
Helenus and Ajax, Two Groups of Non-Autonomous LTR Retrotransposons, Represent a New Type of Small RNA Gene-Derived Mobile Elements

Genetic Information Research Institute, Cupertino, CA 95014, USA
Biology 2024, 13(2), 119; https://doi.org/10.3390/biology13020119
Submission received: 14 December 2023
/
Revised: 6 February 2024
/
Accepted: 10 February 2024
/
Published: 13 February 2024
(This article belongs to the Special Issue De Novo Detection of Transposons)
Abstract
:Simple Summary
Long terminal repeat (LTR) retrotransposons and non-LTR retrotransposons are two major repetitive components of eukaryotic genomes. Short intersperse elements (SINEs) are non-autonomous non-LTR retrotransposons with a sequence similar to small RNAs such as tRNA and 5S rRNA. Cassandra is a non-autonomous LTR retrotransposon with a 5S rRNA-like sequence in its LTRs. This observation led me to investigate the presence of non-autonomous LTR retrotransposons containing other small RNA sequences in their LTRs. Here, I discover and describe non-autonomous LTR retrotransposons Helenus and Ajax, with LTR regions of respective tRNA and 5S rRNA origin. Helenus and Ajax are likely mobilized by autonomous LTR retrotransposons. Helenus or Ajax are unlikely to encode a functional RNA. The finding of Helenus and Ajax confirms that non-autonomous LTR retrotransposons can be a source of small RNA-like interspersed sequences in the eukaryotic genomes. An understanding of their biology could contribute to the more precise prediction of small RNA genes and cellular RNA components as well as to the understanding of their evolutionary impacts.
Abstract
Terminal repeat retrotransposons in miniature (TRIMs) are short non-autonomous long terminal repeat (LTR) retrotransposons found from various eukaryotes. Cassandra is a unique TRIM lineage which contains a 5S rRNA-derived sequence in its LTRs. Here, two new groups of TRIMs, designated Helenus and Ajax, are reported based on bioinformatics analysis and the usage of Repbase. Helenus is found from fungi, animals, and plants, and its LTRs contain a tRNA-like sequence. It includes two LTRs and between them, a primer-binding site (PBS) and polypurine tract (PPT) exist. Fungal and plant Helenus generate 5 bp target site duplications (TSDs) upon integration, while animal Helenus generates 4 bp TSDs. Ajax includes a 5S rRNA-derived sequence in its LTR and is found from two nemertean genomes. Ajax generates 5 bp TSDs upon integration. These results suggest that despite their unique promoters, Helenus and Ajax are TRIMs whose transposition is dependent on autonomous LTR retrotransposon. These TRIMs can originate through an insertion of SINE in an LTR of TRIM. The discovery of Helenus and Ajax suggests the presence of TRIMs with a promoter for RNA polymerase III derived from a small RNA gene, which is here collectively termed TRIMp3.
Keywords:
LTR retrotransposon; TRIM; tRNA; 5S rRNA; Helenus; Ajax; RNA polymerase III promoter; SINE; Cassandra; TRIMp3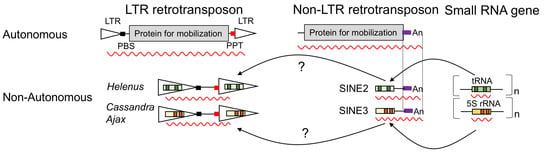
1. Introduction
Retrotransposons are mobile genetic elements that propagate their copies through reverse transcription of their transcribed RNA [1,2]. Retrotransposons encode a reverse transcriptase (RT), synthesizing DNA with an RNA template. The majority of retrotransposons are classified into two groups: long terminal repeat (LTR) retrotransposons and non-LTR retrotransposons [3]. LTR retrotransposons contain two identical LTRs at both ends (Figure 1). In many eukaryotes, retrotransposons occupy a substantial portion of their genomes. The human genome contains more non-LTR retrotransposons than LTR retrotransposons [4], while most plant genomes contain much more LTR retrotransposons than non-LTR retrotransposons [5,6].
LTR retrotransposons and non-LTR retrotransposons employ two distinct mechanisms for their mobilization [7,8]. LTR retrotransposons encode an integrase and a ribonuclease H (RNase H) in addition to an RT. The transcription of an LTR retrotransposon starts inside its 5′ LTR and ends inside its 3′ LTR. It generates identical sequences (R region) at both ends of the RNA. In the mobilization of LTR retrotransposons, an RT reverse-transcribes the retrotransposon RNA using a cellular tRNA as a primer. tRNA is bound to its complementary sequence downstream of 5′ LTR, called the primer-binding site (PBS) (Figure 1). Once the DNA synthesis reaches the 5′ R region of the retrotransposon RNA, the synthesized DNA jumps to the 3′ R region, thanks to the sequence identity between two R regions, to continue reverse transcription. Once the first DNA strand is synthesized, the retrotransposon RNA is degraded by RNase H, except for the polypurine tract (PPT), which tolerates the degradation and is used as a primer for the second strand synthesis. Finally, the double-strand DNA is generated as an extrachromosomal linear DNA, and then, the DNA is integrated into the genome by the integrase. Upon integration, a short genomic sequence is duplicated at both sides of the retrotransposon (target site duplications, TSDs). Non-LTR retrotransposons do not encode an integrase but encode an endonuclease instead. In the transposition mechanism of non-LTR retrotransposons, one genomic DNA strand cleaved by an endonuclease works as a primer for reverse transcription [9]. This mechanism is called target-primed reverse transcription (TPRT). The RT of non-LTR retrotransposons is usually able to distinguish its RNA from other RNAs to initiate reverse transcription [10].
LTR and non-LTR retrotransposons have non-autonomous elements that do not encode an RT. Short interspersed elements (SINEs) are the dominant group dependent on non-LTR retrotransposons for their mobilization [3,11]. SINEs often contain two or more components, originating independently. The 5′ terminal portion of SINEs, called “head”, contributes to their transcription. The 3′ region, called “tail”, mimics the tail of autonomous non-LTR retrotransposons and is recognized by their transposition machinery [10]. While non-LTR retrotransposons are transcribed by RNA polymerase II, SINEs are transcribed by RNA polymerase III [12]. The majority of SINEs contain an internal promoter derived from tRNA in their heads [3]. At least two other types of promoters contribute to the transcription of SINEs. One is the 7SL RNA-derived promoter in SINE1, represented by the human Alu family [13]. The other is the 5S rRNA-derived promoter seen in SINE3 [14]. SINE1 and SINE3 are known to be born independently multiple times [15,16].
LTR retrotransposons also have short non-autonomous elements, called terminal repeat retrotransposons in miniature (TRIMs) [17] or small LTR retrotransposons (SMARTs) [18]. TRIMs were first reported from plant genomes and are widely distributed from land plants to green algae [17,19]. Outside plants, TRIMs have been reported from hymenopteran insects [20,21,22], Taeniid cestodes [23], and bivalves [24]. TRIMs are shorter than 1000 bp in length and do not encode any proteins responsible for mobilization. Still, TRIMs contain two LTRs at both sides, and in their internal region, a PBS and a PPT are found at the termini. The sequence similarity between TRIMs and autonomous LTR retrotransposons suggests that TRIMs are transposed in trans by their counterpart LTR retrotransposons that may belong to Copia or Gypsy [19]. SMARTs are the reported shortest non-autonomous LTR retrotransposon group, which are only 292 bp long and are likely mobilized by a Copia superfamily of LTR retrotransposon [18].
Cassandra is an extraordinary group of TRIMs; it carries a 5S rRNA sequence in its LTRs [25] (Figure 1). The sequence comparison of Cassandra clearly shows high sequence conservation of RNA polymerase III promoters and terminators in their 5S sequences. The transcription of Cassandra initiates from the 5S sequence inside of the 5′ LTR and terminates in the 5S sequence of the 3′ LTR, generating short 18 bp duplications at both ends of the RNA. Cassandra is distributed widely in land plants and likely dates back to the Permian, 250 MYA [26].
