K.; Noble, P.W.; Lake, F.R.; Henson, P.M. Divergence in macrophage insulin-like growth factor-I (IGF-I) synthesis induced by TNF-alpha and prostaglandin E2. J. Immunol. 1995, 155, 2123–2133. [Google Scholar] [PubMed]
- O’Connor, J.C.; Cusker, R.H.; Strle, K.; Johnson, R.W.; Dantzer, R.; Kelley, K.W. Regulation of IGF-I function by proinflammatory cytokines: At the interface of immunology and endocrinology. Cell. Immunol. 2008, 252, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Palin, K.; Bluthé, R.M.; McCusker, R.H.; Moos, F.; Dantzer, R.; Kelley, K.W. TNFalpha-induced sickness behavior in mice with functional 55 kD TNF receptors is blocked by central IGF-I. J. Neuroimmunol. 2007, 187, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Meazza, C.; Elsedfy, H.H.; Pagani, S.; Bozzola, E.; El Kholy, M.; Bozzola, M. Metabolic parameters and adipokine profile in growth hormone deficient (GHD) children before and after 12-month GH treatment. Horm. Metab. Res. 2014, 46, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Uronen-Hansson, H.; Allen, M.L.; Lichtarowicz-Krynska, E.; Aynsley-Green, A.; Cole, T.J.; Höidén-Guthenberg, I.; Fryklund, L.; Klein, N. Growth hormone enhances proinflammatory cytokine production by monocytes in whole blood. Growth Horm IGF Res. 2003, 13, 282–286. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T.; Hong, S.; Peddie, S.; Crampe, M.; Laing, K.J.; Cunningham, C.; Zou, J. Cytokines and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 713–723. [Google Scholar] [CrossRef]
- Goetz, F.W.; Planas, J.V.; MacKenzie, S. Tumor necrosis factors. Dev. Comp. Immunol. 2004, 28, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.H.; Moon, J.Y.; Kim, Y.O.; Kong, H.J.; Kim, W.J.; Kim, K.K.; Lee, S.J. Molecular and functional analyses of growth hormone-releasing hormone (GHRH) from olive flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. B 2011, 159, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Wenger, M.; Sattler, U.; Goldschmidt-Clermont, E.; Segner, H. 17Beta-estradiol affects the response of complement components and survival of rainbow trout (Oncorhynchus mykiss) challenged by bacterial infection. Fish Shellfish Immunol. 2011, 31, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Wenger, M.; Krasnov, A.; Skugor, S.; Goldschmidt-Clermont, E.; Sattler, U.; Segner, H. Estrogen modulates hepatic gene expression and survival of rainbow trout infected with pathogenic bacteria Yersinia ruckeri. Mar. Biotechnol. 2012, 14, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Leatherland, J.F. The interaction between maternal stress and the ontogeny of the innate immune system during teleost embryogenesis: Implications for aquaculture practice. J. Fish Biol. 2012, 81, 1793–1814. [Google Scholar] [CrossRef] [PubMed]
- Seemann, F.; Knigge, T.; Rocher, B.; Minier, C.; Monsinjon, T. 17β-estradiol induces changes in cytokine levels in head kidney and blood of juvenile sea bass (Dicentrarchus labrax, L., 1758). Mar. Environ. Res. 2013, 87–88, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Raida, M.K.; Holten-Andersen, L.; Buchmann, K. Association between Yersinia ruckeri infection, cytokine expression and survival in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2011, 30, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish Shellfish Immunol. 2013, 35, 1703–1718. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franz, A.-C.; Faass, O.; Köllner, B.; Shved, N.; Link, K.; Casanova, A.; Wenger, M.; D’Cotta, H.; Baroiller, J.-F.; Ullrich, O.; et al. Endocrine and Local IGF-I in the Bony Fish Immune System. Biology 2016, 5, 9. https://doi.org/10.3390/biology5010009
Franz A-C, Faass O, Köllner B, Shved N, Link K, Casanova A, Wenger M, D’Cotta H, Baroiller J-F, Ullrich O, et al. Endocrine and Local IGF-I in the Bony Fish Immune System. Biology. 2016; 5(1):9. https://doi.org/10.3390/biology5010009
Chicago/Turabian StyleFranz, Anne-Constance, Oliver Faass, Bernd Köllner, Natallia Shved, Karl Link, Ayako Casanova, Michael Wenger, Helena D’Cotta, Jean-François Baroiller, Oliver Ullrich, and et al. 2016. "Endocrine and Local IGF-I in the Bony Fish Immune System" Biology 5, no. 1: 9. https://doi.org/10.3390/biology5010009