Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria

1
Department of Veterinary Biosciences, University of Helsinki, P.O. Box 66, FI-00014 Helsinki, Finland
2
General Microbiology, Department of Biosciences, University of Helsinki, P.O. Box 56, FI-00014 Helsinki, Finland
*
Author to whom correspondence should be addressed.
Biology 2014, 3(1), 178-204; https://doi.org/10.3390/biology3010178
Submission received: 8 January 2014
/
Revised: 26 February 2014
/
Accepted: 27 February 2014
/
Published: 10 March 2014
(This article belongs to the Special Issue Bacterial Adhesion)
Abstract
:Biological moonlighting refers to proteins which express more than one function. Moonlighting proteins occur in pathogenic and commensal as well as in Gram-positive and Gram-negative bacteria. The canonical functions of moonlighting proteins are in essential cellular processes, i.e., glycolysis, protein synthesis, chaperone activity, and nucleic acid stability, and their moonlighting functions include binding to host epithelial and phagocytic cells, subepithelia, cytoskeleton as well as to mucins and circulating proteins of the immune and hemostatic systems. Sequences of the moonlighting proteins do not contain known motifs for surface export or anchoring, and it has remained open whether bacterial moonlighting proteins are actively secreted to the cell wall or whether they are released from traumatized cells and then rebind onto the bacteria. In lactobacilli, ionic interactions with lipoteichoic acids and with cell division sites are important for surface localization of the proteins. Moonlighting proteins represent an abundant class of bacterial adhesins that are part of bacterial interactions with the environment and in responses to environmental changes. Multifunctionality in bacterial surface proteins appears common: the canonical adhesion proteins fimbriae express also nonadhesive functions, whereas the mobility organelles flagella as well as surface proteases express adhesive functions.
1. Introduction—Concept of Moonlighting Proteins
Moonlighting proteins include an expanding class of adhesion proteins in both prokaryotic and eukaryotic organisms. They are multifunctional and go against the general one gene-one protein-one function hypothesis. Moonlighting proteins possess two or more separate functions that cannot be ascribed to gene fusions, splice variants, or protein fragments expressing different functions after proteolysis (see the excellent review by Copley [1]). Moonlighting offers an individual protein a mechanism to increase network complexity and to participate in apparently unrelated cellular functions [2]. Moonlighting proteins were first described in eukaryotic cell biology 25 years ago, with the identification of the cytokine neuroleukin as phosphoglucose isomerase (PGI) and of a lens structural protein as enolase, i.e., as cytoplasmic enzymes of glucose metabolism [3,4]. Extracellular production of a glycolytic enzyme, invertase, in several yeast species was reported already in 1960s [5]. In bacteria, the first identified moonlighting enzyme was glyceraldehyde 3-phosphate dehydrogenase (GAPDH)—also a glycolytic enzyme—that was found in the cytoplasm as well as on the cell surface of group A streptococci. This bacterium, Streptococcus pyogenes, inhabits human throat and skin and is responsible for a variety of non-invasive and invasive infections, and GAPDH was found to exhibit in vitro multiple adhesive functions with a potential to increase bacterial virulence [6]. Since then, several moonlighting proteins have been identified or suggested in eukaryotic and prokaryotic organisms. In bacteria alone, the current number of protein species with an identified adhesive moonlighting function exceeds 20, and the on-going proteomic studies will undoubtedly reveal many more. As discussed in this review, practically all enzymes in the Embden-Mayerhof glycolytic pathway of bacteria have been assigned with an adhesive moonlighting function. In addition to adhesiveness, bacterial moonlighting proteins may influence other aspects of bacterial physiology. The glycolytic enzymes also moonlight by forming complexes with themselves as well as with proteins that function in RNA processing and such complexes are thought to potentiate metabolic channelling and glycolytic fluxes within the cell [7].
Moonlighting proteins often perform their canonical and moonlighting functions in separate cell compartments, e.g., the bacterial cytoplasm and the cell surface, and therefore such a dual cellular localization of a protein is suggestive of moonlighting. This is a limited criterium but has remained common in identification of a bacterial moonlighting protein, especially in proteomic studies. Detailed analyses of genes encoding moonlighting proteins, the proteins and their domains as well as functions in bacteria are few, whereas in vitro studies demonstrating unexpected adherence or binding partners for a moonlighting protein are more numerous. Here, we will address those bacterial moonlighting proteins that have been characterized for an adhesive function as well as for a cellular location. Our main emphasis is on their interactions with mammalian host components as well as on their anchorage on the bacterial surface. For general reviews on moonlighting proteins, the reader is referred to recent reviews [1,2,8,9].
In bacteria, the first moonlighting proteins were identified in pathogenic Gram-positive bacteria and shown to have pathogenesis-related functions, which stressed their importance as putative virulence factors (reviewed by Henderson and Martin [10]). However, recent research has also demonstrated moonlighting proteins in Gram-negative and in non-pathogenic or commensal bacteria as well. This indicates that moonlighting is common in bacteria. Most of the currently known moonlighting proteins and functions, however, come from Gram-positive bacteria (see Table 1), which may reflect their less complex cell wall structure and easier protein translocation. Bacteria differ in immobilization of moonlighting proteins on their surface which can seriously affect the function of the protein in different organisms (discussed below). A single bacterial strain can harbor several different moonlighting proteins, and a moonlighting protein species can express several distinct moonlighting functions. On the other hand, the same moonlighting function can be provided by different proteins [1]. Canonical bacterial adhesion proteins, such as the fimbriae, exhibit functions that are not directly involved in adherence to host tissues or cells, and, on the other hand, bacterial flagella as well as surface-associated proteases possess adhesive properties. These proteins are not generally included in discussion of bacterial moonlighting, probably because they are multimeric and perform their functions in the same cellular compartment, i.e., the bacterial surface. Their diverse functions however illustrate that multitasking is common in bacterial surface proteins, and we will end our review on a short description of fimbriae, flagella, and surface proteases.
2. Bacterial Moonlighting Proteins Come in Many Forms
Table 1 lists characterized bacterial moonlighting proteins with adhesive interactions, their organisms, and their primary and moonlighting functions. In general, moonlighting proteins in a bacterial species have been studied individually, independently from those in other bacterial species, which complicates obtaining a comprehensive and mechanistic view on bacterial moonlighting. The currently known bacterial moonlighting proteins have their canonical function in essential cellular processes, such as glycolysis, chaperone activity, protein synthesis, and nucleic acid stability (Table 1). This means that silencing of a moonlighting-associated gene most likely has broad phenotypic effects that can be hard to interpret in terms of either the canonical or the moonlighting function. The adhesive moonlighting functions that have been recognized thus far are diverse and include binding to secreted mucins, epithelial cells, lymphocytes and monocytes, extracellular matrices, circulating effector molecules, and other microbes (Table 1). The physiological significance and biological consequences of these interactions are known only in few cases. The structure of most bacterial moonlighting proteins, such as enolase and GAPDH, are highly conserved across the kingdoms and do not markedly differ in prokaryotes or eukaryotes, which is understandable in view of their essential metabolic functions and the fact that they moonlight in both organisms. Our limited knowledge of structure-function relationships in bacterial moonlighting proteins has limited the prediction of possible moonlighting functions in proteins; however, bioinformatic approaches for such predictions have been initiated [11,12].

{kind=link}
{kind=link}
{kind=link}
| Moonlighting protein | Moonlighting function | Bacterial species | Reference |
|---|---|---|---|
| Metabolic enzymes | |||
| GAPDH | |||
| Binding to plasmin(ogen) | B. anthracis, E. coli, La. crispatus, La. plantarum, Li. monocytogens, S. aureus, St. agalactiae, St. anginosus, St. epidermis, St. equisimilis, St. oralis, St. pneumoniae, St. pyogenes, St. suis | [6,13,14,15,16,17,18,19,20,21,22,23,24] | |
| Binding to urokinase receptor on human pharyngeal cells | St. pyogenes | [25] | |
| Binding to lysozyme | St. pyogenes | [6] | |
| Binding to actin | St. pyogenes, St. agalactiae | [6,22] | |
| Binding to myosin | St. pyogenes | [6] | |
| Binding to albumin | St. suis | [26] | |
| Binding to fibrinogen | St. agalactiae | [22] | |
| Binding to fibronectin | E. coli, L. plantarum, St. pyogenes | [6,14,16,27] | |
| Binding to other bacterial species | Group B Streptococcus, La. crispatus | [28,29] | |
| Binding to intestinal epithelial cells, competitive exclusion and displacement of Clostridium sporogenes and Enterococcus faecalis | La. plantarum | [30] | |
| Coadhesin of Porfyromonas gingivalis in periodontal tissue | St. oralis | [31] | |
| Binding to intestinal epithelial cells | E. coli, La. plantarum | [14,16] | |
| Binding to colonic, porcine or vaginal mucin | La. plantarum, M. genitalium | [16,32,33] | |
| Binding to A and B blood antigens | La. plantarum | [34] | |
| Binding to porcine tracheal rings | St. suis | [35] | |
| EGF-receptor | M. avium, M. tuberculosis | [36] | |
| Binding to C5a complement protein | St. pyogenes | [37] | |
| Enolase | |||
| Binding to plasmin(ogen) | B. anthracis, Bi. animalis subspecies lactis, Bi. bifidum, Bi. breve, Bi. longum, Bo. burgdorferi, La. crispatus, La. johnsonii, La. plantarum, Li. monocytogenes, M. pneumoniae, S. aureus, St. anginosus, St. mutans, St. oralis, S. pneumoniae, S. pyogenes | [16,17,19,21,38,39,40,41,42,43,44,45,46,47,48] | |
| Binding to fibronectin | La. plantarum | [16,49] | |
| Binding to laminin | La. crispatus, La. johnsonii, S. aureus | [48,50] | |
| Binding to collagen | La. crispatus, S. aureus | [48] | |
| Binding to albumin | St. pyogenes | [51] | |
| Binding to salivary mucin | St. gordonii, St. mutans | [42,52] | |
| Binding to intestinal epithelial cells | La. plantarum, St. suis | [16,53] | |
| Binding to C4b-binding proteins | St. pneumoniae | [54] | |
| Binding to other bacterial species | La. crispatus | [28] | |
| Aldolase | |||
| Binding to flamingo cadherin | St. pneumoniae | [55] | |
| Glucose-6-phosphate isomerase (GPI) | |||
| Binding to collagen | La. crispatus | [28] | |
| Binding to other bacterial species | La. crispatus | [28] | |
| Phosphofructokinase | |||
| Binding to plasmin(ogen) | St. oralis | [19] | |
| Phosphoglycerate kinase | |||
| Binding to plasmin(ogen) | St. anginosus, St. oralis, Group B streptococci | [19,56] | |
| Binding to actin | Group B streptococci | [56,57] | |
| Phosphoglycerate mutase | |||
| Binding to plasmin(ogen) | Bi. animalis subspecies lactis, St. anginosus, St. oralis | [19,58] | |
| Triosephosphate isomerase | |||
| Binding to plasmin(ogen) | St. anginosus, St. oralis | [19] | |
| Binding to intestinal epithelial cells, competitive exclusion and displacement of Clostridium sporogenes and Enterococcus faecalis | La. plantarum | [30] | |
| Glutamine synthetase | |||
| Binding to plasmin(ogen) | Bi. animalis subspecies lactis, La. crispatus, M. tuberculosis | [28,58,59] | |
| Binding to fibronectin | M. tuberculosis | [59] | |
| Binding to collagen I and laminin | La. crispatus | [28] | |
| Binding to other bacterial species | La. crispatus | [28] | |
| Ribonucleotide reductase | |||
| Binding to plasmin(ogen) | S. aureus | [44] | |
| Inosine 5′-monophosphate dehydrogenase (IMPDH) | |||
| Binding to plasmin(ogen) | S. aureus | [44] | |
| Alcohol acetaldehyde dehydrogenase | |||
| Binding to Caco-2 cells | Li. monocytogenes | [60] | |
| Binding to eukaryotic Hsp60 | Li. monocytogenes | [61] | |
| Malate synthase | |||
| Binding to laminin and fibronectin | M. tuberculosis | [62] | |
| SarA; oligopeptide-binding protein | |||
| Binding to salivary mucin | St. gordonii | [52] | |
| Pyruvate dehydrogenase | |||
| Binding to fibronectin | La. plantarum | [47] | |
| Binding to plasmin(ogen) | M. pneumoniae | [46] | |
| Binding to fibronectin | M. pneumoniae | [63] | |
| Puryvate kinase | |||
| Binding to salivary mucin | St. gordonii | [52] | |
| Bile salt hydrolase | |||
| Binding to plasmin(ogen) | Bi. animalis subspecies lactis | [58] | |
| Molecular chaperones | |||
| DnaK | |||
| Binding to plasmin(ogen) | Bi. animalis subspecies lactis, M. tuberculosis, Li. monocytogenes | [21,58,59,64] | |
| Stimulation of dendritic cell maturation by binding CCR5 | M. tuberculosis | [65] | |
| Competition with HIVfor CCR5 binding | M. tuberculosis | [65,66] | |
| Mediation of LAB adherence to yeast cells | L. lactis | [67] | |
| GroEL | |||
| Binding to intestinal HT-29 cells and mucus; stimulation of IL-8 secretion in human macrophages and HT-29 cells; aggregation of H. pylori cells | La. johnsonii | [68] | |
| Translational elongational factors | |||
| EF-Tu | |||
| Binding to plasmin(ogen) | Li. monocytogenes, Ps. aeruginosa | [21,69] | |
| Binding to plasma Factor H and Factor H-related protein 1 (FHR-1) | Ps. aeruginosa | [69] | |
| Binding to intestinal epithelial cells and HT-MTX-derived mucus | La. johnsonii | [70] | |
| Binding to salivary mucin | St. gordonii | [52] | |
| Binding to intestinal epithelial cells, competitive exclusion and displacement of Clostridium sporogenes and Enterococcus faecalis | La. plantarum | [30] | |
| Binding to fibronectin | M. pneumoniae | [63] | |
| EF-G | |||
| Binding to salivary mucin | St. gordonii | [52] | |
| Other proteins | |||
| Ag85 complex of M. tuberculosis | |||
| Binding of plasmi(ogen) | M. tuberculosis | [59] | |
| Binding to fibronectin | M. tuberculosis | [59] | |
| DNA-directed RNA polymerase beta´subunit | |||
| Binding to salivary mucin | St. gordonii | [52] | |
| Endopeptidase O | |||
| Binding of plasmin(ogen) | St. pneumoniae | [71] | |
| Binding to fibronectin | St. pneumoniae | [71] | |
| Binding and invasion to epithelial and endothelial cells | St. pneumoniae | [71] | |
| SecA | |||
| Binding to salivary mucin | St. gordonii | [52] | |
| Superoxide dismutase | |||
| Binding to epithelial cell ldolase, GAPDH and cyclophilin A | M. avidum | [72] | |