The sequence derived from 5S rRNA plays a role in the transcription of Cassandra. It is analogous to the function of the 5S rRNA sequence in SINE3. This observation led me to investigate the presence of TRIMs containing other small RNA sequences in their LTRs. Here, two new types of TRIMs, Helenus and Ajax, are reported. Helenus is found in fungi, animals, and plants, and includes a tRNA-derived sequence inside its LTRs. Ajax is found in nemertean, and its LTRs include a part of the 5S rRNA sequence. In both Helenus and Ajax, the sequence conservation is observed around the promotor motifs in small RNA-derived sequences, while their predicted secondary structures of RNA do not support their functions as either tRNA or 5S rRNA. The findings of Helenus and Ajax suggest the universality of captured small RNA sequences working as promoters in mobile genetic elements.
2. Materials and Methods
2.1. Screening of TRIMs with tRNA-Derived Sequences
Repbase [3] was downloaded from the website of the Genetic Information Research Institute (last access: 27 November 2023). The genome assemblies were downloaded from the NCBI Assembly (https://www.ncbi.nlm.nih.gov/assembly/) (last access: 6 November 2023) (Table S1). Censor [27] searches were performed against the LTR dataset extracted from Repbase with the tRNA sequences or a subset of SINE2 sequences, both of which were also extracted from Repbase. All Censor hits were manually investigated, and only TRIMs shorter than 1000 bp in length were used for further analysis. TRIMs with tRNA-derived sequences in their LTRs were designated as Helenus.
Censor searches were performed with a Helenus consensus sequence against the genome assemblies that are evolutionarily related to the genome harboring the respective Helenus family (Table S1). Censor hits were extracted and clustered with BLASTCLUST 2.2.25 in the NCBI BLAST package with the thresholds at 75% length coverage and 75% sequence identity. The consensus sequence for each cluster was generated with the 50% majority rule applied with the help of homemade scripts. Censor search was performed with the consensus sequence of each cluster against the respective genome. Up to 10 Censor hits were extracted with 500 bp flanking sequences at both sides. The consensus sequence for each cluster was regenerated to be elongated to reach both termini. The termini were determined based on the presence of LTRs and 4 to 6 bp TSDs.
The prediction of tRNA sequences in all Helenus consensus sequences was performed on the tRNAscan-SE server (http://lowelab.ucsc.edu/tRNAscan-SE/index.html) (last access: 17 November 2023) [28]. All predicted tRNA sequences from Puccinia graminis, Puccinia striiformis, Puccinia triticina, Uromyces viciae-fabae, Melampsora larici-populina, Cronartium ribicola, Saccharomyces cerevisiae, Aplysia californica, Apis mellifera, and Homo sapiens were downloaded from GtRNAdb (http://gtrnadb.ucsc.edu/index.html) (last access: 21 November 2023) [29]. Sequence comparison between the predicted tRNA sequences and Helenus consensus sequences was performed with Censor with default parameters.
Sequence alignment was performed with MUSCLE v.3.8.1551 [30] with default parameters. The phylogenetic tree based on the tRNA-like sequences of the fungal Helenus families was reconstructed using the BioNJ method implemented in SEAVIEW 4.6.1 [31], with the J-C distances and 100 bootstrapping supports.
2.2. Prediction of tRNA Primer Usage and Similarity
To determine PBS and PPT sequences, the 5′ and 3′ terminal 50 bp sequences of the internal portions of LTR retrotransposons from Puccinia hordei, Crassostrea gigas, Nasonia vitripennis, and Danio rerio were extracted from Repbase (last access: 27 November 2023) [3]. The 5′ terminal sequences of Helenus and autonomous LTR retrotransposons were aligned with MUSCLE v.3.8.1551 [30] with default parameters. The 5′ sequences were then manually compared with the tRNA datasets from GtRNAdb (http://gtrnadb.ucsc.edu/index.html) (last access: 21 November 2023) [29].
2.3. Screening of TRIMs with 5S rRNA-Derived Sequences
RepeatModeler (https://www.repeatmasker.org/RepeatModeler/) was used for the initial screening of TEs from the genome of nemertean Notospermus geniculatus. Censor [27] searches were performed against the genome with the consensus sequences of repeats generated by RepeatModeler. Up to 10 Censor hits were extracted with 5000 bp flanking sequences at both sides. Consensus sequences were regenerated to be elongated to reach both termini. A TRIM family with a 5S rRNA-like sequence was identified and named as Ajax-1_NGe. Censor searches were performed with Ajax-1_NGe from N. geniculatus against the assembly of another nemertean genome, Lineus longissimus. Consensus sequences were generated as described above.
The secondary structures of LTRs of Ajax were predicted on the RNAfold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi) (last access: 28 November 2023) and were compared with the secondary structures reported in 5SrRNAdb (http://combio.pl/rrna/) (last access: 28 November 2023).
All reported consensus sequences for Helenus and Ajax are available in the Supplementary Materials as well as in Repbase (https://www.girinst.org/repbase/) [3].
2.4. Characterization of SINE3 from Lepidopteran Genomes
The genome assemblies of four lepidopteran insects (the common copper brome Lycaena phlaeas (ilLycPhla1.1), the garden grass-veneer Chrysoteuchia culmella (ilChrCulm1.1), the dusky thorn Ennomos fuscantarius (ilEnnFusc2.1), and the ringed china-mark Parapoynx stratiotata (ilParStra1.1)) were downloaded from the NCBI Assembly (https://www.ncbi.nlm.nih.gov/assembly/) (last access: 6 November 2023). Censor [27] searches were performed with the HaSE3 consensus sequence against the genome assemblies. Consensus sequences were generated as described above. The termini were determined based on the sequence similarity to HaSE3 or other SINEs in Repbase [3].
Sequence alignment of Ajax, SINE3, and 5S rRNA sequences was performed with MUSCLE v.3.8.1551 [30] with default parameters.
2.5. Estimation of Copy Numbers
Censor [27] searches were performed with six genomes (P. triticina, C. ribicola, D. rerio, C. gigas, N. vitripennins, and L. longissimus) as queries against their respective Helenus/Ajax consensus sequences. The hits longer than 90% of the respective consensus and with over 90% sequence identity were assumed as full length. The LTR and the internal portion were counted independently.
3. Results
3.1. Helenus: TRIM with a tRNA-Derived Sequence
Censor [27] searches with tRNA and SINE2 sequences against LTRs in Repbase [3] revealed that not a few LTRs contain a sequence similar to a part of tRNA. Among them, LTR-1_PGr from the rust P. graminis, LTR-1_PSt and LTR-2_PSt from P. striiformis, LTR-1_PTrit from P. triticina, LTR1_LCh and LTR2_Lch from the coelacanth Latimeria charmnae, LTR-11_DR from the zebrafish D. rerio, and ULTR-1_NVit from the jewel wasp N. vitripennis could be classified as TRIMs based on their lengths and structures (Table S1). Here, the TRIMs with tRNA-derived sequences in their LTRs are designated as Helenus, the twin brother of Cassandra.
