Ocotea glomerata (Nees) Mez Extract and Fractions: Chemical Characterization, Anti-Candida Activity and Related Mechanism of Action

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the Ocotea glomerata Methanol Extract and the Fraction Components
2.2. Antifungal Susceptibility Testing
2.3. Checkerboard Analysis
2.4. Sorbitol and Ergosterol Test
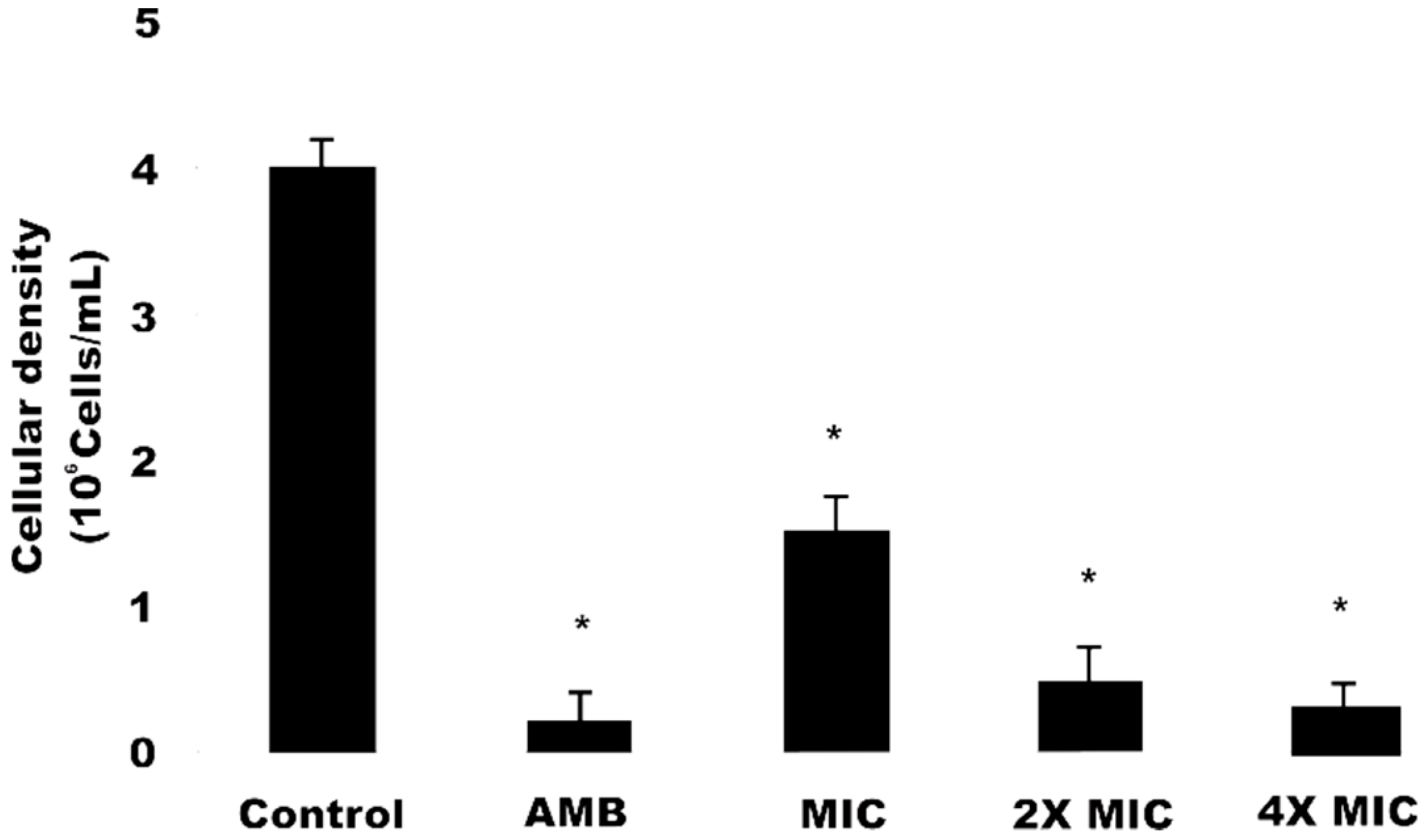
2.5. Cell Viability
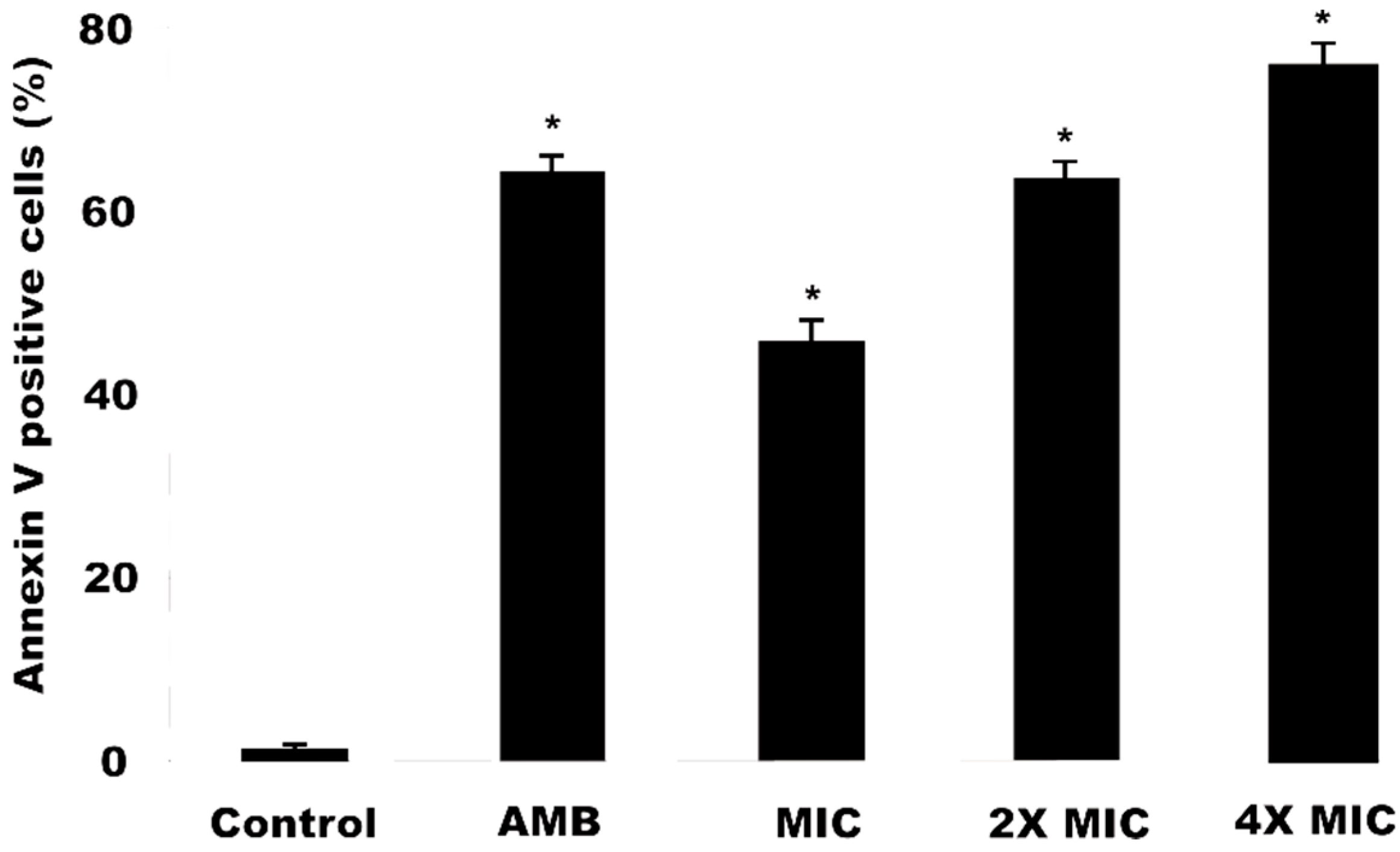
2.6. Detection of Phosphatidylserine (PS) in C. krusei Cells Induced by Methanol Extract
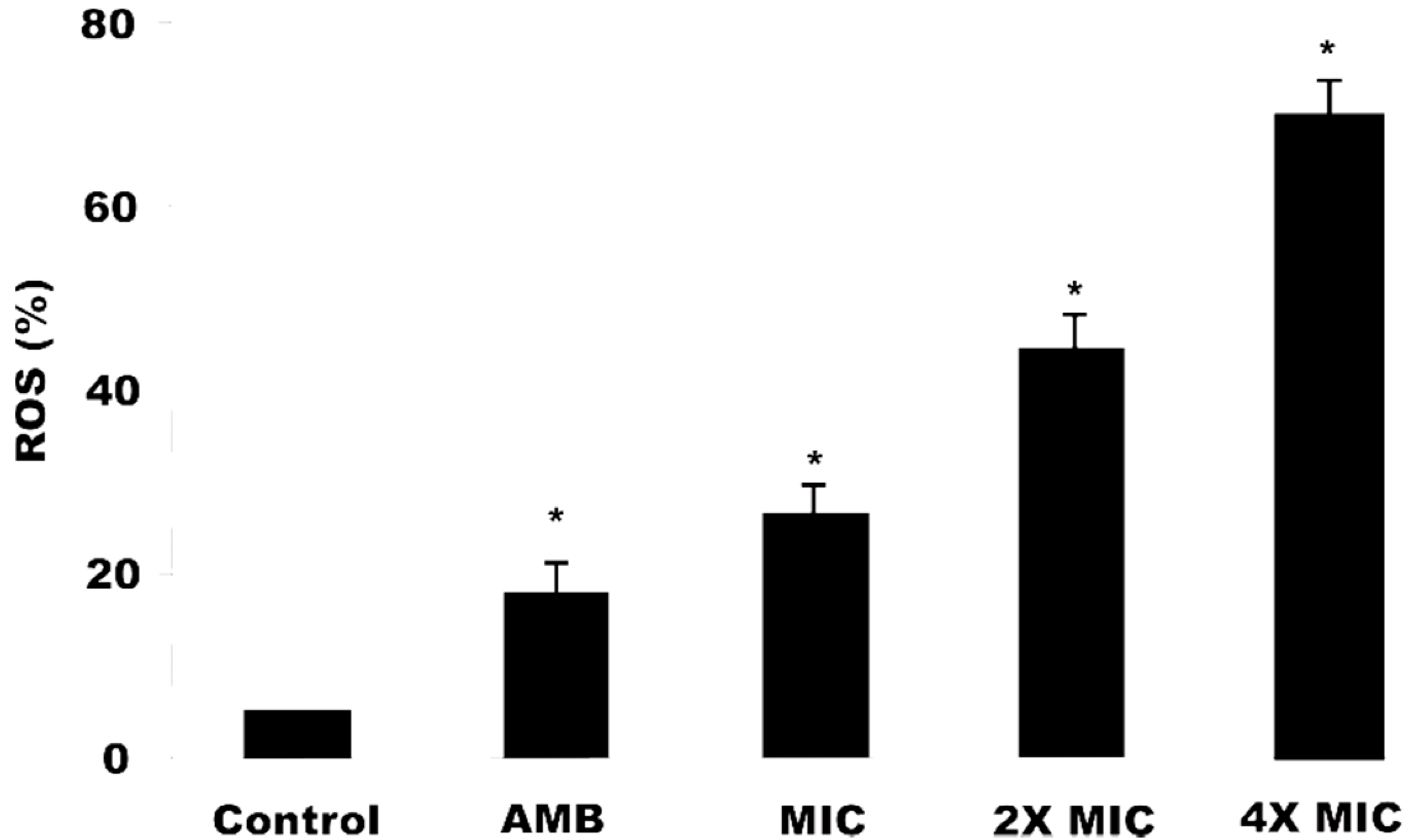
2.7. Reactive Oxygen Species (ERO) Production
3. Materials and Methods
3.1. Extract Acquisition
3.2. Methanol Extract Fractionation
3.3. Phytochemical Prospecting of Secondary Metabolites
3.4. Identification of Crude Methanolic Extract and Fraction Compounds by High-Performance Liquid Chromatography (HPLC)
3.5. Antifungal Activity
3.6. Checkerboard Analysis
3.7. Ergosterol Test
3.8. Sorbitol Test
3.9. Cell Viability Determination
3.10. Phosphatidylserine Externalization Determination
3.11. Reactive Oxygen Species Detection