3. How Can the Separate Functions Be Arranged in a Moonlighting Protein?
Commonly, the canonical and the moonlighting functions are provided by separate parts of the protein. Alternatively, they can be partially overlapping or provided by alternative conformations of the protein. The latter may involve post-translational modifications, binding of ligands, a pH change, expression in a different cell type or a cellular location, immobilization on a surface, oligomerization, or other perturbation [1,51,73]. Binding of GAPDH of Lactobacillus plantarum to human colonic mucins is inhibited by exogenous NAD, which was inferred to indicate that the catalytic region of the enzyme is involved in the adhesion [32]. The well characterized enolase of the severe pathogen Streptococcus pneumoniae has multiple adhesive properties which include binding of the human circulating protease precursor, plasminogen [39]. Plasminogen harbors lysine-binding kringle domains, and the interaction with pneumococcal enolase apparently takes place via two pairs of lysines in enolase, whose mutations influence plasminogen binding [47,74,75]. The two lysines in the sequence 248FYNKDDHKY are exposed on the surface of the enolase octamer [76], and the mutations affect the enzymatic activity only partially or not at all, which suggests that the enzymatic and the adhesive functions do not involve overlapping regions. Amino acid sequences similar to this pneumococcal plasminogen-binding motif are present in other bacterial enolases as well [43,75]. For example, the genome of Lactobacillus johnsonii contains three copies of the enolase gene which are 50%–72% identical to each other and encode sequences homologous to the plasminogen-binding motif of pneumococcal enolase [77]. Denaturation of the octameric form of pneumococcal enolase exposes C-terminal lysine residues that also bind plasminogen, which gives an example of a bacterial moonlighting function depending on a variation of protein conformation. However, terminal lysine residues are not present in most of the enolase proteins of lactobacilli that, however, show binding to plasminogen [48,78]. Moonlighting enzymes are released from the surface of lactic acid bacteria under stress conditions (see below), and hence small conformational changes may be relevant in their biology. Kornblatt and coworkers [51] studied in vitro the interaction between plasminogen and Group A streptococcal enolase by a wealth of biophysical and biochemical analyses and concluded that the interaction was seen only when at least one of the partners was in a non-native state, which could be induced by a temporary pH change or immobilization of enolase in a phospholipid micelle. The non-native conformation of enolase bound also other proteins, such as albumin and enolases from other organisms [51]. Their study concluded that the moonlighting adhesiveness of enolase is not highly specific; which is in accordance with the multiple adhesion targets identified for bacterial enolases (Table 1). These results agree with the suggestion by Tompa and coworkers [2] that the structural flexibility of unstructured proteins or protein regions give rise to unprecedented molecular interactions that may lead to moonlighting. This hypothesis predicts that structural flexibility, not the structure per se, is critical for a moonlighting function in a protein.
It is possible that specific paralocous alleles of a gene encode moonlighting variants. Two enolase genes are present in the genome of L. plantarum [78] and both genes are expressed in standard growth conditions [49]. Enolase preparation from the surface of L. plantarum, apparently containing both enolases, bound plasminogen as well as fibronectin, [16] and mutagenesis studies showed that fibronectin-binding was performed by only one of the enolases [49]. The genome of M. tuberculosis contains two genes encoding the molecular chaperonins GroEL1 (also termed Cnp60.1) and GroEL2 (Cnp60.2), which both have impact on virulence of M. tuberculosis and induce host inflammatory responses [79]. Mutagenesis analyses revealed that GroEL2 is essential for survival of the bacteria whereas the cnp60.1 gene could be inactivated; the mutated strain failed to produce granulomatous inflammation in either mice or guinea pigs [80]. GroEL1 of M. tuberculosis is released into culture medium during bacterial growth [81] and GroEL2 was identified among capsule-associated proteins on the bacterial surface [82]. Inhibition assays using recombinant GroEL2 protein as well as anti-GroEL2 polyclonal antibody showed that GroEL2 facilitates association of M. tuberculosis with murine macrophages [76], the receptor was identified as the CD43 molecule, a major sialoglycoprotein on the surface of human T lymphocytes, monocytes, granulocytes and some B lymphocytes [83]. Purified GroEL1 and GroEL2 proteins induce cytokine release in vitro from human peripheral blood mononucleated cells but show differences in their potency and mechanism [80,81,84]. It is generally considered that alveolar macrophage is the primary host niche of M. tuberculosis [81]; the observed interactions of the GroEL proteins with monocytes indicate their importance as virulence factors in the pathogenesis of tuberculosis. These results suggest that the two enolase variants and the two GroEL variants are moonlighting proteins but express differing functions. Overall, the possible presence of specific moonlighting protein-encoding alleles in bacteria remains open.
4. Are Moonlighting Proteins Secreted or Are They Released from Traumatized Cells?
The moonlighting proteins have been called “surface-associated housekeeping enzymes”, “anchorless surface proteins”, or “unconvential secreted proteins”, which all refer to the fact that their sequences do not contain known sequence motifs for secretion or anchoring onto the bacterial or the eukaryotic cell surface [1,10,85]. The mechanisms of how moonlighting proteins translocate to the cell exterior have remained open. Alternatives are that they are released from dead or traumatized bacterial cells and then bind onto the surface of neighboring cells, or that they are secreted onto the cell surface by an as yet-undescribed mechanism. In bacteria, experimental evidence for both hypotheses have been described, but overall, the data remains scattered and only suggestive, also, the two alternatives are not fully mutually exclusive.
The list of moonlighting adhesion proteins (Table 1) is comprehensive, and the proteins do not show any obvious, over-all conformational relatedness that would explain their translocation to cell surface. The secretion hypothesis has been put forward by the observations that genetic fusion of a C-terminal hydrophobic tail of 12 amino acids to GAPDH prevented its export to the surface of S. pyogenes [86]. In enolase of Escherichia coli, substitution of Lys341 at an automodification site (see below) with other amino acids abolished its export to cell surroundings [87], whereas in Bacillus subtilis enolase deletion of a central hydrophobic α-helical domain of 19 amino acids abolished surface translocation [88]. Further, an accessory secA2 gene in Listeria monocytogenesis is involved in the secretion of enolase and other proteins [89], but a homologous gene is lacking in several bacterial species, such as lactobacilli that express moonlighting enolase and often have multiple gene copies. Enolases of E. coli, Enterococcus faecalis, and B. subtilis are partially automodified covalently by their substrate 2-phosphoglycerate, and in E. coli, an amino acid substitution at the modification site abolished translocation of enolase into the medium [87], which suggests that posttranslational modifications could have a role in translocation of moonlighting surface proteins.
There is also evidence suggesting that moonlighting proteins are released by stressed cells in a bacterial population and reassociate at favorable conditions onto the surface of the same or neighboring cells or other surfaces. Spontaneous lysis of B. subtilis cells at stationary growth phase leads to leakage of 5% of the activity of isocitrate dehydrogenase, considered to be a cytoplasmic enzyme marker [90]. Chloramphenicol or a proton motive force inhibitor did not inhibit translocation of several moonlighting proteins of B. subtilis into the stationary-phase growth medium [88]. In L. plantarum, the amount of surface-associated GAPDH activity correlates with the growth phase and with increase in membrane permeability [91], and growth of Bifidobacterium animalis subsp. lactis with bile acids increased levels of DnaK on the cell surface [64]. DnaK is a molecular chaperone with multiple adhesive and immunostimulatory moonlighting functions (Table 1). Binding at low pH of extracellular moonlighting proteins onto the surface of Lactobacillus crispatus as well as their release from L. crispatus surface by alkaline stress [28,92] have been reported. GAPDH of group B streptococci (GBS) was not released into medium by pilus mutants that display a lower level of cell lysis; on the other hand, factors enhancing cell lysis, such as Triton X-100, penicillin, and overexpression of an autolysin, enhanced the amount of GAPDH at the cell surface as well as in the culture supernatant [29]. These results favor the hypothesis that moonlighting proteins detach from damaged cell walls or leach from lysed cells, however, they do not explain how the moonlighting proteins are translocated to the cell wall.
To summarize, the translocation mechanism(s) of moonlighting proteins remain unknown, and we do not know whether one or more mechanisms are involved. The studies have been performed with different organisms and different proteins which complicates obtaining a mechanistic general view.
5. Adhesive Properties in Bacterial Moonlighting Proteins
The adhesive functions of moonlighting proteins include adhesion to host epithelial cells, mucus and extracellular matrix (ECM) components, as well as interaction with circulating host components and modulation of host immune responses (Table 1). It is interesting that, when the target surfaces (epithelial cells, mucus) or a limited number of target molecules (ECM proteins, circulating proteins) are considered, several adhesive moonlighting proteins exhibit similar in vitro functions in pathogenic and commensal bacteria (Table 1). This supports studies on the concept of surface exclusion of pathogens [93,94,95] by commensal bacteria, where the highly numerous bacteria in the microbiota prevent pathogen entry by competing for colonization and space on mucosal surfaces or for physiologically important circulating proteins. This has mostly been addressed with epithelial cell lines in vitro, where, e.g., L. plantarum, Lactobacillus fermentum and Lactobacillus jensenii were found to use moonlighting proteins in competitive exclusion and displacement of pathogens [30,96,97]. Whether the surface exclusion is based on recognition of the same receptor molecules or whether it is based on a less-specific steric hindrance remains yet open.
Another frequently studied adhesive function of bacterial moonlighting proteins is their ability to act as a plasminogen/plasmin receptor. Immobilization of plasminogen on bacterial surface, onto plasminogen receptors, enhances its proteolytic activation by human activators to plasmin, a potent serine protease, and also protects plasmin from inactivation by the circulating inhibitor α2-antiplasmin (α2AP) [98,99]. The process enhances surface-bound proteolytic activity which can advance bacterial survival or spread in the host [99]. Plasmin is broad spectrum protease which degrades fibrin, extracellular matrices, and activates human enzymes attacking connective tissue components [100]. The interaction with plasminogen may also increase bacterial adhesiveness. Plasminogen binds to bacterial cell surface receptors as well as to integrin molecules on eukaryotic cells and can thus also enhance bacterial adherence to host epithelia by a bridging mechanisms [101,102,103]. Plasminogen, but not plasmin, mediates in vitro adherence of S. pneumoniae to pulmonary epithelial and vascular endothelial cells. Transmigration of S. pneumoniae across endothelial and epithelial cell layers was potentiated by damage of intracellular junctions by active plasmin [104], which may promote the migration of pneumococci through tissue barriers. Plasminogen bound on surface of human respiratory epithelial cell lines and on brain-derived endothelial cells supported in vitro adherence of pneumococci but did not increase pneumococcal invasion into the target cells [105]. Plasminogen increased adherence as well as β-1integrin-dependent invasion of S. pyogenes to keratinocytes [102], and moonlighting plasminogen receptors are important virulence factors also for group A streptococci [86,106]. However, they also occur (Table 1) on a surprisingly high number of commensal bacteria species, where their biological functions remain largely unknown
5.1. GAPDH
The two best characterized bacterial moonlighting enzymes are GAPDH and enolase (Table 1). GAPDH is involved in bacterial adhesion to several host components: blood group antigens, the cytoskeleton proteins actin and myosin, the serum proteins albumin, fibrinogen and plasminogen, the ECM proteins, as well as colonic mucins. Targets for GAPDH also include epithelial and endothelial cells, and it has been associated with several physiological and immunological functions (Table 1). GAPDH of S. pyogenes has been identified as an ADP-ribosylating enzyme [102]. ADP-ribosylation of host components is common for bacterial toxins [103]. Streptococcal cells as well as surface GAPDH activate the protein tyrosine kinase and the protein kinase C in human pharyngeal cells, which indicates that GAPDH regulates phosphorylation of cellular proteins and may play a role in pathogenesis and in cellular communication between streptococci and the host [107].