3.2. Helenus from Fungi
Using characterized Helenus sequences as seeds, the extensive analysis to characterize LTRs with tRNA-derived sequences was performed (Table 1 and Table S1). In fungi, Pucciniomycetes including P. graminis, P. striiformis, and P. triticina harbor many Helenus families. Helenus families were characterized in the genera Puccinia, Austropuccinia, Melampsora, Uromyces, Cronartium, and Hemileia, all in the order Pucciniales. Outside of Pucciniales, no Helenus was found from Pucciniomycetes, including Microbotryum, Leucosporidium, and Rhodotorula. Helenus was also detected from several Agaricomycetes fungi: Agaricus bisporus, Coprinopsis cinerea, and Amylostereum areolatum. In total, 113 Helenus families were found from Pucciniales fungi, and multiple Helenus families co-exist in the same genome. Over 10 Helenus families were found from P. coronata, P. horiana, P. triticina, and C. ribicola. These results suggest that Helenus has been maintained in the genomes of Pucciniales fungi. Pucciniales fungi date back to the Jurassic, ~130 million years ago (MYA) [32].
Some Helenus families retain no full-length copies, but rather fragments and solo LTRs, such as Helenus-6_PSt and Helenus-7_PHord (Table S1). Many of the solo LTRs of Helenus are flanked by 5 bp TSDs as they have experienced LTR-LTR recombination after transposition.
3.3. Helenus from Animals
In animals, six phyla (Arthropoda, Brachiopoda, Chordata, Ctenophora, Mollusca, and Nemertea) include at least one species whose genome maintains Helenus (Table 1 and Table S1). In Brachiopoda, Lingula anatina contains at least three families of Helenus. The genome of Mnemiopsis leidyi in Ctenophora contains a Helenus family. In Nemertea, two related ribbon worms L. longissimus and N. geniculatus, both in the family Lineidae, have Helenus families.
In Arthropoda, using ULTR-1_NVi as a seed, diverse Helenus families were characterized from six families of parasitoid jewel wasps (Pteromalidae, Eurytomidae, Eupelmidae, Megastigmidae, Cynipidae, and Torymidae) in the superfamily Chalcidoidea, Hymenoptera. In total, 40 Helenus families were found from 14 genomes. No Helenus family was found in Trichogrammatidae, Ormyridae, Eulophidae, Aphelinidae, Mymaridae, Eucyrtidae, Agaonidae, or Chalcididae in Chalcidoidea. The common ancestor of the wasps with Helenus dates back to the Early Cretaceous, over 100 MYA [33]. Hymenopteran Helenus families include the longest family among Helenus found in this study. The longest Helenus family is Helenus-1_SyUm from Synergus umbraculus, which is 1263 bps. Its internal portion is also the longest, 621 bps, and its LTRs are the second longest, 321 bps, only two bps shorter than the LTR of Helnus-1_SyJa, 323 bps from Synergus japonicus.
In Chordata, the genome of coelacanth L. chalumnae contains two families of Helenus: LTR1_LCh and LTR2_Lch [34]. The reported consensus sequences of these two families include both LTRs and the internal sequence. The sterlet Acipenser ruthenus and American paddlefish Polyodon spathula are distant relatives but contain closely related Helenus families whose consensus sequences are ~98% identical to each other. LTR-11_DR from the zebrafish D. rerio also shows similarity to tRNA sequences in its LTRs. Helenus families related to LTR-11_DR are found in various Cypriniformes genomes and the two genomes outside of Cypriniformes: Asian redtail catfish Hemibagrus wyckioides (Siluriformes) and the milkfish Chanos chanos (Gonorynchiformes). These three orders belong to the superorder Ostariophysi. Some copies of Helenus-1_CaAu from the goldfish Carassius auratus are ~99.5% identical to their consensus, showing its very recent activity. On the other hand, these LTR-11_DR-related Helenus families are very conserved in sequence. Helenus-1_CaAu has the internal portion completely identical to that of LTR-11_DR. The distribution and sequence conservation indicate that LTR-11_DR-related Helenus families date back to the Triassic, >200 MYA [35].
In Mollusca, bivalves in the five orders (Ostreida, Pterioida, Mytilida, Pectinida, and Arcoida) in the subclass Pteriomorphia contain at least one Helenus family in their genomes. In total, 51 Helenus families were found from these five orders. Limoida, the other order in Pteriomorphia, includes no available genome assembly as of Nov. 22, 2023. The wide distribution in Pteriomorphia suggests the origin of Helenus in the common ancestor of Pteriomorphia, which dates back to the Early Ordovician, over 470 MYA [36].
3.4. Helenus from Plants
3.5. tRNA-Derived Sequences in Helenus
The comparison between tRNA genes and Helenus LTRs revealed the conservation around boxes A and B, responsible for the RNA polymerase III transcription (Figure 2). Most Helenus LTRs contain TRGCY and YGG in box A, and GGTTC is conserved in box B. When tRNAScan-SE [28] is used to determine the types of tRNA sequences in Helenus LTRs, the highest scoring isotype model does not match the anticodon-inferred isotype in many Helenus families. Such isotype prediction disagreement (IPD) indicates that the predicted tRNA is unfunctional. When excluding these cases, only a few Helenus from bivalves and fungi are predicted to encode a tRNA (Figure S1). In bivalves, seven Helenus families can encode a tRNA-Thr. In fungi, 15 Helenus families can encode a tRNA for His or Thr. This raises the possibility that Helenus contributes to the expansion of tRNA genes.
GtRNADB (http://gtrnadb.ucsc.edu/) (last access: 21 November 2023) [29] reports substantial numbers of predicted tRNA genes in Pucciniales fungi. P. triticina is predicted to have 1538 tRNA genes, including 225 predicted pseudogenes. They include 492 tRNA-Thr-AGT, 189 tRNA-Ser-TGA, 137 tRNA-Ala-TGC, and 128 tRNA-Pro-TGG. P. striiformis is predicted to have 890 tRNA genes including 138 predicted pseudogenes. P. graminis is predicted to have 506 tRNA genes including 103 predicted pseudogenes. Outside the genus Puccinia, Uromyces viciae-fabae contains 588 predicted tRNA genes, and Melanopsola larici-populina has 251 predicted tRNA genes. Cronartium ribicola is predicted to have 1670 tRNA genes, including 519 predicted pseudogenes. No predicted tRNA dataset is available for mollusks except Aplysia californica in GtRNADB.
There is an expansion of certain types of tRNA genes in Pucciniales fungi. In the genus Puccinia, tRNA-Thr-AGT, tRNA-Ser-TGA, tRNA-Ser-AGA, tRNA-Ala-TGC, tRNA-Ala-AGC, and tRNA-Pro-TGG are predicted in extremely high copy numbers. Many of these predicted tRNA genes correspond to the tRNA-like sequences in one or more Helenus families (Table 2). When excluding predicted tRNA genes in Helenus copies, only a small number of predicted tRNA genes remain in each Puccinia genome. In the genera Uromyces and Cronartium, numerous genes for tRNA-Thr-AGT, tRNA-Ser-AGA, tRNA-Sup-CTA (suppressor tRNA), tRNA-Ala-AGC, and tRNA-Val-GAC are predicted; however, after excluding predicted tRNA genes showing high similarity to Helenus, less than 10 genes for each tRNA type remain in each genome. In the cases of tRNA-Sup-CTA, all predicted genes originate from Helenus.
Among the Helenus families that show sequence similarity to predicted tRNA genes of high copy number (Table 2), only two families, Helenus-4_PSt and Helenus-4_PGr, are able to encode a functional tRNA (Figure S1). They may contribute to the tRNA gene increase by 21 and 15, respectively (Table 2). Even if they are functional, their contribution to the tRNA gene increase is limited. Helenus is less likely to contribute to the diversity and expansion of functional tRNAs, although functional validation is needed to eliminate that possibility.