3.12. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Paramythiotou, E.; Frantzeskaki, F.; Flevari, A.; Armaganidis, A.; Dimopoulos, G. Invasive Fungal Infections in the ICU: How to Approach, How to Treat. Molecules 2014, 19, 1085–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i4–i13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, G.C.O.; Vasconcelos, C.C.; Lopes, A.J.O.; Cartágenes, M.S.S.; Filho, A.K.D.B.; Nascimento, F.R.F.; Ramos, R.M.; Pires, E.R.R.B.; de Andrade, M.S.; Rocha, F.M.G.; et al. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front. Microbiol. 2018, 9, 1351–1373. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, F.; Orsetti, E.; Osimani, P.; Catassi, C.; Santelli, F.; Manso, E. Factors related to outcome of bloodstream infections due to Candida parapsilosis complex. BMC Infect. Dis. 2016, 16, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Shoham, S.; Marr, K.A. Invasive fungal infections in solid organ transplant recipients. Future Microbiol. 2012, 7, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical practice guideline for the management of candidiasis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [Green Version]
- Da, X.; Takahashi, H.; Hein, K.Z.; Morita, E. In vitro antifungal activity of Kampo medicine water extracts against Trichophyton rubrum. Nat. Prod. Commun. 2016, 2016, 763–766. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Saiki, I. Juzen-taiho-to (shi-quan-da-bu-tang): Scientific Evaluation and Clinical Applications. Evid. Based Complement. Alternat. Med. 2006, 3, 106–107. [Google Scholar]