Bacteria may also use GAPDH to evade and modulate the host immune system. GAPDH of S. pyogenes binds C5a and inhibits C5a-activated chemotaxis and H2O2 production by human neutrophils [37], and GAPDH of S. agalactiae is a virulence-associated immunomodulatory protein [108] that induces B and T cell activation of spleen lymphocytes in vitro. The elevated IL-10 production induced by recombinant GAPDH seems to be important for successful colonization of the host by Streptococcus agalactiae [108]. Also the GAPDH of S. pneumoniae was reported to be antigenic in humans, and vaccination of mice with recombinant GAPDH elicited protective immune response against respiratory challenge with virulent pneumococci [109].
Genetic evidence also suggests that GAPDH is a virulence factor for S. pyogenes. Fusion of a hydrophobic tail into the C-terminus of GAPDH prevented the surface translocation and diminished plasminogen binding to the bacteria as well as bacterial adhesion to pharyngeal cells [86]. The mutant strain was completely attenuated for virulence in a mouse peritonitis model, and surface expression of GAPDH thus seems mandatory for the virulence of S. pyogenes [106].
5.2. Enolase
Enolase has been reported to bind mucus and epithelial cells, but its best-described moonlighting activity is its function as a plasminogen receptor (Table 1). Invasion of pneumococci through basement membranes is thought to be important in meningitis, and plasminogen activation by host activators was found to potentiate pneumococcal penetration through the reconstituted basement membrane preparation Matrigel [110]. Soluble recombinant enolase binds in vitro at neutral pH to cell surface of S. pneumoniae when associated with plasminogen [39], and pneumococcal enolase-coated microspheres degraded radiolabeled ECM and Matrigel [111]. Also enolase of Bacillus anthracis binds plasminogen and bacteria incubated with plasminogen together with streptokinase (a plasminogen activator) were in vitro capable of degrading fibronectin [38]. Pneumococci expressing enolase substituted for the internal plasminogen binding site show reduced virulence in a mouse model of intranasal infection [74], and immunization of mice with enolase of S. suis protects against streptococcal infection [53].
It was recently found that pneumococci use enolase also to evade the host immune system [54]. Pneumococcal enolase binds C4b-binding protein (C4BP) and thereby protects bacteria from complement-mediated killing. Interestingly, plasminogen and C4BP bind to different domains of the enolase protein indicating that enolase has a dual role in pneumococcal pathogenesis. Immunization of rats with enolase from the oral pathogen Streptococcus sobrinus increased the levels of salivary IgA and IgG antibodies, and recombinant enolase was considered a promising candidate as a vaccine against dental caries in humans [112].
5.3. Molecular Chaperones
Molecular chaperones are a major class of bacterial moonlighting proteins. Their canonical function is to bind and assist folding or unfolding of macromolecular structures, and, given the prevailing nature of moonlighting in bacteria, it is not surprising that a variety of moonlighting functions have been identified for molecular chaperones. The bacterial Hsp70 protein, DnaK, was identified in the cell wall proteome of L. monocytogens [21]. Moonlighting functions of DnaK have been intensively studied in Mycobacterium tuberculosis. Recombinant DnaK binds human plasminogen [59], stimulates monocyte chemokine synthesis by binding to the CD40 receptor [113] and dendritic cell maturation by binding to chemokine receptor CCR5 [65]. CCR5 is also used by HIV as a co-receptor, and by binding the same receptor DnaK can compete with HIV for the binding [65,66]. Virus infection of activated CD4+ T cells was inhibited by the C-terminal peptide 407–426, which can have potential as a novel strategy to block the binding of HIV to its receptor [66]. As in M. tuberculosis, GroEL of L. johnsonii has an immunomodulatory activity and stimulates interleukin-8 secretion in human macrophages in vitro. It also aggregates cells of the human-specific gastric pathogen Helicobacter pylori [68] and thus can enhance clearance of the pathogen from gastric mucosa.
6. Moonlighting Proteins of Lactobacilli: Ionic Interactions Are Important in Anchorage and Activity
Species of Lactobacillus are fermentative and secrete lactic acid as a primary metabolite and thereby rapidly acidify their environment down to pH 4. These bacteria commonly inhabit acidic environments, such as human small intestine and vagina [114]. Several moonlighting proteins have been identified in species of Lactobacillus, these include GAPDH, enolase, glucose-6-phosphate isomerase (GPI), triose phosphate isomerase, phosphoglycerate kinase, elongation factor Tu, glutamine synthetase (GS) as well as GroEL [17,27,28,30,32,34,49,68,115]. These proteins exhibit a wealth of adhesive properties (Table 1) whose biological relevance, however, remain largely unidefined.
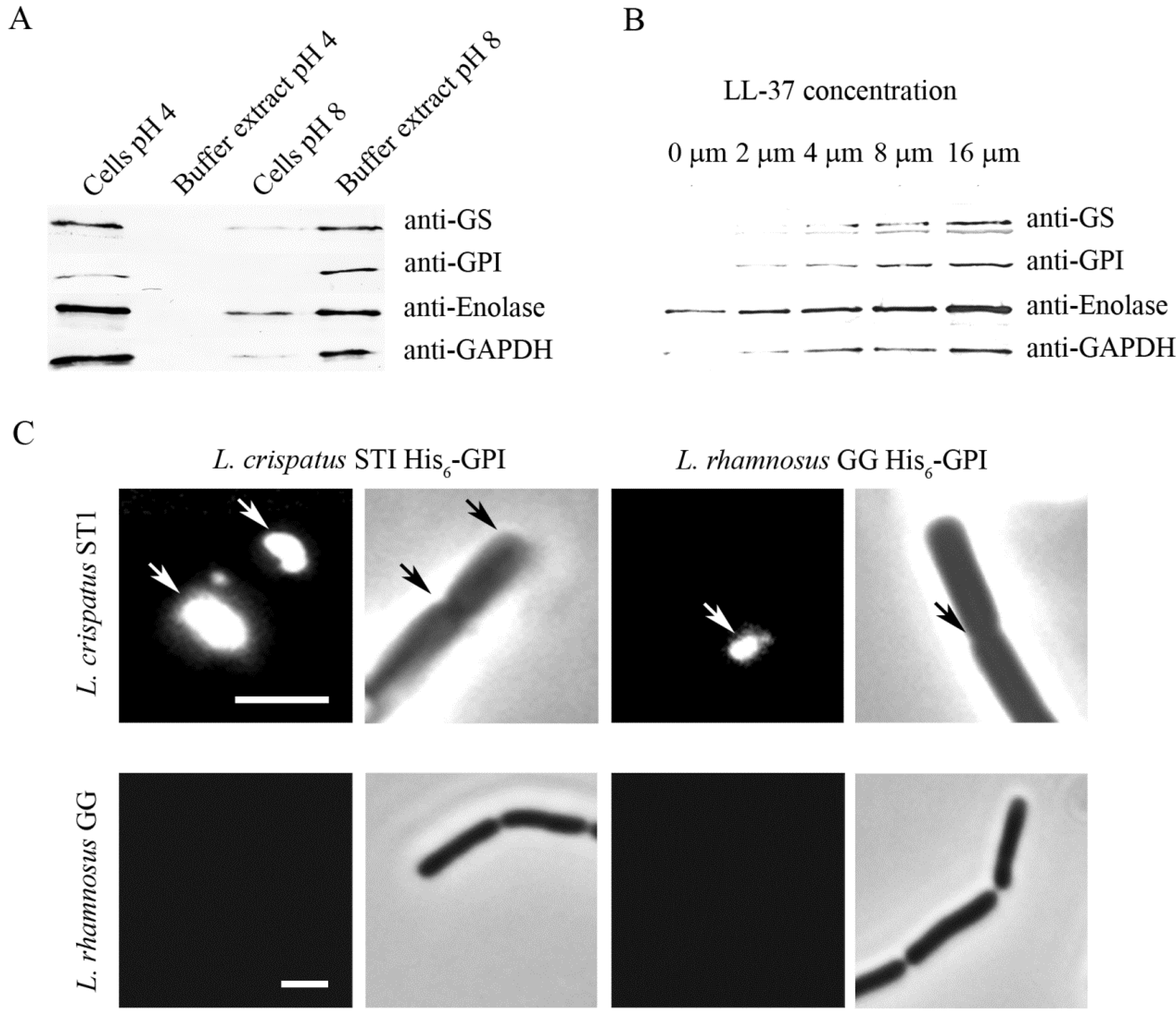
Recent in vitro evidence indicates that ionic interactions are critical in surface anchorance and in adherence activity of moonlighting proteins of lactobacilli. L. crispatus inhabits human vagina where it is thought to exert beneficial, probiotic effects and diminish inflammatory vaginosis, possibly through adhesion-associated mechanism [116]. The strain ST1 of L. crispatus shows pH-dependent adhesion to Matrigel in vitro (Figure 1A) [28] and in binding to human plasminogen [92]. The low adhesiveness at neutral or basic pH seems to depend on two aspects, (i) the anchoring of the adhesins on the cell surface, and (ii) their binding activity. First, several adhesive moonlighting proteins, i.e., enolase, GAPDH, GS, and GPI of L. crispatus ST1 are detached from cell surface at neutral or basic pH but remain surface-bound at pH 4 (Figure 2A) [17,28,92]. This seems to also hold for several other species of Lactobacillus [17,47]. These proteins have isoelectric points around 5 and thus a positive net charge at pH values below 5, a condition where purified enolase and GAPDH of L. crispatus ST1 bind in vitro to negatively charged lipoteichoic acids as well as to the L. crispatus ST1 cell surface [28,92]. Lipoteichoid acids are common surface constituents in Gram-positive bacteria, and the results suggest that they may anchor moonlighting proteins on the bacterial surface via ionic interactions. The role of ionic interactions in the anchoring is further indicated by the findings that moonlighting proteins are released from the L. crispatus ST1 surface in 0.25 mM sodium or choline chloride [92] and that the release is increased by μM concentrations of LL-37 (Figure 2B) [28]. LL-37 is a cationic antimicrobial peptide that is part of innate immune defense against bacteria and produced mainly by epithelial cells and phagocytes [117]. Contrary to moonlighting proteins of L. crispatus ST1, the abundant S-layer proteins of L. crispatus and other lactobacilli have a pI of ca. 10 [118], and they are not released from cell surface at neutral or basic pH [92].
Figure 1.
Adherence of L. crispatus ST1 and binding of His6-GS of ST1 are affected by pH. (A) Adherence of L. crispatus ST1 at pH 4 and pH 8 to the immobilized basement membrane preparation Matrigel and to BSA. Prior to the assays, the bacteria from an overnight culture were washed with the pH 4 or the pH 8 buffer, and the adhesion assays were performed at the corresponding pH. Means of adherent bacteria in 20 randomly chosen microscopic fields of 1.6 × 104 μm2 and standard deviations are shown. (B) Binding of purified His6-GS to immobilized laminin was tested by surface plasmon resonance at pH 5.5, 6.0 and 6.5 (Adapted from [28] Copyright© American Society for Microbiology).
Figure 1.
Adherence of L. crispatus ST1 and binding of His6-GS of ST1 are affected by pH. (A) Adherence of L. crispatus ST1 at pH 4 and pH 8 to the immobilized basement membrane preparation Matrigel and to BSA. Prior to the assays, the bacteria from an overnight culture were washed with the pH 4 or the pH 8 buffer, and the adhesion assays were performed at the corresponding pH. Means of adherent bacteria in 20 randomly chosen microscopic fields of 1.6 × 104 μm2 and standard deviations are shown. (B) Binding of purified His6-GS to immobilized laminin was tested by surface plasmon resonance at pH 5.5, 6.0 and 6.5 (Adapted from [28] Copyright© American Society for Microbiology).

At pH 4, but not at pH 8, purified His6-tagged GS and GPI showed a localized in vitro binding onto L. crispatus ST1 cells and bound to cell division sites and cell poles (Figure 2C) [28]. The cell division septa have structural and functional specifics [119,120,121] and it is tempting to speculate that moonlighting proteins are translocated to cell surface via these structures, either by diffusion or by a transport system. It is interesting that moonlighting proteins of L. crispatus ST1 and of the well-studied probiotic Lactobacillus rhamnosus GG bound in vitro to L. crispatus ST1 cells but not to L. rhamnosus GG cells (Figure 2C) [28]. The basis of this difference remains to be defined but the observation, however, suggests that differences in cell surface architecture and chemical composition influence the amount of moonlighting proteins bound on bacterial cell surfaces.
Another reason for poor adhesiveness at pH 8 by L. crispatus ST1 cells (Figure 1A) is that the moonlighting proteins of L. crispatus ST1 bind more efficiently at low pH [28]. Matrigel is composed of mainly laminin and type IV collagen [122], and it was found that purified His6-GS of L. crispatus ST1 bound to laminin much better at pH 5.5 than at pH 6.5 (Figure 1B) [28]. Further, His6-tagged GS and GPI showed a similar pH-dependency in binding to type I collagen, which is the most abundant protein of mammalian ECMs, but failed to bind to the ECM proteins type IV collagen and fibronectin. Increased binding at acidic pH has been also observed for mucus-binding proteins of Lactobacillus reuteri [123] and in the binding of enolase and GAPDH of L. plantarum [16] as well as of EF-Tu [70] and GroEL [68] of L. johnsonii to epithelial cells and mucins.
Figure 2.