3.6. Primer-Binding Sites (PBSs) of Helenus
A primer-binding site (PBS) is located near the 5′ terminus of internal regions, just downstream of the junction of the 5′ LTR. Most LTR retrotransposons and retroviruses use the 3′ end of a tRNA as a primer for reverse transcription, and the PBS is often a sequence complementary to the 3′ end of tRNA. Hymenopteran Helenus families use the 3′ end of either tRNA-Lys, tRNA-Leu, tRNA-Trp, or tRNA-Arg as a primer, determined based on the sequence complementarity between the PBS and tRNA (Figure S2A). Vertebrate Helenus families use the 3′ end of tRNA-His, tRNA-Leu, or tRNA-Lys (Figure S2B). Helenus families from bivalves use the 3′ end of tRNA-Lys, tRNA-His, tRNA-Ile, tRNA-Pro, tRNA-Glu, or tRNA-Ser as a primer (Figure S2C).
In contrast, only several fungal Helenus families use the 3′ end of tRNA as a primer for reverse transcription. The majority of fungal Helenus families contain PBSs complementary to the anticodon loop of tRNA (Figure 3). The anticodon loop sequence of tRNA-Tyr, tRNA-Ile, tRNA-Asp, and tRNA-iMet appears to be used as a primer for reverse transcription. These four types of tRNA genes contain an intron downstream of their anticodons. The PBS sequences suggest the usage of mature tRNA (without intron) as a primer for reverse transcription.
Helenus and some autonomous LTR retrotransposons show sequence similarity at the PBS region. Only autonomous Gypsy LTR retrotransposons show similarity to the PBS region of animal Helenus families. In bivalves, Gypsy-125_CGi uses tRNA-Lys as a primer, Gypsy-39_CGi uses tRNA-His, Gypsy-157_CGi uses tRNA-Ile, and Gypsy-13_CGi uses tRNA-Pro (Figure S2C). In insects, Gypsy-16_NVi uses tRNA-Lys as a primer for reverse transcription, Gypsy-28_NVi uses tRNA-Leu, Gypsy-32_NVi uses tRNA-Trp, and Gypsy-31_NVi uses tRNA-Arg (Figure S2A).
Unlike animal Helenus families, fungal Helenus families show sequence similarity at the PBS region with the Copia superfamily (Figure 3). Copia-50_PHord uses the anticodon loop of tRNA-Tyr as a primer, Copia-18_PHord uses that of tRNA-Ile, Copia-6_PHord uses that of tRNA-Asp, and Copia-28_PHord uses that of tRNA-iMet. Copia-39_PHord uses the 3′ end of tRNA-iMet as a primer for reverse transcription. This suggests that animal Helenus is mobilized by the Gypsy superfamily, while fungal Helenus is mobilized by the Copia superfamily of LTR retrotransposons.
The priming of reverse transcription at the anticodon loop of tRNA is known for Ty5 from Saccharomyces cerevisiae and Copia from Drosophila melanogaster, both belonging to the Copia superfamily of LTR retrotransposons [37,38]. A systematic analysis of plant LTR retrotransposons reported that the 3′-truncated tRNA primer is used only by several lineages in the Copia superfamily [39]. Finally, the lengths of TSDs of animal and fungal Helenus families show a clear distinction between them; animal Helenus families generate 4 bp TSDs, while fungal Helenus generates 5 bp TSDs. The Copia families showing similarity to Helenus PBSs generate 5 bp TSDs [3].
The sequence similarity between Copia-9_PGr and LTR-1_PTrit is a more direct support for the mobilization of fungal Helenus by Copia (Figure 4). The 5′ 198 bp and the 3′ 12 bp sequences of the internal portion of Copia-9_PGr correspond to the entire internal portion of LTR-1_PTrit. Because Copia-9_PGr is from P. graminis and LTR-1_PTrit is from P. triticina, LTR-1_PTrit is not mobilized by Copia-9_PGr. More likely, a close relative of Copia-9_PGr transposes LTR-1_PTrit. Helenus families related to LTR-1_PTrit are distributed among the genera Puccinia, Uromyces, and Cronartium, indicating the coevolution between Helenus and Copia. LTRs of Copia-9_PGr and LTR-1_PTrit are not similar to each other.
Different tRNA primer usage in fungal, hymenopteran, or molluscan Helenus lineages suggests the co-evolution between Helenus and its autonomous counterpart. As LTRs are not well conserved, a phylogenetic tree based on the tRNA-like sequences was produced for the fungal Helenus families (Figure 3). Due to the low conservation among Helenus families, the phylogeny does not have strong statistical support. The dominant tRNA primer type in the fungal Helenus families is the anticodon loop of tRNA-iMet-CAT. At least five lineages of Helenus changed their tRNA primer type. tRNA-Asn-GTT was chosen as a primer once in the evolution of fungal Helenus (Figure 3, green). tRNA-Ile-AAT was likely selected once (Figure 3, brown). The primer usage of tRNA-Tyr-GTA (Figure 3, blue) and the 3′ terminus of tRNA-iMet-CAT (Figure 3, purple) evolved multiple times.
One might think that the tRNA-like sequence in the LTRs works as a primer for reverse transcription. However, we could not find any sequence complementarity between the tRNA-like sequence in the LTRs and its corresponding PBS. Thus, the tRNA-like sequence in the LTRs does not contribute to the reverse transcription initiation.
3.7. Ajax: TRIM with a 5S rRNA Promoter
Routine screening of repetitive sequences from the nemertean Notospermus geniculatus revealed the presence of TRIMs whose LTR contains a fragment of the 5S rRNA gene (Figure 5). This TRIM lineage with a partial 5S rRNA-derived sequence in its LTRs is designated as Ajax. Ajax was found in two closely related nemerteans: N. geniculatus and L. longissimus (Table S2). The 5S rRNA-derived sequences in Ajax only cover the promoter region containing three motifs: box A, IE, and box C (Figure 3). As Ajax includes only a part of the 5S rRNA sequence, it is unlikely to encode a functional 5S rRNA. The secondary structure of LTR of Ajax-1_LiLo predicted by RNAfold (http://rna.tbi.univie.ac.at/) (last access: 28 November 2023) does not show similarity to the canonical structure of 5S rRNA. The sequence conservation of the promoter motifs suggests that the function of the 5S rRNA-derived sequence is to promote the transcription of Ajax by RNA polymerase III.
It is speculated that Cassandra originated from the retrotransposition of a SINE3 element into an LTR, which was then copied into the other LTR by canonical retrotransposition [25]. Ajax, SINE3, and 5S rRNA sequences were compared to investigate the origin of the 5S rRNA-like sequence in Ajax LTRs (Figure 3). A Censor search against Repbase revealed the sequence similarity between Ajax and HaSE3, a SINE3 family from the lepidopteran insect Helicoverpa armigera [16]. It is unlikely that HaSE3 contributed to the birth of Ajax considering the divergence of the host organisms, but the comparison could shed light on the evolution of Ajax. SINE3 families have been identified from other lepidopteran insects [40]. Four more SINE3 families were characterized here to compare the 5S rRNA-derived sequences; they are SINE3-1_LyPhl from the common copper brome Lycaena phlaeas, SINE3-1_ChrCul and SINE3-2_ChrCul from the garden grass-veneer Chrysoteuchia culmella, and SINE3-1_EnnFus from the dusky thorn Ennomos fuscantarius. The comparison revealed the higher sequence conservation in the promoter regions and the higher divergence in the upstream regions. It is noteworthy that Ajax shows a longer similar sequence to several SINE3 families: CsSE2 and SINE3-1_ChrCul. This suggests that the 5S rRNA-like sequence in Ajax is not a direct descendant of 5S rRNA. The sequence upstream of the promoter regions of Ajax might have diverged in the ancestral SINE3 family, then integrated into an LTR of the ancestor of Ajax.
The sequence downstream of the 5′ LTR is complimentary to the 3′ end of tRNA-Leu (Figure S3), indicating that tRNA-Leu works as a primer for reverse transcription.
3.8. Copy Number Estimates of Helenus and Ajax
The copy numbers of Helenus and Ajax were estimated from the six genomes of divergent species: P. triticina, C. ribicola, D. rerio, C. gigas, N. vitripennis, and L. longissimus (Table S3). Here, the sequences longer than 90% of the respective consensus were examined. The copy number of LTRs could be higher than twice that of internal portions, considering the presence of solo LTRs. The largest Helenus family in the two species of fungi is LTR-1_PTrit, which contains 529 LTRs and 144 internal portions. Small families such as Helenus-10_PTrit and Helenus-12_PTrit retain less than 10 LTRs. In the four analyzed animal species, LTR-11_DR shows enormous copies: 5575 LTRs and 2618 internal portions. Tandem arrays of LTR-11_DR are observed. Helenus-2C_CGi retains 1124 LTRs and 402 internal portions. Ajax-1_LiLo has 1114 LTRs and 151 internal portions.
4. Discussion
4.1. TRIMp3—Proposed TRIM Families Containing an RNA Polymerase III Promoter
Helenus, Ajax, and Cassandra [25] represent a unique type of TRIMs; they include a small RNA gene-derived sequence in their LTRs. Cassandra produces transcripts from the RNA polymerase III promoter. Without experimental validation, it cannot be concluded that Helenus and Ajax contain a functional RNA polymerase III promoter, but the sequence conservation indicates the functionality. Ajax includes only a part of the 5S rRNA sequence, likely the part only responsible for promoting the transcription by RNA polymerase III. Helenus is unlikely to encode a functional tRNA either. These observations indicate that although they do not exclude the possibility that these TRIM families encode a functional RNA, the main selective drive for such TRIMs is propagating themselves, not contributing to the repertoire of cellular RNA components.
These TRIMs share some properties with SINEs. Both contain a small RNA-derived sequence, likely functioning as an RNA polymerase III promoter. Because retrotransposon RNA works as a template for reverse transcription as well as for protein synthesis, the RNA length is a restriction factor for the retrotransposon length. The length of SINEs is restricted, related to the weak processivity of RNA polymerase III compared with RNA polymerase II. SINEs are 100 to 700 bp [11]. Helenus is between 311 and 1263 bp in length. Ajax is 421 (Ajax-1_LiLo) or 434 bp (Ajax-1_NGe). Cassandra is from 565 to 860 bp in length [25]. Thus, the length of these TRIMs seems also restricted by the processivity of RNA polymerase III.
Here, it is proposed to call the TRIM families containing an RNA polymerase III promoter together “TRIMp3.” Cassandra is the only validated TRIMp3 lineage to date, and Helenus and Ajax are expected to be added. Canonical TRIM families can be called “TRIMp2.” We can speculate a TRIM family with a 7SL RNA sequence. SINE1 has been found only in primates, rodents, and hagfishes [13,15,41]. It seems related to the reasons why a TRIM family with a 7SL RNA-derived sequence has not been found yet.
SINEs work as an enhancer, promoter, insulator, splicing signal, and transcription factor-binding site [42,43]. As TRIMp3 and SINEs are both short interspersed repetitive elements that remain a promoter recruiting RNA polymerase III, it is likely that many biological functions of SINEs can also be applied to TRIMp3. Rapid transcriptional regulatory rewriting mediated by TRIMp3 mobilization might happen in TRIMp3-rich lineages such as Pucciniales fungi and parasitoid wasps.
4.2. The Evolution of TRIMp3
SINEs are mobilized by non-LTR retrotransposons, while TRIMs are mobilized by LTR retrotransposons. The presence of the PBS and PPT in Helenus and Ajax supports that their retrotransposition mechanism is identical to that of most LTR retrotransposons. A tRNA primer works for the first-strand DNA synthesis, and the PPT contributes to the second-strand synthesis.
It is almost certain that Ajax and Cassandra have independent origins. The wide distribution of Cassandra in green plants suggests that they originated in the common ancestor of angiosperms and ferns, over 250 MYA [25]. Ajax is found only in two nemertean species and likely originated in this lineage. The divergent sequence upstream of the promoter motifs in Ajax LTRs (Figure 5) suggests that they originated by SINE3 insertion into the ancestral TRIM, as proposed for Cassandra.
SINE2 is much more abundant than SINE3, so it is also likely that Helenus was born multiple times through SINE2 insertion into an LTR of the ancestral TRIM. Their patchy distribution and diversity also support their multiple origins. Several lines of data suggest that fungal Helenus families depend on the Copia superfamily of LTR retrotransposon for their mobilization, while animal Helenus depend on the Gypsy superfamily of LTR retrotransposon. It is speculated that molluscan Helenus in the subclass Pteriomorphia originated > 470 MYA, fish Helenus in the superorder Ostariophysi ~200 MYA, fungal Helenus in the order Pucciniales ~130 MYA, and hymenopteran Helenus in the superfamily Chalcidoidea > 100 MYA in their respective ancestors.
During the long history of the evolution of Helenus, there have been many diversification events. Multiple Helenus families can co-exist in a single genome (Table S1). The speciation of Helenus families can occur independently from their counterpart LTR retrotransposons. In that case, multiple Helenus families depend on a single family of LTR retrotransposon for mobilization. It is also possible that the speciation of Helenus occurs along with the diversification of its counterpart LTR retrotransposon. Such co-speciation events could have happened given the observed PBS diversity. Although the detailed mechanism is unknown, LTR retrotransposons have changed their tRNA primer type during evolution [39]. Fungal Helenus also shows the change in tRNA primer type during evolution (Figure 3). Other groups of Helenus also show the diversity of PBS, so they also have experienced the change in tRNA primer type. The change in tRNA primer type by Helenus could have happened simultaneously with the change by its counterpart LTR retrotransposon. Autonomous Copia families reported from the genome of P. hordei show all varieties of tRNA primers used by fungal Helenus families (Figure 3). Long-term maintenance of Helenus must coincide with the continuous presence of active autonomous counterparts. Co-evolution between Helenus and its autonomous counterpart is the most likely scenario to explain their maintenance and diversity. Or Helenus could have shifted its tRNA primer type by changing its autonomous counterpart. The change in autonomous counterpart is common in SINE evolution [15].
It is not known how the dependence of TRIM on its autonomous LTR retrotransposon is maintained. The sequence similarity in LTRs between TRIMs and autonomous LTR retrotransposons is an indication of their relationships [19]. Some of these pairs also show sequence similarity in the internal portions between two LTRs. In the case of SMARTs, while there is no sequence similarity between the LTRs of SMARTs and their putative autonomous counterpart FRetro64, the internal portion of SMARTs shows high sequence identity to a part of FRetro64 [18]. Considering the weak processivity of RNA polymerase III, their counterpart autonomous LTR retrotransposons must use a promoter for RNA polymerase II similar to canonical LTR retrotransposons. Therefore, it is unlikely that a long stretch of sequence is shared between TRIMp3 and its autonomous counterpart in their LTRs. In fact, LTR-1_PTrit and related Helenus families include a sequence similar to the internal portion but not the LTRs of Copia-9_PGr (Figure 4). Although Cassandra also has a long history of evolution and likely has diversified into multiple lineages, we have no indication for the autonomous LTR retrotransposon family responsible for Cassandra mobilization. Another unique group of non-autonomous LTR retrotransposons, called retrozymes, also contain LTRs that do not share sequences with canonical LTR retrotransposons [44]. These observations suggest that the homology of LTRs is not necessary for trans-mobilization. Many recently active Copia families have been reported from various Puccinia species and Gypsy families from C. gigas [3]. They could be a source for further studies of the evolution, diversification, and mobilization of TRIMp3 families.