Release of moonlighting proteins from L. crispatus ST1 surface and reassociation of recombinant proteins onto the cell wall. (A) Western blotting of GS, GPI, enolase, and GAPDH from the surface of L. crispatus ST1 cells and from the cell-free buffer. The cells were incubated at pH 4 or pH 8 for 1 h, and the moonlighting proteins in neutralized samples were visualized by immunoblotting. (B) Release of moonlighting proteins in the presence of increasing concentrations of the cationic peptide LL-37. L. crispatus ST1 cells were treated with LL-37 at the indicated concentrations, and the moonlighting proteins in the buffer were visualized by Western blotting. (C) Binding of His6-GPI proteins of L. crispatus ST1 and L. rhamnosus GG to bacterial cells at pH 4. The binding was visualized by indirect immunofluorescence, phase contrast images of the microscopic field are also shown. The arrows indicate protein binding to bacterial cell surface. Size bar, 3 µm. (Adapted from [28] Copyright© American Society for Microbiology).
Figure 2.
Release of moonlighting proteins from L. crispatus ST1 surface and reassociation of recombinant proteins onto the cell wall. (A) Western blotting of GS, GPI, enolase, and GAPDH from the surface of L. crispatus ST1 cells and from the cell-free buffer. The cells were incubated at pH 4 or pH 8 for 1 h, and the moonlighting proteins in neutralized samples were visualized by immunoblotting. (B) Release of moonlighting proteins in the presence of increasing concentrations of the cationic peptide LL-37. L. crispatus ST1 cells were treated with LL-37 at the indicated concentrations, and the moonlighting proteins in the buffer were visualized by Western blotting. (C) Binding of His6-GPI proteins of L. crispatus ST1 and L. rhamnosus GG to bacterial cells at pH 4. The binding was visualized by indirect immunofluorescence, phase contrast images of the microscopic field are also shown. The arrows indicate protein binding to bacterial cell surface. Size bar, 3 µm. (Adapted from [28] Copyright© American Society for Microbiology).
Saad and coworkers [91] reported that, at acidic pH, the amount of surface-associated GAPDH on L. plantarum cells correlates with stationary growth phase and increased cell permeability. Similarly, stress by alkaline pH or by LL-37 increases cell wall permeability and release of moonlighting proteins from the surface of L. crispatus ST1 cells (Figure 2B) [28]. LL-37 is a cationic antimicrobial peptide produced by epithelial cells and phagocytes and disrupts bacterial cell wall integrity by binding to lipid molecules [124]; whether it merely facilitates release of moonlighting proteins from traumatized cells or whether it also potentiates their transfer to cell surface, remains to be studied. Stress induced by growth of bifidobacteria in the presence of bile acids [64] and of streptococci under iron starvation [125] increases the amount of moonlighting proteins in the cell exterior. These findings indicate that stress is a determinant of the translocation of moonlighting proteins in several Gram-positive bacteria.
7. Adhesive Moonlighting Goes in Several Directions: Multitasking in Fimbriae, Flagella, and Surface Proteases
Fimbrial filaments are composed of hundreds of copies of the major fimbrillin subunit and a few copies, often located at tip of the filament, of minor proteins that are responsible for the adhesive, lectin-like characteristics of the fimbrial types [126,127]. Several fimbrial types of E. coli and Salmonella enterica immobilize plasminogen on the bacterial surface in a lysine-sensitive manner and enhance its activation, thus localizing the bacterium-plasmin complex on target tissues recognized by the bacterial lectin [128,129,130]. The molecular details of these interactions have remained mechanistically uncharacterized; it is however well established that fimbrial interaction with plasminogen is not based on lectin-like properties and thus can be considered moonlighting by the filamentous structure. In the S-fimbria associated with E. coli strains causing newborn meningitis, genetic studies indicated that the interaction involves a fimbrial minor subunit other than the sialic acid-binding lectin subunit [130]. Curli is a common filamentous adhesin expressed by E. coli and S. enterica and shown to interact with human contact phase proteins [131], which are involved in fever, hypotension and bleeding disorders associated with the infections. These examples show that fimbrial filaments affect bacterial colonization and pathogenicity also via functions other than their adhesiveness to host tissues. Also, flagellar filaments immobilize plasminogen apparently via lysine residues exposed on the filament surface and thus increase proteolytic activity on the cells [132]. Flagella are associated with bacterial mobility but also possess adhesive properties. Flagella of Pseudomonas aeruginosa bind to heparan sulphate proteoglycans at the basolateral surface of airway epithelium [133] and to human Lex and sialyl Lex determinants in human respiratory mucins [134]. Flagella of S. enterica moonlight not only in plasminogen binding [129] but also bind to and enhance biofilm formation on cholesterol-containing surfaces [135]. The latter was concluded to enhance the chronic carrier state of the bacteria on gallstones. Flagella of enteropathogenic (EPEC), enterohemorrhagic (EHEC), meningitic, enterotoxigenic, and probiotic E. coli strains have been identified to mediate bacterial adhesion to epithelial cells [136,137,138,139,140]. These flagella are of different serotypes, and the identified molecular targets include human or animal gut mucins. Interestingly, the EPEC and the EHEC flagella differ in binding to collagen, laminin and fibronectin [137]. Fimbriae and flagella are multimeric protein complexes, and the subunits active in the above activities have not yet been identified, hence it is open whether they represent true moonlighting of a single protein species.
The classical example of a surface protease with moonlighting adhesive functions is the Hap hemagglutinin/protease secreted by Vibrio cholerae [141], which perturbs pericellular barrier function in epithelial cells by degrading tight junctions [142] and by hydrolyzing fibronectin and ovomucin on human epithelial cells [143]. Hap is a metalloproteinase that also degrades lactoferrin and nicks enterotoxin as well as agglutinates chicken erythrocytes and participates in bacterial attachment to gut epithelium. These properties are thought to enhance spread of the cholera bacterium within the human intestine. Pla is a surface protease—a plasminogen activator—and a dramatic virulence factor of the plague bacterium Yersinia pestis that destroys control of the hemostasis balance in humans [144]. Pla increases virulence of Y. pestis also by mechanisms that do not involve proteolysis. It mediates adhesion to laminin of basement membranes [145], which is sensitive to plasmin activity created by e.g., Pla itself. Degradation of laminin is thought to loosen tissue barriers and enhance migration of bacteria at early stages of plague. Another non-proteolytic mechanism of how Pla enhances bacterial dissemination is invasiveness to alveolar macrophages, where Pla utilizes the C-type lectin receptor DEC-205 for invasion [146].
These findings demonstrate that moonlighting, or multifunctionality, in bacteria goes in several directions, metabolic enzymes moonlight as adhesins and as immunomodulators, canonical adhesion proteins moonlight with host circulating effector proteins, and taxis proteins and proteases moonlight as adhesins and/or invasins.
8. Conclusions
It has become evident that moonlighting, or the capacity to perform biological functions not related to the canonical function assigned to the protein, is common in bacterial proteins. Moonlighting was originally considered a virulence trait in pathogenic bacteria, but the same protein species with moonlighting functions have been described in pathogens and in members of the human or the animal microbiota. A general conclusion we wish to stress is that moonlighting proteins appear to be part of bacterial routine in communication with the environment and in responses to environmental changes or stress. The findings that moonlighting proteins released from a single bacterial species can reassociate onto the surface of another species [28,29,68] can be seen as a novel mechanism in bacteria-bacteria interactions and may have importance in bacteria-host interactions.
Our hypothesis is, that the moonlighting proteins in pathogenic species and in commensal lactobacilli are functionally different because they are anchored differently on the bacterial surface. Several pathogenic species seem to retain moonlighting proteins on cell surface at neutral pH [38,39,41,42,45,50,53,60], whereas this is not the case for several lactobacilli [16,28,32,47,92]. The latter is illustrated for L. crispatus in Figure 3. Moonlighting proteins remain cell-bound by ionic interactions and are highly adhesive at low pH, whereas at neutral or basic pH they are detached from the cell surface into the medium, where they may inhibit bacterial adhesion or have other interactions with human cells or tissues. Also, in the case of L. crispatus ST1 described here, the bacterium has other adhesins that are surface-bound by the sortase mechanism [147] and function well at neutral pH [148,149]. The detachment of moonlighting proteins from L. crispatus cells is enhanced by low concentrations of epithelium-derived cationic peptides such as LL-37, which suggests that the anchoring/detachment phenomenon may take place in vivo. Binding and activation of the human plasminogen is an established virulence function in several bacterial infections [99]. Lactobacillus is regarded as a safe organism for humans and a member of the microbiota, but cells of several Lactobacillus species very efficiently enhance activation of plasminogen at neutral pH. However, unlike with several pathogens (reviewed in Bergmann and Hammerschimdt, 2007 [150]; Lähteenmäki and coworkers [99]), the activity at neutral pH is in the cell-free buffer and not protected from α2-antiplasmin [17,28,47]. This means that the plasmin created in the presence of lactobacillar cells remains proteolytically active only when complexed onto a near-by lysine-containing surface, such as the fibrin clot which is the main physiological target for plasmin proteolysis [151,152]. Indeed, a lowered fibrin deposition in the presence of lactobacilli has been observed in mice suffering from pneumococcal lung infection [153,154]. These findings exemplify a mechanism of how bacterial species may differ in the biology of a moonlighting function and stress the importance of our understanding of the mechanisms and the efficiency of surface anchoring of moonlighting proteins.
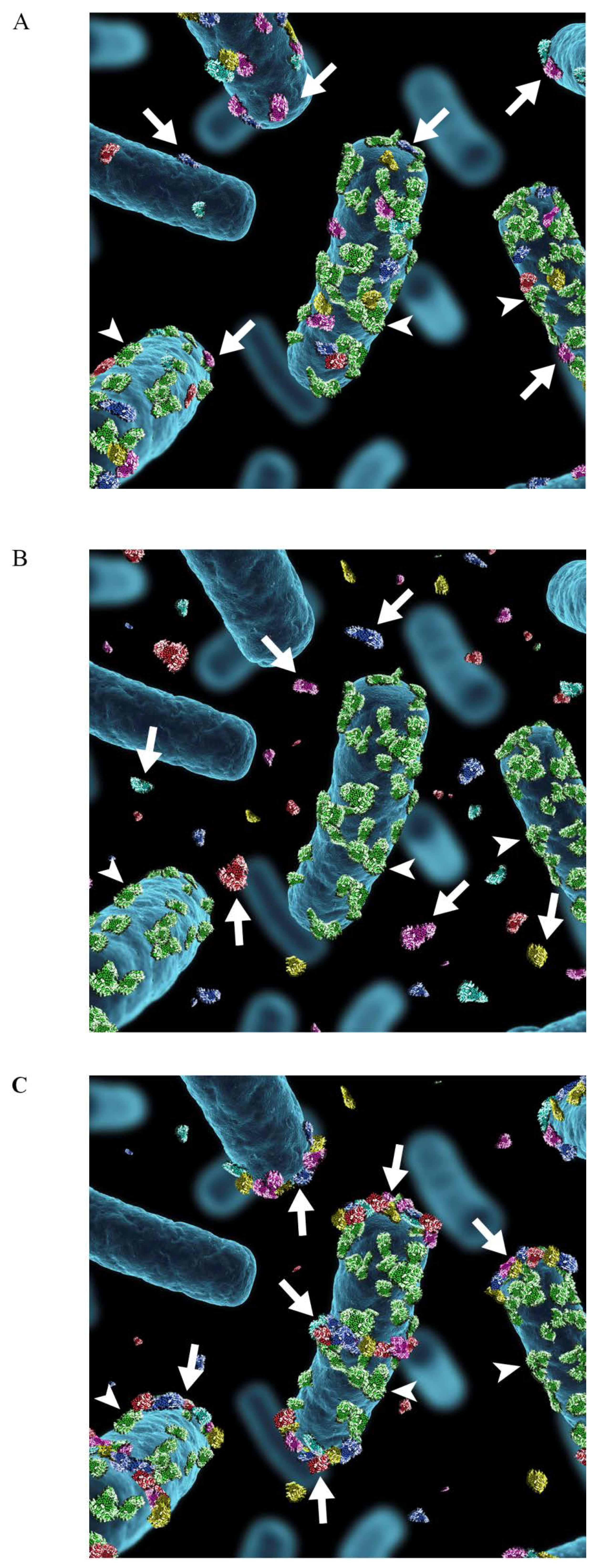
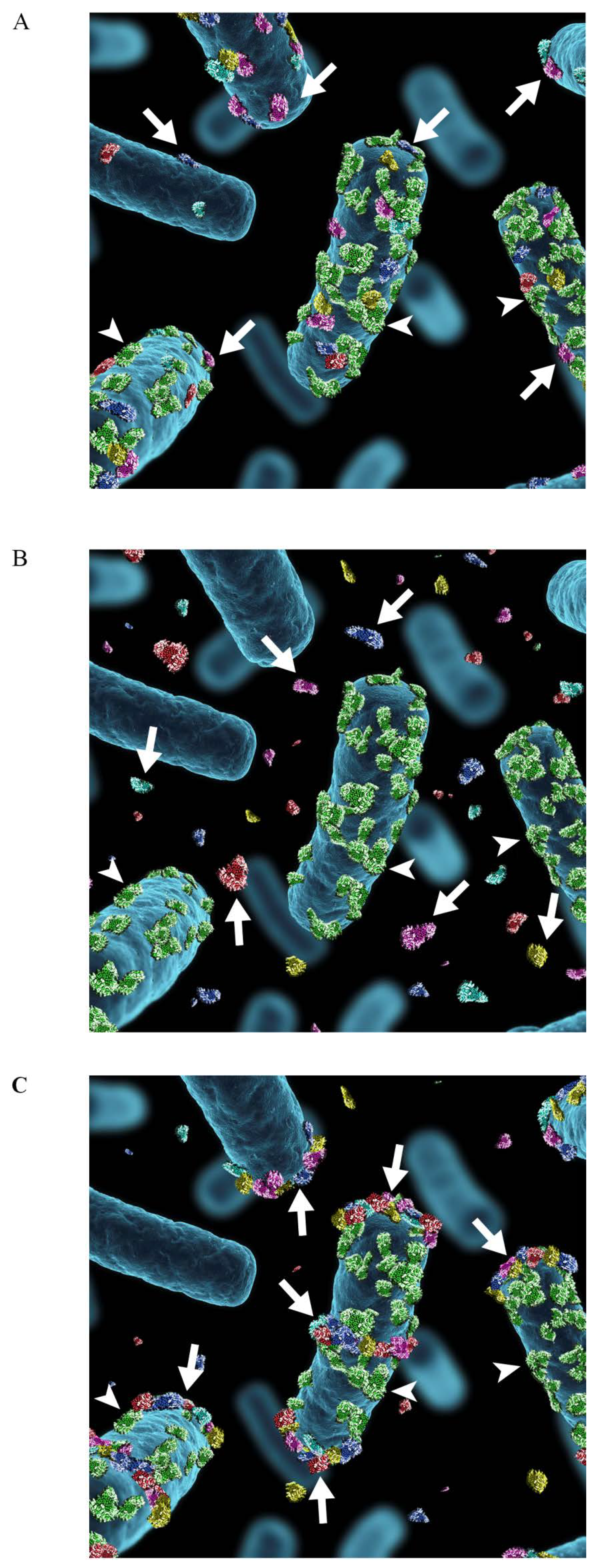
Figure 3.
Schematic illustration of the association of moonlighting proteins with the cell surface of Lactobacillus. (A) Moonlighting proteins (shown in red, blue, yellow, purple and turquoise) of Lactobacillus associate to the cell surface via electrostatic or ionic interactions (B) and they are released into surroundings in stress situations, such as neutral or alkaline pH or presence of cathelidicins or bile acids. The surface location of the S-layer protein (shown in green) with the pI of 10, is not affected by environmental changes. (C) The extracellularly released moonlighting proteins associate back onto the cell surface in favorable environmental conditions, i.e., at acidic pH. The proteins associate with the cell surface of the same lactobacillar species where the proteins were originally released, but also with the cell surface of other lactobacillar species. The binding is not evenly distributed around the cells but is concentrated to cell division areas as well as to the cell poles. The arrows indicate the moonlighting proteins and arrowheads the S-layer protein.
Figure 3.
Schematic illustration of the association of moonlighting proteins with the cell surface of Lactobacillus. (A) Moonlighting proteins (shown in red, blue, yellow, purple and turquoise) of Lactobacillus associate to the cell surface via electrostatic or ionic interactions (B) and they are released into surroundings in stress situations, such as neutral or alkaline pH or presence of cathelidicins or bile acids. The surface location of the S-layer protein (shown in green) with the pI of 10, is not affected by environmental changes. (C) The extracellularly released moonlighting proteins associate back onto the cell surface in favorable environmental conditions, i.e., at acidic pH. The proteins associate with the cell surface of the same lactobacillar species where the proteins were originally released, but also with the cell surface of other lactobacillar species. The binding is not evenly distributed around the cells but is concentrated to cell division areas as well as to the cell poles. The arrows indicate the moonlighting proteins and arrowheads the S-layer protein.
Acknowledgements
The authors thank Pasi Kainulainen for technical assistance in generating Figure 3.
Author Contributions
VK compiled the figures and tables and VK and TKK wrote the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Copley, S.D. Moonlighting is Mainstream: Paradigm Adjustment Required. Bioessays 2012, 34, 578–588. [Google Scholar] [CrossRef]
- Tompa, P.; Szasz, C.; Buday, L. Structural Disorder Throws New Light on Moonlighting. Trends Biochem. Sci. 2005, 30, 484–489. [Google Scholar] [CrossRef]
- Chaput, M.; Claes, V.; Portetelle, D.; Cludts, I.; Cravador, A.; Burny, A.; Gras, H.; Tartar, A. The Neurotrophic Factor Neuroleukin is 90% Homologous with Phosphohexose Isomerase. Nature 1988, 332, 454–455. [Google Scholar] [CrossRef]
- Wistow, G.J.; Lietman, T.; Williams, L.A.; Stapel, S.O.; de Jong, W.W.; Horwitz, J.; Piatigorsky, J. Tau-crystallin/alpha-Enolase: One Gene Encodes both an Enzyme and a Lens Structural Protein. J. Cell Biol. 1988, 107, 2729–2736. [Google Scholar] [CrossRef]
- Dworschack, R.G.; Wickerham, L.J. Production of Extracellular and Total Invertase by Candida Utilis, Saccharomyces Cerevisiae, and Other Yeasts. Appl. Microbiol. 1961, 9, 291–294. [Google Scholar]
- Pancholi, V.; Fischetti, V.A. A Major Surface Protein on Group A Streptococci is a Glyceraldehyde-3-Phosphate-Dehydrogenase with Multiple Binding Activity. J. Exp. Med. 1992, 176, 415–426. [Google Scholar] [CrossRef]
- Commichau, F.M.; Rothe, F.M.; Herzberg, C.; Wagner, E.; Hellwig, D.; Lehnik-Habrink, M.; Hammer, E.; Volker, U.; Stulke, J. Novel Activities of Glycolytic Enzymes in Bacillus subtilis: Interactions with Essential Proteins Involved in mRNA Processing. Mol. Cell. Proteomics 2009, 8, 1350–1360. [Google Scholar] [CrossRef]
- Huberts, D.H.; van der Klei, I.J. Moonlighting Proteins: An Intriguing Mode of Multitasking. Biochim. Biophys. Acta 2010, 1803, 520–525. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting Proteins—An Update. Mol. Biosyst. 2009, 5, 345–350. [Google Scholar] [CrossRef]
- Henderson, B.; Martin, A. Bacterial Virulence in the Moonlight: Multitasking Bacterial Moonlighting Proteins are Virulence Determinants in Infectious Disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef]
- Gomez, A.; Hernandez, S.; Amela, I.; Pinol, J.; Cedano, J.; Querol, E. Do Protein-Protein Interaction Databases Identify Moonlighting Proteins? Mol. Biosyst. 2011, 7, 2379–2382. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Chitale, M.; Rayon, C.; Kihara, D. Evaluation of Function Predictions by PFP, ESG, and PSI-BLAST for Moonlighting Proteins. BMC Proc. 2012, 6. [Google Scholar] [CrossRef]
- Bergmann, S.; Rohde, M.; Chhatwal, G.S.; Hammerschmidt, S. Characterization of Plasmin(Ogen) Binding to Streptococcus pneumoniae. Indian J. Med. Res. 2004, 119, 29–32. [Google Scholar]
- Egea, L.; Aguilera, L.; Gimenez, R.; Sorolla, M.A.; Aguilar, J.; Badia, J.; Baldoma, L. Role of Secreted Glyceraldehyde-3-Phosphate Dehydrogenase in the Infection Mechanism of Enterohemorrhagic and Enteropathogenic Escherichia coli: Interaction of the Extracellular Enzyme with Human Plasminogen and Fibrinogen. Int. J. Biochem. Cell Biol. 2007, 39, 1190–1203. [Google Scholar] [CrossRef]
- Gase, K.; Gase, A.; Schirmer, H.; Malke, H. Cloning, Sequencing and Functional Overexpression of the Streptococcus equisimilis H46A gapC Gene Encoding a Glyceraldehyde-3-Phosphate Dehydrogenase that also Functions as a Plasmin(Ogen)-Binding Protein. Purification and Biochemical Characterization of the Protein. Eur. J. Biochem. 1996, 239, 42–51. [Google Scholar]
- Glenting, J.; Beck, H.C.; Vrang, A.; Riemann, H.; Ravn, P.; Hansen, A.M.; Antonsson, M.; Ahrne, S.; Israelsen, H.; Madsen, S. Anchorless Surface Associated Glycolytic Enzymes from Lactobacillus plantarum 299v Bind to Epithelial Cells and Extracellular Matrix Proteins. Microbiol. Res. 2013, 168, 245–253. [Google Scholar] [CrossRef]
- Hurmalainen, V.; Edelman, S.; Antikainen, J.; Baumann, M.; Lähteenmäki, K.; Korhonen, T.K. Extracellular Proteins of Lactobacillus crispatus enhance Activation of Human Plasminogen. Microbiology 2007, 153, 1112–1122. [Google Scholar] [CrossRef]
- Jobin, M.C.; Brassard, J.; Quessy, S.; Gottschalk, M.; Grenier, D. Acquisition of Host Plasmin Activity by the Swine Pathogen Streptococcus suis Serotype 2. Infect. Immun. 2004, 72, 606–610. [Google Scholar] [CrossRef]
- Kinnby, B.; Booth, N.A.; Svensater, G. Plasminogen Binding by Oral Streptococci from Dental Plaque and Inflammatory Lesions. Microbiology 2008, 154, 924–931. [Google Scholar] [CrossRef]
- Modun, B.; Williams, P. The Staphylococcal Transferrin-Binding Protein is a Cell Wall Glyceraldehyde-3-Phosphate Dehydrogenase. Infect. Immun. 1999, 67, 1086–1092. [Google Scholar]
- Schaumburg, J.; Diekmann, O.; Hagendorff, P.; Bergmann, S.; Rohde, M.; Hammerschmidt, S.; Jänsch, L.; Wehland, J.; Kärst, U. The Cell Wall Subproteome of Listeria monocytogenes. Proteomics 2004, 4, 2991–3006. [Google Scholar] [CrossRef]
- Seifert, K.N.; McArthur, W.P.; Bleiweis, A.S.; Brady, L.J. Characterization of Group B Streptococcal Glyceraldehyde-3-Phosphate Dehydrogenase: Surface Localization, Enzymatic Activity, and Protein-Protein Interactions. Can. J. Microbiol. 2003, 49, 350–356. [Google Scholar] [CrossRef]
- Winram, S.B.; Lottenberg, R. The Plasmin-Binding Protein Plr of Group A Streptococci is Identified as Glyceraldehyde-3-Phosphate Dehydrogenase. Microbiology 1996, 142, 2311–2320. [Google Scholar] [CrossRef]
- Matta, S.K.; Agarwal, S.; Bhatnagar, R. Surface Localized and Extracellular Glyceraldehyde-3-Phosphate Dehydrogenase of Bacillus anthracis is a Plasminogen Binding Protein. Biochim. Biophys. Acta 2010, 1804, 2111–2120. [Google Scholar] [CrossRef]
- Jin, H.; Song, Y.P.; Boel, G.; Kochar, J.; Pancholi, V. Group A Streptococcal Surface GAPDH, SDH, Recognizes uPAR/CD87 as its Receptor on the Human Pharyngeal Cell and Mediates Bacterial Adherence to Host Cells. J. Mol. Biol. 2005, 350, 27–41. [Google Scholar] [CrossRef]
- Quessy, S.; Busque, P.; Higgins, R.; Jacques, M.; Dubreuil, J.D. Description of an Albumin Binding Activity for Streptococcus suis Serotype 2. FEMS Microbiol. Lett. 1997, 147, 245–250. [Google Scholar] [CrossRef]
- Sanchez, B.; Schmitter, J.M.; Urdaci, M.C. Identification of Novel Proteins Secreted by Lactobacillus plantarum that Bind to Mucin and Fibronectin. J. Mol. Microbiol. Biotechnol. 2009, 17, 158–162. [Google Scholar] [CrossRef]
- Kainulainen, V.; Loimaranta, V.; Pekkala, A.; Edelman, S.; Antikainen, J.; Kylväjä, R.; Laaksonen, M.; Laakkonen, L.; Finne, J.; Korhonen, T.K. Glutamine Synthetase and Glucose-6-Phosphate Isomerase are Adhesive Moonlighting Proteins of Lactobacillus crispatus Released by Epithelial Cathelicidin LL-37. J. Bacteriol. 2012, 194, 2509–2519. [Google Scholar] [CrossRef]
- Oliveira, L.; Madureira, P.; Andrade, E.B.; Bouaboud, A.; Morello, E.; Ferreira, P.; Poyart, C.; Trieu-Cuot, P.; Dramsi, S. Group B Streptococcus GAPDH is Released upon Cell Lysis, Associates with Bacterial Surface, and Induces Apoptosis in Murine Macrophages. PLoS One 2012, 7, e29963. [Google Scholar] [CrossRef] [Green Version]
- Ramiah, K.; van Reenen, C.A.; Dicks, L.M. Surface-Bound Proteins of Lactobacillus plantarum 423 that Contribute to Adhesion of Caco-2 Cells and their Role in Competitive Exclusion and Displacement of Clostridium Sporogenes and Enterococcus Faecalis. Res. Microbiol. 2008, 159, 470–475. [Google Scholar] [CrossRef]
- Maeda, K.; Nagata, H.; Yamamoto, Y.; Tanaka, M.; Tanaka, J.; Minamino, N.; Shizukuishi, S. Glyceraldehyde-3-Phosphate Dehydrogenase of Streptococcus oralis Functions as a Coadhesin for Porphyromonas gingivalis Major Fimbriae. Infect. Immun. 2004, 72, 1341–1348. [Google Scholar] [CrossRef]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kawasaki, T.; Wakahara, N.; Matsuo, H.; Watanabe, M.; Kitazawa, H.; Ohnuma, S.; Miura, K.; et al. Cell Surface Lactobacillus plantarum LA 318 Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) Adheres to Human Colonic Mucin. J. Appl. Microbiol. 2008, 104, 1667–1674. [Google Scholar] [CrossRef]