5. Conclusions
Here, two new groups of non-autonomous LTR retrotransposons are characterized: Helenus and Ajax. Helenus is found in fungi, animals, and plants and contains a tRNA-like sequence in its LTRs. Ajax is found in nemertean species and includes a short sequence corresponding to the promoter of the 5S rRNA gene. The finding of Helenus and Ajax suggests the presence of short non-autonomous LTR retrotransposons (TRIMs) with a small RNA-derived sequence functioning as a promoter for RNA polymerase III, here designated TRIMp3. These small RNA-derived sequences in TRIMp3 likely function to produce a template RNA for reverse transcription. There may be a direct relationship between TRIMp3s and SINEs; a SINE insertion into an LTR of TRIM may lead to the birth of TRIMp3.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biology13020119/s1, Figure S1: Structural alignment of predicted tRNA sequences encoded in Helenus; Figure S2: Primer-binding sites (PBSs) of Helenus families; Figure S3: Primer-binding sites (PBSs) of Ajax families; Table S1: List of Helenus families; Table S2: List of Ajax families; Table S3: Copy numbers of Helenus and Ajax families from six representative organisms. Data S1: Consensus sequences of Helenus and Ajax families reported in this study in the fasta format.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kojima, K.K. Structural and sequence diversity of eukaryotic transposable elements. Genes Genet. Syst. 2020, 94, 233–252. [Google Scholar] [CrossRef]
- Arkhipova, I.R. Using bioinformatic and phylogenetic approaches to classify transposable elements and understand their complex evolutionary histories. Mob. DNA 2017, 8, 19. [Google Scholar] [CrossRef]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Luo, X.; Chen, S.; Zhang, Y. PlantRep: A database of plant repetitive elements. Plant Cell Rep. 2022, 41, 1163–1166. [Google Scholar] [CrossRef]
- Stitzer, M.C.; Anderson, S.N.; Springer, N.M.; Ross-Ibarra, J. The genomic ecosystem of transposable elements in maize. PLoS Genet. 2021, 17, e1009768. [Google Scholar] [CrossRef]
- Curcio, M.J.; Derbyshire, K.M. The outs and ins of transposition: From mu to kangaroo. Nat. Rev. Mol. Cell Biol. 2003, 4, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Telesnitsky, A.; Goff, S.P. Reverse Transcriptase and the Generation of Retroviral DNA. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Luan, D.D.; Korman, M.H.; Jakubczak, J.L.; Eickbush, T.H. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site: A mechanism for non-LTR retrotransposition. Cell 1993, 72, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, M.; Okada, N. LINEs mobilize SINEs in the eel through a shared 3′ sequence. Cell 2002, 111, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Vassetzky, N.S.; Kramerov, D.A. SINEBase: A database and tool for SINE analysis. Nucleic Acids Res. 2013, 41, D83–D89. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Okada, N. Generality of the tRNA origin of short interspersed repetitive elements (SINEs). Characterization of three different tRNA-derived retroposons in the octopus. J. Mol. Biol. 1994, 243, 25–37. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L.; Hellmann-Blumberg, U.; Jurka, J.; Labuda, D.; Rubin, C.M.; Schmid, C.W.; Zietkiewicz, E.; Zuckerkandl, E. Standardized nomenclature for Alu repeats. J. Mol. Evol. 1996, 42, 3–6. [Google Scholar] [CrossRef]
- Kapitonov, V.V.; Jurka, J. A novel class of SINE elements derived from 5S rRNA. Mol. Biol. Evol. 2003, 20, 694–702. [Google Scholar] [CrossRef]
- Kojima, K.K. Hagfish genome reveals parallel evolution of 7SL RNA-derived SINEs. Mob. DNA 2020, 11, 18. [Google Scholar] [CrossRef]
- Wang, J.; Wang, A.; Han, Z.; Zhang, Z.; Li, F.; Li, X. Characterization of three novel SINE families with unusual features in Helicoverpa armigera. PLoS ONE 2012, 7, e31355. [Google Scholar] [CrossRef]
- Witte, C.P.; Le, Q.H.; Bureau, T.; Kumar, A. Terminal-repeat retrotransposons in miniature (TRIM) are involved in restructuring plant genomes. Proc. Natl. Acad. Sci. USA 2001, 98, 13778–13783. [Google Scholar] [CrossRef]
- Gao, D.; Chen, J.; Chen, M.; Meyers, B.C.; Jackson, S. A highly conserved, small LTR retrotransposon that preferentially targets genes in grass genomes. PLoS ONE 2012, 7, e32010. [Google Scholar] [CrossRef]
- Gao, D.; Li, Y.; Kim, K.D.; Abernathy, B.; Jackson, S.A. Landscape and evolutionary dynamics of terminal repeat retrotransposons in miniature in plant genomes. Genome Biol. 2016, 17, 7. [Google Scholar] [CrossRef]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 76. [Google Scholar] [CrossRef]
- Zhou, Y.; Cahan, S.H. A novel family of terminal-repeat retrotransposon in miniature (TRIM) in the genome of the red harvester ant, Pogonomyrmex barbatus. PLoS ONE 2012, 7, e53401. [Google Scholar] [CrossRef]
- Elsik, C.G.; Worley, K.C.; Bennett, A.K.; Beye, M.; Camara, F.; Childers, C.P.; de Graaf, D.C.; Debyser, G.; Deng, J.; Devreese, B.; et al. Finding the missing honey bee genes: Lessons learned from a genome upgrade. BMC Genom. 2014, 15, 86. [Google Scholar] [CrossRef]
- Koziol, U.; Radio, S.; Smircich, P.; Zarowiecki, M.; Fernandez, C.; Brehm, K. A Novel Terminal-Repeat Retrotransposon in Miniature (TRIM) Is Massively Expressed in Echinococcus multilocularis Stem Cells. Genome Biol. Evol. 2015, 7, 2136–2153. [Google Scholar] [CrossRef]
- Satovic, E.; Luchetti, A.; Pasantes, J.J.; Garcia-Souto, D.; Cedilak, A.; Mantovani, B.; Plohl, M. Terminal-Repeat Retrotransposons in Miniature (TRIMs) in bivalves. Sci. Rep. 2019, 9, 19962. [Google Scholar] [CrossRef]
- Kalendar, R.; Tanskanen, J.; Chang, W.; Antonius, K.; Sela, H.; Peleg, O.; Schulman, A.H. Cassandra retrotransposons carry independently transcribed 5S RNA. Proc. Natl. Acad. Sci. USA 2008, 105, 5833–5838. [Google Scholar] [CrossRef]
- Kalendar, R.; Raskina, O.; Belyayev, A.; Schulman, A.H. Long Tandem Arrays of Cassandra Retroelements and Their Role in Genome Dynamics in Plants. Int. J. Mol. Sci. 2020, 21, 2931. [Google Scholar] [CrossRef]
- Kohany, O.; Gentles, A.J.; Hankus, L.; Jurka, J. Annotation, submission and screening of repetitive elements in Repbase: RepbaseSubmitter and Censor. BMC Bioinform. 2006, 7, 474. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. GtRNAdb 2.0: An expanded database of transfer RNA genes identified in complete and draft genomes. Nucleic Acids Res. 2016, 44, D184–D189. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Aime, M.C.; Bell, C.D.; Wilson, A.W. Deconstructing the evolutionary complexity between rust fungi (Pucciniales) and their plant hosts. Stud. Mycol. 2018, 89, 143–152. [Google Scholar] [CrossRef]
- Cruaud, A.; Rasplus, J.Y.; Zhang, J.; Burks, R.; Delvare, G.; Fusu, L.; Gumovsky, A.; Huber, J.T.; Jansta, P.; Mitroiu, M.D.; et al. The Chalcidoidea bush of life: Evolutionary history of a massive radiation of minute wasps. Cladistics 2023, 40, 34–63. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, M.; Noguchi, H.; Nishihara, H.; Toyoda, A.; Suzuki, Y.; Kajitani, R.; Suzuki, H.; Okuno, M.; Aibara, M.; Ngatunga, B.P.; et al. Coelacanth genomes reveal signatures for evolutionary transition from water to land. Genome Res. 2013, 23, 1740–1748. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; Eytan, R.I.; Dornburg, A.; Kuhn, K.L.; Moore, J.A.; Davis, M.P.; Wainwright, P.C.; Friedman, M.; Smith, W.L. Resolution of ray-finned fish phylogeny and timing of diversification. Proc. Natl. Acad. Sci. USA 2012, 109, 13698–13703. [Google Scholar] [CrossRef]