- Alvarez, R.A.; Blaylock, M.W.; Baseman, J.B. Surface Localized Glyceraldehyde-3-Phosphate Dehydrogenase of Mycoplasma genitalium Binds Mucin. Mol. Microbiol. 2003, 48, 1417–1425. [Google Scholar] [CrossRef]
- Kinoshita, H.; Wakahara, N.; Watanabe, M.; Kawasaki, T.; Matsuo, H.; Kawai, Y.; Kitazawa, H.; Ohnuma, S.; Miura, K.; Horii, A.; et al. Cell Surface Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) of Lactobacillus plantarum LA 318 Recognizes Human A and B Blood Group Antigens. Res. Microbiol. 2008, 159, 685–691. [Google Scholar] [CrossRef]
- Brassard, J.; Gottschalk, M.; Quessy, S. Cloning and Purification of the Streptococcus suis Serotype 2 Glyceraldehyde-3-Phosphate Dehydrogenase and its Involvement as an Adhesin. Vet. Microbiol. 2004, 102, 87–94. [Google Scholar] [CrossRef]
- Bermudez, L.E.; Petrofsky, M.; Shelton, K. Epidermal Growth Factor-Binding Protein in Mycobacterium avium and Mycobacterium tuberculosis: A Possible Role in the Mechanism of Infection. Infect. Immun. 1996, 64, 2917–2922. [Google Scholar]
- Terao, Y.; Yamaguchi, M.; Hamada, S.; Kawabata, S. Multifunctional Glyceraldehyde-3-Phosphate Dehydrogenase of Streptococcus pyogenes is Essential for Evasion from Neutrophils. J. Biol. Chem. 2006, 281, 14215–14223. [Google Scholar] [CrossRef]
- Agarwal, S.; Kulshreshtha, P.; Bambah Mukku, D.; Bhatnagar, R. Alpha-Enolase Binds to Human Plasminogen on the Surface of Bacillus anthracis. Biochim. Biophys. Acta 2008, 1784, 986–994. [Google Scholar] [CrossRef]
- Bergmann, S.; Rohde, M.; Chhatwal, G.S.; Hammerschmidt, S. Alpha-Enolase of Streptococcus pneumoniae is a Plasmin(Ogen)-Binding Protein Displayed on the Bacterial Cell Surface. Mol. Microbiol. 2001, 40, 1273–1287. [Google Scholar] [CrossRef]
- Candela, M.; Biagi, E.; Centanni, M.; Turroni, S.; Vici, M.; Musiani, F.; Vitali, B.; Bergmann, S.; Hammerschmidt, S.; Brigidi, P. Bifidobacterial Enolase, a Cell Surface Receptor for Human Plasminogen Involved in the Interaction with the Host. Microbiology 2009, 155, 3294–3303. [Google Scholar] [CrossRef]
- Floden, A.M.; Watt, J.A.; Brissette, C.A. Borrelia burgdorferi Enolase is a Surface-Exposed Plasminogen Binding Protein. PLoS One 2011, 6, e27502. [Google Scholar] [CrossRef]
- Ge, J.; Catt, D.M.; Gregory, R.L. Streptococcus mutans Surface Alpha-Enolase Binds Salivary Mucin MG2 and Human Plasminogen. Infect. Immun. 2004, 72, 6748–6752. [Google Scholar] [CrossRef]
- Itzek, A.; Gillen, C.M.; Fulde, M.; Friedrichs, C.; Rodloff, A.C.; Chhatwal, G.S.; Nitsche-Schmitz, D.P. Contribution of Plasminogen Activation Towards the Pathogenic Potential of Oral Streptococci. PLoS One 2010, 5, e13826. [Google Scholar] [CrossRef]
- Mölkänen, T.; Tyynelä, J.; Helin, J.; Kalkkinen, N.; Kuusela, P. Enhanced Activation of Bound Plasminogen on Staphylococcus aureus by Staphylokinase. FEBS Lett. 2002, 517, 72–78. [Google Scholar] [CrossRef]
- Pancholi, V.; Fischetti, V.A. Alpha-Enolase, a Novel Strong Plasmin(Ogen) Binding Protein on the Surface of Pathogenic Streptococci. J. Biol. Chem. 1998, 273, 14503–14515. [Google Scholar] [CrossRef]
- Thomas, C.; Jacobs, E.; Dumke, R. Characterization of Pyruvate Dehydrogenase Subunit B and Enolase as Plasminogen-Binding Proteins in Mycoplasma Pneumoniae. Microbiology 2013, 159, 352–365. [Google Scholar] [CrossRef]
- Vastano, V.; Capri, U.; Candela, M.; Siciliano, R.A.; Russo, L.; Renda, M.; Sacco, M. Identification of Binding Sites of Lactobacillus plantarum Enolase Involved in the Interaction with Human Plasminogen. Microbiol. Res. 2013, 168, 65–72. [Google Scholar] [CrossRef]
- Antikainen, J.; Kuparinen, V.; Lähteenmäki, K.; Korhonen, T.K. Enolases from Gram-Positive Bacterial Pathogens and Commensal Lactobacilli Share Functional Similarity in Virulence-Associated Traits. FEMS Immunol. Med. Microbiol. 2007, 51, 526–534. [Google Scholar] [CrossRef]
- Castaldo, C.; Vastano, V.; Siciliano, R.A.; Candela, M.; Vici, M.; Muscariello, L.; Marasco, R.; Sacco, M. Surface Displaced Alfa-Enolase of Lactobacillus plantarum is a Fibronectin Binding Protein. Microb. Cell. Fact. 2009, 8, 14. [Google Scholar] [CrossRef]
- Carneiro, C.R.; Postol, E.; Nomizo, R.; Reis, L.F.; Brentani, R.R. Identification of Enolase as a Laminin-Binding Protein on the Surface of Staphylococcus aureus. Microbes Infect. 2004, 6, 604–608. [Google Scholar] [CrossRef]
- Kornblatt, M.J.; Kornblatt, J.A.; Hancock, M.A. The Interaction of Canine Plasminogen with Streptococcus Pyogenes Enolase: They Bind to One another but what is the Nature of the Structures Involved? PLoS One 2011, 6, e28481. [Google Scholar]
- Kesimer, M.; Kilic, N.; Mehrotra, R.; Thornton, D.J.; Sheehan, J.K. Identification of Salivary Mucin MUC7 Binding Proteins from Streptococcus gordonii. BMC Microbiol. 2009, 9, 163. [Google Scholar] [CrossRef]
- Feng, Y.; Pan, X.; Sun, W.; Wang, C.; Zhang, H.; Li, X.; Ma, Y.; Shao, Z.; Ge, J.; Zheng, F.; et al. Streptococcus suis Enolase Functions as a Protective Antigen Displayed on the Bacterial Cell Surface. J. Infect. Dis. 2009, 200, 1583–1592. [Google Scholar] [CrossRef]
- Agarwal, V.; Hammerschmidt, S.; Malm, S.; Bergmann, S.; Riesbeck, K.; Blom, A.M. Enolase of Streptococcus pneumoniae Binds Human Complement Inhibitor C4b-Binding Protein and Contributes to Complement Evasion. J. Immunol. 2012, 189, 3575–3584. [Google Scholar] [CrossRef]
- Blau, K.; Portnoi, M.; Shagan, M.; Kaganovich, A.; Rom, S.; Kafka, D.; Chalifa Caspi, V.; Porgador, A.; Givon-Lavi, N.; Gershoni, J.M.; et al. Flamingo Cadherin: A Putative Host Receptor for Streptococcus pneumoniae. J. Infect. Dis. 2007, 195, 1828–1837. [Google Scholar] [CrossRef]
- Boone, T.J.; Burnham, C.A.; Tyrrell, G.J. Binding of Group B Streptococcal Phosphoglycerate Kinase to Plasminogen and Actin. Microb. Pathog. 2011, 51, 255–261. [Google Scholar] [CrossRef]
- Burnham, C.A.; Shokoples, S.E.; Tyrrell, G.J. Phosphoglycerate Kinase Inhibits Epithelial Cell Invasion by Group B Streptococci. Microb. Pathog. 2005, 38, 189–200. [Google Scholar] [CrossRef]
- Candela, M.; Bergmann, S.; Vici, M.; Vitali, B.; Turroni, S.; Eikmanns, B.J.; Hammerschmidt, S.; Brigidi, P. Binding of Human Plasminogen to Bifidobacterium. J. Bacteriol. 2007, 189, 5929–5936. [Google Scholar] [CrossRef]
- Xolalpa, W.; Vallecillo, A.J.; Lara, M.; Mendoza-Hernandez, G.; Comini, M.; Spallek, R.; Singh, M.; Espitia, C. Identification of Novel Bacterial Plasminogen-Binding Proteins in the Human Pathogen Mycobacterium tuberculosis. Proteomics 2007, 7, 3332–3341. [Google Scholar] [CrossRef]
- Jagadeesan, B.; Koo, O.K.; Kim, K.P.; Burkholder, K.M.; Mishra, K.K.; Aroonnual, A.; Bhunia, A.K. LAP, an Alcohol Acetaldehyde Dehydrogenase Enzyme in Listeria, Promotes Bacterial Adhesion to Enterocyte-Like Caco-2 Cells Only in Pathogenic Species. Microbiology 2010, 156, 2782–2795. [Google Scholar] [CrossRef]
- Kim, K.P.; Jagadeesan, B.; Burkholder, K.M.; Jaradat, Z.W.; Wampler, J.L.; Lathrop, A.A.; Morgan, M.T.; Bhunia, A.K. Adhesion Characteristics of Listeria Adhesion Protein (LAP)-Expressing Escherichia coli to Caco-2 Cells and of Recombinant LAP to Eukaryotic Receptor Hsp60 as Examined in a Surface Plasmon Resonance Sensor. FEMS Microbiol. Lett. 2006, 256, 324–332. [Google Scholar] [CrossRef]
- Kinhikar, A.G.; Vargas, D.; Li, H.; Mahaffey, S.B.; Hinds, L.; Belisle, J.T.; Laal, S. Mycobacterium tuberculosis Malate Synthase is a Laminin-Binding Adhesin. Mol. Microbiol. 2006, 60, 999–1013. [Google Scholar] [CrossRef]
- Dallo, S.F.; Kannan, T.R.; Blaylock, M.W.; Baseman, J.B. Elongation Factor Tu and E1 Beta Subunit of Pyruvate Dehydrogenase Complex Act as Fibronectin Binding Proteins in Mycoplasma pneumoniae. Mol. Microbiol. 2002, 46, 1041–1051. [Google Scholar] [CrossRef]
- Candela, M.; Centanni, M.; Fiori, J.; Biagi, E.; Turroni, S.; Orrico, C.; Bergmann, S.; Hammerschmidt, S.; Brigidi, P. DnaK from Bifidobacterium animalis Subsp. lactis is a Surface-Exposed Human Plasminogen Receptor Upregulated in Response to Bile Salts. Microbiology 2010, 156, 1609–1618. [Google Scholar] [CrossRef]
- Floto, R.A.; MacAry, P.A.; Boname, J.M.; Mien, T.S.; Kampmann, B.; Hair, J.R.; Huey, O.S.; Houben, E.N.; Pieters, J.; Day, C.; et al. Dendritic Cell Stimulation by Mycobacterial Hsp70 is Mediated through CCR5. Science 2006, 314, 454–458. [Google Scholar] [CrossRef]
- Babaahmady, K.; Oehlmann, W.; Singh, M.; Lehner, T. Inhibition of Human Immunodeficiency Virus Type 1 Infection of Human CD4+ T Cells by Microbial HSP70 and the Peptide Epitope 407–426. J. Virol. 2007, 81, 3354–3360. [Google Scholar] [CrossRef]
- Katakura, Y.; Sano, R.; Hashimoto, T.; Ninomiya, K.; Shioya, S. Lactic Acid Bacteria Display on the Cell Surface Cytosolic Proteins that Recognize Yeast Mannan. Appl. Microbiol. Biotechnol. 2010, 86, 319–326. [Google Scholar] [CrossRef]
- Bergonzelli, G.E.; Granato, D.; Pridmore, R.D.; Marvin-Guy, L.F.; Donnicola, D.; Corthésy-Theulaz, I.E. GroEL of Lactobacillus johnsonii La1 (NCC 533) is Cell Surface Associated: Potential Role in Interactions with the Host and the Gastric Pathogen Helicobacter pylori. Infect. Immun. 2006, 74, 425–434. [Google Scholar] [CrossRef]
- Kunert, A.; Losse, J.; Gruszin, C.; Huhn, M.; Kaendler, K.; Mikkat, S.; Volke, D.; Hoffmann, R.; Jokiranta, T.S.; Seeberger, H.; et al. Immune Evasion of the Human Pathogen Pseudomonas aeruginosa: Elongation Factor Tuf is a Factor H and Plasminogen Binding Protein. J. Immunol. 2007, 179, 2979–2988. [Google Scholar]
- Granato, D.; Bergonzelli, G.E.; Pridmore, R.D.; Marvin, L.; Rouvet, M.; Corthésy-Theulaz, I.E. Cell Surface-Associated Elongation Factor Tu Mediates the Attachment of Lactobacillus johnsonii NCC533 (La1) to Human Intestinal Cells and Mucins. Infect. Immun. 2004, 72, 2160–2169. [Google Scholar] [CrossRef]
- Agarwal, V.; Kuchipudi, A.; Fulde, M.; Riesbeck, K.; Bergmann, S.; Blom, A.M. Streptococcus pneumoniae Endopeptidase O (PepO) is a Multifunctional Plasminogen- and Fibronectin-Binding Protein, Facilitating Evasion of Innate Immunity and Invasion of Host Cells. J. Biol. Chem. 2013, 288, 6849–6863. [Google Scholar] [CrossRef]
- Reddy, V.M.; Suleman, F.G. Mycobacterium avium-Superoxide Dismutase Binds to Epithelial Cell Aldolase, Glyceraldehyde-3-Phosphate Dehydrogenase and Cyclophilin A. Microb. Pathog. 2004, 36, 67–74. [Google Scholar] [CrossRef]
- Jeffery, C.J. Mass Spectrometry and the Search for Moonlighting Proteins. Mass Spectrom. Rev. 2005, 24, 772–782. [Google Scholar] [CrossRef]
- Bergmann, S.; Wild, D.; Diekmann, O.; Frank, R.; Bracht, D.; Chhatwal, G.S.; Hammerschmidt, S. Identification of a Novel Plasmin(Ogen)-Binding Motif in Surface Displayed Alpha-Enolase of Streptococcus pneumoniae. Mol. Microbiol. 2003, 49, 411–423. [Google Scholar] [CrossRef]
- Cork, A.J.; Jergic, S.; Hammerschmidt, S.; Kobe, B.; Pancholi, V.; Benesch, J.L.; Robinson, C.V.; Dixon, N.E.; Aquilina, J.A.; Walker, M.J. Defining the Structural Basis of Human Plasminogen Binding by Streptococcal Surface Enolase. J. Biol. Chem. 2009, 284, 17129–17137. [Google Scholar] [CrossRef]
- Ehinger, S.; Schubert, W.D.; Bergmann, S.; Hammerschmidt, S.; Heinz, D.W. Plasmin(Ogen)-Binding Alpha-Enolase from Streptococcus pneumoniae: Crystal Structure and Evaluation of Plasmin(Ogen)-Binding Sites. J. Mol. Biol. 2004, 343, 997–1005. [Google Scholar] [CrossRef]
- Pridmore, R.D.; Berger, B.; Desiere, F.; Vilanova, D.; Barretto, C.; Pittet, A.C.; Zwahlen, M.C.; Rouvet, M.; Altermann, E.; Barrangou, R.; et al. The Genome Sequence of the Probiotic Intestinal Bacterium Lactobacillus johnsonii NCC 533. Proc. Natl. Acad. Sci. USA 2004, 101, 2512–2517. [Google Scholar]
- Siezen, R.J.; Francke, C.; Renckens, B.; Boekhorst, J.; Wels, M.; Kleerebezem, M.; van Hijum, S.A. Complete Resequencing and Reannotation of the Lactobacillus plantarum WCFS1 Genome. J. Bacteriol. 2012, 194, 195–196. [Google Scholar] [CrossRef]
- Lewthwaite, J.; Skinner, A.; Henderson, B. Are Molecular Chaperones Microbial Virulence Factors? Trends Microbiol. 1998, 6, 426–428. [Google Scholar] [CrossRef]
- Hu, Y.; Henderson, B.; Lund, P.A.; Tormay, P.; Ahmed, M.T.; Gurcha, S.S.; Besra, G.S.; Coates, A.R. A Mycobacterium tuberculosis Mutant Lacking the groEL Homologue cpn60.1 is Viable but Fails to Induce an Inflammatory Response in Animal Models of Infection. Infect. Immun. 2008, 76, 1535–1546. [Google Scholar] [CrossRef]
- Cehovin, A.; Coates, A.R.; Hu, Y.; Riffo-Vasquez, Y.; Tormay, P.; Botanch, C.; Altare, F.; Henderson, B. Comparison of the Moonlighting Actions of the Two Highly Homologous Chaperonin 60 Proteins of Mycobacterium tuberculosis. Infect. Immun. 2010, 78, 3196–3206. [Google Scholar] [CrossRef]
- Hickey, T.B.; Thorson, L.M.; Speert, D.P.; Daffe, M.; Stokes, R.W. Mycobacterium tuberculosis Cpn60.2 and DnaK are Located on the Bacterial Surface, Where Cpn60.2 Facilitates Efficient Bacterial Association with Macrophages. Infect. Immun. 2009, 77, 3389–3401. [Google Scholar] [CrossRef]
- Hickey, T.B.; Ziltener, H.J.; Speert, D.P.; Stokes, R.W. Mycobacterium tuberculosis Employs Cpn60.2 as an Adhesin that Binds CD43 on the Macrophage Surface. Cell. Microbiol. 2010, 12, 1634–1647. [Google Scholar] [CrossRef]
- Lewthwaite, J.C.; Coates, A.R.; Tormay, P.; Singh, M.; Mascagni, P.; Poole, S.; Roberts, M.; Sharp, L.; Henderson, B. Mycobacterium tuberculosis Chaperonin 60.1 is a More Potent Cytokine Stimulator than Chaperonin 60.2 (Hsp 65) and Contains a CD14-Binding Domain. Infect. Immun. 2001, 69, 7349–7355. [Google Scholar] [CrossRef]
- Pancholi, V.; Chhatwal, G.S. Housekeeping Enzymes as Virulence Factors for Pathogens. Int. J. Med. Microbiol. 2003, 293, 391–401. [Google Scholar] [CrossRef]
- Boël, G.; Jin, H.; Pancholi, V. Inhibition of Cell Surface Export of Group A Streptococcal Anchorless Surface Dehydrogenase Affects Bacterial Adherence and Antiphagocytic Properties. Infect. Immun. 2005, 73, 6237–6248. [Google Scholar] [CrossRef]
- Boël, G.; Pichereau, V.; Mijakovic, I.; Mazé, A.; Poncet, S.; Gillet, S.; Giard, J.C.; Hartke, A.; Auffray, Y.; Deutscher, J. Is 2-Phosphoglycerate-Dependent Automodification of Bacterial Enolases Implicated in their Export? J. Mol. Biol. 2004, 337, 485–496. [Google Scholar] [CrossRef]
- Yang, C.K.; Ewis, H.E.; Zhang, X.; Lu, C.D.; Hu, H.J.; Pan, Y.; Abdelal, A.T.; Tai, P.C. Nonclassical Protein Secretion by Bacillus subtilis in the Stationary Phase is Not due to Cell Lysis. J. Bacteriol. 2011, 193, 5607–5615. [Google Scholar] [CrossRef]
- Lenz, L.L.; Mohammadi, S.; Geissler, A.; Portnoy, D.A. SecA2-Dependent Secretion of Autolytic Enzymes Promotes Listeria monocytogenes Pathogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 12432–12437. [Google Scholar] [CrossRef]
- Stephenson, K.; Bron, S.; Harwood, C.R. Cellular Lysis in Bacillus subtilis; the Affect of Multiple Extracellular Protease Deficiencies. Lett. Appl. Microbiol. 1999, 29, 141–145. [Google Scholar] [CrossRef]
- Saad, N.; Urdaci, M.; Vignoles, C.; Chaignepain, S.; Tallon, R.; Schmitter, J.M.; Bressollier, P. Lactobacillus plantarum 299v Surface-Bound GAPDH: A New Insight into Enzyme Cell Walls Location. J. Microbiol. Biotechnol. 2009, 19, 1635–1643. [Google Scholar] [CrossRef]
- Antikainen, J.; Hurmalainen, V.; Lähteenmäki, K.; Korhonen, T.K. PH-Dependent Association of Enolase and GAPDH of Lactobacillus crispatus with the Cell Wall and Lipoteichoic Acids. J. Bacteriol. 2007, 189, 4539–4543. [Google Scholar] [CrossRef]
- Baugher, J.L.; Klaenhammer, T.R. Invited Review: Application of Omics Tools to Understanding Probiotic Functionality. J. Dairy Sci. 2011, 94, 4753–4765. [Google Scholar] [CrossRef]
- Corr, S.C.; Hill, C.; Gahan, C.G. Understanding the Mechanisms by which Probiotics Inhibit Gastrointestinal Pathogens. Adv. Food Nutr. Res. 2009, 56, 1–15. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic Activities of Lactobacilli and Bifidobacteria Against Microbial Pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef]
- Heinemann, C.; van Hylckama Vlieg, J.E.; Janssen, D.B.; Busscher, H.J.; van der Mei, H.C.; Reid, G. Purification and Characterization of a Surface-Binding Protein from Lactobacillus fermentum RC-14 that Inhibits Adhesion of Enterococcus faecalis 1131. FEMS Microbiol. Lett. 2000, 190, 177–180. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Arvidson, C.G. Lactobacillus jensenii Surface-Associated Proteins Inhibit Neisseria gonorrhoeae Adherence to Epithelial Cells. Infect. Immun. 2010, 78, 3103–3111. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Ploplis, V.A.; Castellino, F.J. Bacterial Plasminogen Receptors Utilize Host Plasminogen System for Effective Invasion and Dissemination. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Lähteenmäki, K.; Edelman, S.; Korhonen, T.K. Bacterial Metastasis: The Host Plasminogen System in Bacterial Invasion. Trends Microbiol. 2005, 13, 79–85. [Google Scholar] [CrossRef]
- Cesarman-Maus, G.; Hajjar, K.A. Molecular Mechanisms of Fibrinolysis. Br. J. Haematol. 2005, 129, 307–321. [Google Scholar] [CrossRef]
- Sanderson-Smith, M.L.; de Oliveira, D.M.; Ranson, M.; McArthur, J.D. Bacterial Plasminogen Receptors: Mediators of a Multifaceted Relationship. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Siemens, N.; Patenge, N.; Otto, J.; Fiedler, T.; Kreikemeyer, B. Streptococcus pyogenes M49 plasminogen/plasmin Binding Facilitates Keratinocyte Invasion Via Integrin-Integrin-Linked Kinase (ILK) Pathways and Protects from Macrophage Killing. J. Biol. Chem. 2011, 286, 21612–21622. [Google Scholar] [CrossRef]
- Pluskota, E.; Soloviev, D.A.; Bdeir, K.; Cines, D.B.; Plow, E.F. Integrin alphaMbeta2 Orchestrates and Accelerates Plasminogen Activation and Fibrinolysis by Neutrophils. J. Biol. Chem. 2004, 279, 18063–18072. [Google Scholar] [CrossRef]
- Attali, C.; Durmort, C.; Vernet, T.; di Guilmi, A.M. The Interaction of Streptococcus pneumoniae with Plasmin Mediates Transmigration Across Endothelial and Epithelial Monolayers by Intercellular Junction Cleavage. Infect. Immun. 2008, 76, 5350–5356. [Google Scholar] [CrossRef]
- Bergmann, S.; Schoenen, H.; Hammerschmidt, S. The Interaction between Bacterial Enolase and Plasminogen Promotes Adherence of Streptococcus pneumoniae to Epithelial and Endothelial Cells. Int. J. Med. Microbiol. 2013, 303, 452–462. [Google Scholar] [CrossRef]
- Jin, H.; Agarwal, S.; Agarwal, S.; Pancholi, V. Surface Export of GAPDH/SDH, a Glycolytic Enzyme, is Essential for Streptococcus pyogenes Virulence. MBio 2011. [Google Scholar] [CrossRef]
- Pancholi, V.; Fischetti, V.A. Regulation of the Phosphorylation of Human Pharyngeal Cell Proteins by Group A Streptococcal Surface Dehydrogenase: Signal Transduction between Streptococci and Pharyngeal Cells. J. Exp. Med. 1997, 186, 1633–1643. [Google Scholar] [CrossRef]
- Madureira, P.; Baptista, M.; Vieira, M.; Magalhães, V.; Camelo, A.; Oliveira, L.; Ribeiro, A.; Tavares, D.; Trieu-Cuot, P.; Vilanova, M.; et al. Streptococcus agalactiae GAPDH is a Virulence-Associated Immunomodulatory Protein. J. Immunol. 2007, 178, 1379–1387. [Google Scholar]
- Ling, E.; Feldman, G.; Portnoi, M.; Dagan, R.; Overweg, K.; Mulholland, F.; Chalifa-Caspi, V.; Wells, J.; Mizrachi-Nebenzahl, Y. Glycolytic Enzymes Associated with the Cell Surface of Streptococcus pneumoniae are Antigenic in Humans and Elicit Protective Immune Responses in the Mouse. Clin. Exp. Immunol. 2004, 138, 290–298. [Google Scholar] [CrossRef]
- Eberhard, T.; Kronvall, G.; Ullberg, M. Surface Bound Plasmin Promotes Migration of Streptococcus pneumoniae through Reconstituted Basement Membranes. Microb. Pathog. 1999, 26, 175–181. [Google Scholar] [CrossRef]
- Bergmann, S.; Rohde, M.; Preissner, K.T.; Hammerschmidt, S. The Nine Residue Plasminogen-Binding Motif of the Pneumococcal Enolase is the Major Cofactor of Plasmin-Mediated Degradation of Extracellular Matrix, Dissolution of Fibrin and Transmigration. Thromb. Haemost. 2005, 94, 304–311. [Google Scholar]
- Dinis, M.; Tavares, D.; Veiga-Malta, I.; Fonseca, A.J.; Andrade, E.B.; Trigo, G.; Ribeiro, A.; Videira, A.; Cabrita, A.M.; Ferreira, P. Oral Therapeutic Vaccination with Streptococcus sobrinus Recombinant Enolase Confers Protection Against Dental Caries in Rats. J. Infect. Dis. 2009, 199, 116–123. [Google Scholar] [CrossRef]