- Combosch, D.J.; Giribet, G. Clarifying phylogenetic relationships and the evolutionary history of the bivalve order Arcida (Mollusca: Bivalvia: Pteriomorphia). Mol. Phylogenet Evol. 2016, 94, 298–312. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Ando, Y.; Shiba, T. Unusual priming mechanism of RNA-directed DNA synthesis in copia retrovirus-like particles of Drosophila. Nature 1986, 323, 824–826. [Google Scholar] [CrossRef]
- Ke, N.; Gao, X.; Keeney, J.B.; Boeke, J.D.; Voytas, D.F. The yeast retrotransposon Ty5 uses the anticodon stem-loop of the initiator methionine tRNA as a primer for reverse transcription. RNA 1999, 5, 929–938. [Google Scholar] [CrossRef]
- Neumann, P.; Novak, P.; Hostakova, N.; Macas, J. Systematic survey of plant LTR-retrotransposons elucidates phylogenetic relationships of their polyprotein domains and provides a reference for element classification. Mob. DNA 2019, 10, 1. [Google Scholar] [CrossRef]
- Han, G.; Zhang, N.; Jiang, H.; Meng, X.; Qian, K.; Zheng, Y.; Xu, J.; Wang, J. Diversity of short interspersed nuclear elements (SINEs) in lepidopteran insects and evidence of horizontal SINE transfer between baculovirus and lepidopteran hosts. BMC Genom. 2021, 22, 226. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Churakov, G.; Jurka, J.; Brosius, J.; Schmitz, J. Evolutionary history of 7SL RNA-derived SINEs in Supraprimates. Trends Genet. 2007, 23, 158–161. [Google Scholar] [CrossRef]
- de Souza, F.S.; Franchini, L.F.; Rubinstein, M. Exaptation of transposable elements into novel cis-regulatory elements: Is the evidence always strong? Mol. Biol. Evol. 2013, 30, 1239–1251. [Google Scholar] [CrossRef]
- Zhang, X.O.; Pratt, H.; Weng, Z. Investigating the Potential Roles of SINEs in the Human Genome. Annu. Rev. Genom. Hum. Genet. 2021, 22, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Cervera, A.; Urbina, D.; de la Pena, M. Retrozymes are a unique family of non-autonomous retrotransposons with hammerhead ribozymes that propagate in plants through circular RNAs. Genome Biol. 2016, 17, 135. [Google Scholar] [CrossRef]
Figure 1.
Schematic structures of autonomous LTR retrotransposons and TRIMs. LTRs are shown as triangles, while protein-coding regions are as gray rectangles. TSDs are shown as diamonds. PBSs are shown as black rectangles, and PPTs are shown as red rectangles. Small RNA-like sequences are colored in orange (5S rRNA) or in green (tRNA). Solo LTR is generated by the recombination between the two LTRs of an LTR retrotransposon. Some TRIMs are revealed to be generated by the deletion of a part of the internal portion of an autonomous LTR retrotransposon.
Figure 1.
Schematic structures of autonomous LTR retrotransposons and TRIMs. LTRs are shown as triangles, while protein-coding regions are as gray rectangles. TSDs are shown as diamonds. PBSs are shown as black rectangles, and PPTs are shown as red rectangles. Small RNA-like sequences are colored in orange (5S rRNA) or in green (tRNA). Solo LTR is generated by the recombination between the two LTRs of an LTR retrotransposon. Some TRIMs are revealed to be generated by the deletion of a part of the internal portion of an autonomous LTR retrotransposon.

Figure 2.
tRNA-derived sequences in Helenus LTRs. Boxes A and B for RNA polymerase III promoter are shown above the alignment with their motif sequences. The positions of the aligned sequence in the respective consensus is shown at both sides. (a) tRNA-derived sequences in representative fungal Helenus LTRs aligned with tRNA-Thr-CGT from Saccharomyces cerevisiae. (b) tRNA-derived sequences in representative hymenopteran Helenus LTRs aligned with tRNA-Lys-CTT from Apis mellifera. (c) tRNA-derived sequences in representative fish Helenus LTRs aligned with tRNA-Val-CAC from Homo sapiens. (d) tRNA-derived sequences in representative molluscan Helenus LTRs aligned with tRNA-Lys-TTT from Aplysia californica.
Figure 2.
tRNA-derived sequences in Helenus LTRs. Boxes A and B for RNA polymerase III promoter are shown above the alignment with their motif sequences. The positions of the aligned sequence in the respective consensus is shown at both sides. (a) tRNA-derived sequences in representative fungal Helenus LTRs aligned with tRNA-Thr-CGT from Saccharomyces cerevisiae. (b) tRNA-derived sequences in representative hymenopteran Helenus LTRs aligned with tRNA-Lys-CTT from Apis mellifera. (c) tRNA-derived sequences in representative fish Helenus LTRs aligned with tRNA-Val-CAC from Homo sapiens. (d) tRNA-derived sequences in representative molluscan Helenus LTRs aligned with tRNA-Lys-TTT from Aplysia californica.

Figure 3.
The evolution and PBS sequences of the fungal Helenus families. The phylogeny shown on the left is based on the tRNA-derived sequences in Helenus LTRs. The line and leaf color indicate the primer tRNA type. The bootstrap support value over 50% is shown at the branch of Helenus families not using 3′-truncated tRNA-iMet-CAT as a primer. PBS sequences are shown on the right. tRNA sequences are shown in red and in the reverse orientation. The anticodons are shown in bold. The site of intron is indicated by “|”. The PBS nucleotides complementary to the tRNA are highlighted in yellow.
Figure 3.
The evolution and PBS sequences of the fungal Helenus families. The phylogeny shown on the left is based on the tRNA-derived sequences in Helenus LTRs. The line and leaf color indicate the primer tRNA type. The bootstrap support value over 50% is shown at the branch of Helenus families not using 3′-truncated tRNA-iMet-CAT as a primer. PBS sequences are shown on the right. tRNA sequences are shown in red and in the reverse orientation. The anticodons are shown in bold. The site of intron is indicated by “|”. The PBS nucleotides complementary to the tRNA are highlighted in yellow.

Figure 4.
The alignment of Copia-9_PGr and LTR-1_PTrit. (A) Internal portion. (B) LTR. tRNA-like sequence is shown in bold.
Figure 4.
The alignment of Copia-9_PGr and LTR-1_PTrit. (A) Internal portion. (B) LTR. tRNA-like sequence is shown in bold.

Figure 5.
The sequence alignments of the 5S rRNA-like sequences in Ajax, Cassandra, and lepidopteran SINE3 along with 5S rRNA genes. Box A, IE and box C for RNA polymerase III promoter are shown above the alignment with their motif sequences. 5S rDNA sequences are from Oryza sativa (5S_OS), Homo sapiens (5S_HS), and Notospermus geniculatus (5S_NGe). Lepidopteran SINE3 are from Helicoverpa armigera (HaSE3), Lycaena phlaeas (SINE3-1_LycPhl), Plutella xylostella (PxSE4 and PxSE5), Chrysoteuchia culmella (SINE3-1_ChrCul and SINE3-2_ChrCul), Lerema accius (LaSE2), Ennomos fuscantarius (SINE3-1_EnnFus), and Chilo suppressalis (CsSE2). The positions of the aligned sequence in the respective consensus is shown at both sides.
Figure 5.
The sequence alignments of the 5S rRNA-like sequences in Ajax, Cassandra, and lepidopteran SINE3 along with 5S rRNA genes. Box A, IE and box C for RNA polymerase III promoter are shown above the alignment with their motif sequences. 5S rDNA sequences are from Oryza sativa (5S_OS), Homo sapiens (5S_HS), and Notospermus geniculatus (5S_NGe). Lepidopteran SINE3 are from Helicoverpa armigera (HaSE3), Lycaena phlaeas (SINE3-1_LycPhl), Plutella xylostella (PxSE4 and PxSE5), Chrysoteuchia culmella (SINE3-1_ChrCul and SINE3-2_ChrCul), Lerema accius (LaSE2), Ennomos fuscantarius (SINE3-1_EnnFus), and Chilo suppressalis (CsSE2). The positions of the aligned sequence in the respective consensus is shown at both sides.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The distribution of Helenus.
| Kingdom | Phylum | Class | Order | Family | Species |
|---|---|---|---|---|---|
| Fungi | Basidiomycota | Pucciniomycetes | Pucciniales | Pucciniaceae | Puccinia graminis, P. striiformis, P. triticina, P. hordei, P. horiana, P. coronata, P. novopanici, P. sorghi, P. arachidis, P. polysora, Uromyces transversalis, and U. viciae-fabae |
| Sphaerophragmiaceae | Austropuccinia psidii | ||||
| Melampsoraceae | Melampsora larici-populina, M. aecidioides, and M. medusae | ||||
| Coleosporiaceae | Cronartium ribicola | ||||
| Zaghouaniaceae | Hemileia vastatrix | ||||
| Agaricomycetes | Agaricales | Agaricaceae | Agaricus bisporus | ||
| Psathyrellaceae | Coprinopsis cinerea | ||||
| Metazoa | Ctenophora | Tentaculata | Lobata | Bolinopsidae | Mnemiopsis leidyi |
| Chordata | - | Coelacanthiformes | Coelacanthidae | Latimeria chalumnae | |
| Actinopteri | Acipenseriformes | Acipenseridae | Acipenser ruthenus | ||
| Polyodontidae | Polyodon spathula | ||||
| Cypriniformes | Danionidae | Danio rerio | |||
| Cyprinidae | Cyprinus carpio, Carassius auratus, Culter alburnus, Onychostoma macrolepis, Phoxinus phoxinus, and Squalius cephalus | ||||
| Nemacheilidae | Triplophysa dalaica | ||||
| Siluriformes | Bagridae | Hemibagrus wyckioides | |||
| Gonorynchiformes | Chanidae | Chanos chanos | |||
| Mollusca | Bivalvia | Ostreida | Ostreidae | Crassostrea gigas, C. virginica, and Saccostrea glomerata | |
| Pterioida | Pinnidae | Pinna nobilis | |||
| Pteriidae | Pinctada imbricata | ||||
| Mytilida | Mytilidae | Mytilus galloprovincialis, M. coruscus, Modiolus philippinarum, Gigantidas (Bathymodiolus) platifrons, and Limnoperna fortunei | |||
| Pectinida | Pectinidae | Argopecten irradians, Mizuhopecten yessoensis, and Pecten maximus | |||
| Arcoida | Arcidae | Tegillarca granosa | |||
| Brachiopoda | Lingulata | Lingulida | Lingulidae | Lingula anatina | |
| Nemertea | Pilidiophora | Heteronemertea | Lineidae | Lineus longissimus and Notospermus geniculatus | |
| Arthropoda | Insecta | Hymenoptera | Pteromalidae | Nasonia vitripennis, Cecidostiba fungosa, Trichomalopsis sarcophagae, Muscidifurax raptorellus, Philotrypesis tridentata, Theocolax elegans, Gastracanthus pulcherrimus, and Pteromalus puparum | |
| Eurytomidae | Eurytoma adleriae | ||||
| Eupelmidae | Eupelmus annulatus | ||||
| Megastigmidae | Megastigmus dorsalis | ||||
| Cynipidae | Synergus japonicus and S. umbraculus | ||||
| Torymidae | Torymus geranii | ||||
| Viridiplantae | Streptophyta | Lycopodiopsida | Selaginellales | Selaginellaceae | Selaginella moellendorffii |
Table 2.
Helenus and tRNA genes predicted in GtRNADB.
| Species | tRNA Type | Total | Helenus | Not Helenus |
|---|---|---|---|---|
| Puccinia triticina | tRNA-Thr-AGT | 492 | 481 | 11 |
| (LTR-1_PTrit: 426) | ||||
| (Helenus-8_PTrit: 55) | ||||
| tRNA-Ser-TGA | 189 | 183 | 6 | |
| (Helenus-2_PTrit: 95) | ||||
| (Helenus-11_PTrit: 40) | ||||
| (Helenus-11B_PTrit: 33) | ||||
| (Helenus-9_PTrit: 11) | ||||
| tRNA-Ala-TGC | 137 | 133 | 4 | |
| (Helenus-4_PTrit: 133) | ||||
| tRNA-Pro-TGG | 128 | 123 | 5 | |
| (Helenus-5_PTrit: 123) | ||||
| Puccinia striiformis | tRNA-Ala-AGC | 163 | 147 | 16 |
| (LTR-2_PSt: 145) | ||||
| tRNA-Thr-AGT | 49 | 32 | 17 | |
| (Helenus-4_PSt: 21) | ||||
| tRNA-Ser-AGA | 44 | 28 | 16 | |
| (Helenus-9_PSt: 28) | ||||
| Puccinia graminis | tRNA-Thr-AGT | 100 | 89 | 11 |
| (Helenus-3_PGr: 58) | ||||
| (Helenus-4_PGr: 15) | ||||
| Uromyces viciae-fabae | tRNA-Thr-AGT | 119 | 112 | 7 |
| (Helenus-1_UVF: 94) | ||||
| tRNA-Sup-CTA | 109 | 109 | 0 | |
| (Helenus-1_UVF: 105) | ||||
| tRNA-Ser-AGA | 95 | 90 | 5 | |
| (Helenus-2_UVF: 88) | ||||
| Cronartium ribicola | tRNA-Ala-AGC | 258 | 251 | 7 |
| (Helenus-4_CroRib: 249) | ||||
| tRNA-Sup-CTA | 245 | 245 | 0 | |
| (Helenus-5_CroRib: 194) | ||||
| (Helenus-3_CroRib: 51) | ||||
| tRNA-Thr-AGT | 160 | 154 | 6 | |
| (Helenus-2_CroRib: 88) | ||||
| (Helenus-4_CroRib: 29) | ||||
| (Helenus-5_CroRib: 22) | ||||
| tRNA-Ser-AGA | 160 | 154 | 6 | |
| (Helenus-6_CroRib: 118) | ||||
| (Helenus-13_CroRib: 13) | ||||
| tRNA-Val-GAC | 127 | 126 | 1 | |
| (Helenus-16_CroRib: 126) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kojima, K.K. Helenus and Ajax, Two Groups of Non-Autonomous LTR Retrotransposons, Represent a New Type of Small RNA Gene-Derived Mobile Elements. Biology 2024, 13, 119. https://doi.org/10.3390/biology13020119
AMA Style
Kojima KK. Helenus and Ajax, Two Groups of Non-Autonomous LTR Retrotransposons, Represent a New Type of Small RNA Gene-Derived Mobile Elements. Biology. 2024; 13(2):119. https://doi.org/10.3390/biology13020119
Chicago/Turabian StyleKojima, Kenji K. 2024. "Helenus and Ajax, Two Groups of Non-Autonomous LTR Retrotransposons, Represent a New Type of Small RNA Gene-Derived Mobile Elements" Biology 13, no. 2: 119. https://doi.org/10.3390/biology13020119
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.