- Wang, Y.; Kelly, C.G.; Karttunen, J.T.; Whittall, T.; Lehner, P.J.; Duncan, L.; MacAry, P.; Younson, J.S.; Singh, M.; Oehlmann, W.; et al. CD40 is a Cellular Receptor Mediating Mycobacterial Heat Shock Protein 70 Stimulation of CC-Chemokines. Immunity 2001, 15, 971–983. [Google Scholar] [CrossRef]
- Vaughan, E.E.; de Vries, M.C.; Zoetendal, E.G.; Ben-Amor, K.; Akkermans, A.D.; de Vos, W.M. The Intestinal LABs. Antonie Van Leeuwenhoek 2002, 82, 341–352. [Google Scholar] [CrossRef]
- Sanchez, B.; Schmitter, J.M.; Urdaci, M.C. Identification of Novel Proteins Secreted by Lactobacillus rhamnosus GG Grown in De Mann-Rogosa-Sharpe Broth. Lett. Appl. Microbiol. 2009, 48, 618–622. [Google Scholar] [CrossRef]
- Reid, G. Lactobacillus in the Vagina: Why, how, which ones, and what do they do? In Lactobacillus Molecular Biology from Genomics to Probiotics; Ljungh, Å., Wadström, T., Eds.; Caister Academic Press: Norfolk, UK, 2009; p. 183. [Google Scholar]
- Durr, U.H.; Sudheendra, U.S.; Ramamoorthy, A. LL-37, the Only Human Member of the Cathelicidin Family of Antimicrobial Peptides. Biochim. Biophys. Acta 2006, 1758, 1408–1425. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Beveridge, T.J. Bacterial S-Layers. Trends Microbiol. 1999, 7, 253–260. [Google Scholar] [CrossRef]
- Bierne, H.; Dramsi, S. Spatial Positioning of Cell Wall-Anchored Virulence Factors in Gram-Positive Bacteria. Curr. Opin. Microbiol. 2012, 15, 715–723. [Google Scholar] [CrossRef]
- Egan, A.J.; Vollmer, W. The Physiology of Bacterial Cell Division. Ann. N. Y. Acad. Sci. 2013, 1277, 8–28. [Google Scholar] [CrossRef]
- Lutkenhaus, J. The ParA/MinD Family Puts Things in their Place. Trends Microbiol. 2012, 20, 411–418. [Google Scholar] [CrossRef]
- Benton, G.; Kleinman, H.K.; George, J.; Arnaoutova, I. Multiple Uses of Basement Membrane-Like Matrix (BME/Matrigel) in vitro and in vivo with Cancer Cells. Int. J. Canc. 2011, 128, 1751–1757. [Google Scholar] [CrossRef]
- Roos, S.; Jonsson, H. A High-Molecular-Mass Cell-Surface Protein from Lactobacillus reuteri 1063 Adheres to Mucus Components. Microbiology 2002, 148, 433–442. [Google Scholar]
- Sochacki, K.A.; Barns, K.J.; Bucki, R.; Weisshaar, J.C. Real-Time Attack on Single Escherichia coli Cells by the Human Antimicrobial Peptide LL-37. Proc. Natl. Acad. Sci. USA 2011, 108, E77–E81. [Google Scholar] [CrossRef]
- Eichenbaum, Z.; Green, B.D.; Scott, J.R. Iron Starvation Causes Release from the Group A Streptococcus of the ADP-Ribosylating Protein Called Plasmin Receptor Or Surface Glyceraldehyde-3-Phosphate-Dehydrogenase. Infect. Immun. 1996, 64, 1956–1960. [Google Scholar]
- Kline, K.A.; Dodson, K.W.; Caparon, M.G.; Hultgren, S.J. A Tale of Two Pili: Assembly and Function of Pili in Bacteria. Trends Microbiol. 2010, 18, 224–232. [Google Scholar] [CrossRef]
- Proft, T.; Baker, E.N. Pili in Gram-Negative and Gram-Positive Bacteria—Structure, Assembly and their Role in Disease. Cell Mol. Life Sci. 2009, 66, 613–635. [Google Scholar] [CrossRef]
- Kukkonen, M.; Saarela, S.; Lähteenmäki, K.; Hynönen, U.; Westerlund-Wikstrom, B.; Rhen, M.; Korhonen, T.K. Identification of Two Laminin-Binding Fimbriae, the Type 1 Fimbria of Salmonella enterica Serovar Typhimurium and the G Fimbria of Escherichia coli, as Plasminogen Receptors. Infect. Immun. 1998, 66, 4965–4970. [Google Scholar]
- Lähteenmäki, K.; Kuusela, P.; Korhonen, T.K. Bacterial Plasminogen Activators and Receptors. FEMS Microbiol. Rev. 2001, 25, 531–552. [Google Scholar] [CrossRef]
- Parkkinen, J.; Hacker, J.; Korhonen, T.K. Enhancement of Tissue Plasminogen Activator-Catalyzed Plasminogen Activation by Escherichia coli S Fimbriae Associated with Neonatal Septicaemia and Meningitis. Thromb. Haemost. 1991, 65, 483–486. [Google Scholar]
- Ben Nasr, A.; Olsen, A.; Sjobring, U.; Muller-Esterl, W.; Bjorck, L. Assembly of Human Contact Phase Proteins and Release of Bradykinin at the Surface of Curli-Expressing Escherichia coli. Mol. Microbiol. 1996, 20, 927–935. [Google Scholar] [CrossRef]
- Lähteenmäki, K.; Westerlund, B.; Kuusela, P.; Korhonen, T.K. Immobilization of Plasminogen on Escherichia Coli Flagella. FEMS Microbiol. Lett. 1993, 106, 309–314. [Google Scholar] [CrossRef]
- Bucior, I.; Pielage, J.F.; Engel, J.N. Pseudomonas aeruginosa Pili and Flagella Mediate Distinct Binding and Signaling Events at the Apical and Basolateral Surface of Airway Epithelium. PLoS Pathog. 2012, 8, e1002616. [Google Scholar] [CrossRef]
- Scharfman, A.; Arora, S.K.; Delmotte, P.; van Brussel, E.; Mazurier, J.; Ramphal, R.; Roussel, P. Recognition of Lewis x Derivatives Present on Mucins by Flagellar Components of Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5243–5248. [Google Scholar] [CrossRef]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but Not Hyperfimbriated Salmonella enterica Serovar Typhimurium Attaches to and Forms Biofilms on Cholesterol-Coated Surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef]
- Giron, J.A.; Torres, A.G.; Freer, E.; Kaper, J.B. The Flagella of Enteropathogenic Escherichia coli Mediate Adherence to Epithelial Cells. Mol. Microbiol. 2002, 44, 361–379. [Google Scholar] [CrossRef]
- Erdem, A.L.; Avelino, F.; Xicohtencatl-Cortes, J.; Giron, J.A. Host Protein Binding and Adhesive Properties of H6 and H7 Flagella of Attaching and Effacing Escherichia coli. J. Bacteriol. 2007, 189, 7426–7435. [Google Scholar] [CrossRef]
- Mahajan, A.; Currie, C.G.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, T.N.; Roe, A.; La Ragione, R.M.; Woodward, M.J.; et al. An Investigation of the Expression and Adhesin Function of H7 Flagella in the Interaction of Escherichia coli O157: H7 with Bovine Intestinal Epithelium. Cell. Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Yao, Y.; Kim, K.S. Flagella Promote Escherichia coli K1 Association with and Invasion of Human Brain Microvascular Endothelial Cells. Infect. Immun. 2007, 75, 2937–2945. [Google Scholar] [CrossRef]
- Troge, A.; Scheppach, W.; Schroeder, B.O.; Rund, S.A.; Heuner, K.; Wehkamp, J.; Stange, E.F.; Oelschlaeger, T.A. More than a Marine Propeller—The Flagellum of the Probiotic Escherichia coli Strain Nissle 1917 is the Major Adhesin Mediating Binding to Human Mucus. Int. J. Med. Microbiol. 2012, 302, 304–314. [Google Scholar] [CrossRef]
- Finkelstein, R.A.; Boesman-Finkelstein, M.; Chang, Y.; Hase, C.C. Vibrio cholerae hemagglutinin/protease, Colonial Variation, Virulence, and Detachment. Infect. Immun. 1992, 60, 472–478. [Google Scholar]
- Mel, S.F.; Fullner, K.J.; Wimer-Mackin, S.; Lencer, W.I.; Mekalanos, J.J. Association of Protease Activity in Vibrio cholerae Vaccine Strains with Decreases in Transcellular Epithelial Resistance of Polarized T84 Intestinal Epithelial Cells. Infect. Immun. 2000, 68, 6487–6492. [Google Scholar] [CrossRef]
- Finkelstein, R.A.; Boesman-Finkelstein, M.; Holt, P. Vibrio cholerae hemagglutinin/lectin/protease Hydrolyzes Fibronectin and Ovomucin: F.M. Burnet Revisited. Proc. Natl. Acad. Sci. USA 1983, 80, 1092–1095. [Google Scholar] [CrossRef]
- Korhonen, T.K.; Haiko, J.; Laakkonen, L.; Järvinen, H.M.; Westerlund-Wikstrom, B. Fibrinolytic and Coagulative Activities of Yersinia pestis. Front. Cell. Infect. Microbiol. 2013, 3, 35. [Google Scholar]
- Lähteenmäki, K.; Virkola, R.; Saren, A.; Emody, L.; Korhonen, T.K. Expression of Plasminogen Activator Pla of Yersinia pestis Enhances Bacterial Attachment to the Mammalian Extracellular Matrix. Infect. Immun. 1998, 66, 5755–5762. [Google Scholar]
- Zhang, S.S.; Park, C.G.; Zhang, P.; Bartra, S.S.; Plano, G.V.; Klena, J.D.; Skurnik, M.; Hinnebusch, B.J.; Chen, T. Plasminogen Activator Pla of Yersinia pestis Utilizes Murine DEC-205 (CD205) as a Receptor to Promote Dissemination. J. Biol. Chem. 2008, 283, 31511–31521. [Google Scholar] [CrossRef]
- Driessen, A.J.; Nouwen, N. Protein Translocation Across the Bacterial Cytoplasmic Membrane. Annu. Rev. Biochem. 2008, 77, 643–667. [Google Scholar] [CrossRef]
- Edelman, S.M.; Lehti, T.A.; Kainulainen, V.; Antikainen, J.; Kylväjä, R.; Baumann, M.; Westerlund-Wikström, B.; Korhonen, T.K. Identification of a High-Molecular-Mass Lactobacillus Epithelium Adhesin (LEA) of Lactobacillus crispatus ST1 that Binds to Stratified Squamous Epithelium. Microbiology 2012, 158, 1713–1722. [Google Scholar] [CrossRef]
- Ojala, T.; Kuparinen, V.; Koskinen, J.P.; Alatalo, E.; Holm, L.; Auvinen, P.; Edelman, S.; Westerlund-Wikström, B.; Korhonen, T.K.; Paulin, L.; et al. Genome Sequence of Lactobacillus crispatus ST1. J. Bacteriol. 2010, 192, 3547–3548. [Google Scholar] [CrossRef]
- Bergmann, S.; Hammerschmidt, S. Fibrinolysis and Host Response in Bacterial Infections. Thromb. Haemost. 2007, 98, 512–520. [Google Scholar]
- Redlitz, A.; Plow, E.F. Receptors for Plasminogen and t-PA: An Update. Baillieres Clin. Haematol. 1995, 8, 313–327. [Google Scholar] [CrossRef]
- Rijken, D.C.; Lijnen, H.R. New Insights into the Molecular Mechanisms of the Fibrinolytic System. J. Thromb. Haemost. 2009, 7, 4–13. [Google Scholar] [CrossRef]
- Aguero, G.; Villena, J.; Racedo, S.; Haro, C.; Alvarez, S. Beneficial Immunomodulatory Activity of Lactobacillus casei in Malnourished Mice Pneumonia: Effect on Inflammation and Coagulation. Nutrition 2006, 22, 810–819. [Google Scholar] [CrossRef]
- Haro, C.; Villena, J.; Zelaya, H.; Alvarez, S.; Aguero, G. Lactobacillus casei Modulates the Inflammation-Coagulation Interaction in a Pneumococcal Pneumonia Experimental Model. J. Inflamm. (Lond.) 2009, 6, 28. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Kainulainen, V.; Korhonen, T.K. Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria. Biology 2014, 3, 178-204. https://doi.org/10.3390/biology3010178
AMA Style
Kainulainen V, Korhonen TK. Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria. Biology. 2014; 3(1):178-204. https://doi.org/10.3390/biology3010178
Chicago/Turabian StyleKainulainen, Veera, and Timo K. Korhonen. 2014. "Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria" Biology 3, no. 1: 178-204. https://doi.org/10.3390/biology3010178