- Butts, A.; Krysan, D.J. Antifungal drug discovery: Something old and something new. PLoS Pathog. 2012, 8, e1002870. [Google Scholar] [CrossRef]
- Cushnie, T.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.K.L.; Veiga-junior, V.F.; Pedrosa, T.N.; Vasconcellos, M.C.; Lima, E.S. Biological activities of essential oil of Endlicheria citriodora, a methyl geranate—Rich Lauraceae. Quim. Nova 2013, 36, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Olivero-verbel, J.; Cervera, G.T.; Fernandez, G.J.; Colorado, J.B.; Stashenko, E. Chemical composition and antioxidant activity of essential oils isolated from Colombian plants. Rev. Bras. Farmacogn. 2010, 20, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Oltramari, A.C.; Wood, K.V.; Bonham, C.; Verpoorte, R.; Caro, M.S.B.; Viana, A.M.; Pedrotti, E.L.; Maraschin, R.P.; Maraschin, M. Safrole analysis by GC-MS of prototrophic (Ocotea odorifera (Vell.) Rohwer) cell cultures. Plant Cell Tissue Organ Cult. 2004, 78, 231–235. [Google Scholar] [CrossRef]
- Leporatti, M.L.; Pintore, G.; Foddai, M.; Chessa, M.; Piana, A.; Patretto, G.L.; Masia, M.D.; Mangano, G.; Nicoletti, M. Chemical, biological, morphoanatomical and antimicrobial study of Ocotea puchury-major Mart. Nat. Prod. Res. 2014, 28, 294–300. [Google Scholar] [CrossRef]
- Farago, P.V.; De Paula, J.F.P.; Nakashima, T.; Döllboscardin, P.M.; Budel, J.M.; Maia, B.H.L.N.S. Chemical composition and antibacterial activity of the essential oil from bark of Ocotea puberula (Rich.) Ness. Lat. Am. J. Pharm. 2010, 29, 1242–1245. [Google Scholar]
- Orhan, D.D.; Ozeçelik, B.; Ozgen, S.; Ergun, F. Antibacterial, antifungal, and antiviral activities of some flavonoids. Microbiol. Res. 2010, 165, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Savoia, D. Plant-derived antimicrobial compounds: Alternatives to antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Approved Standard-Third Edition; CLSI document M27-S4; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Ben-Ami, R.; Olshtain-Pops, K.; Krieger, M.; Oren, I.; Bishara, J.; Dan, M.; Wiener-Well, Y.; Weinberger, M.; Zimhony, O.; Chowers, M.; et al. Antibiotic exposure as a risk factor for fluconazole-resistant Candida bloodstream infection. Antimicrob. Agents Chemother. 2012, 56, 2518–2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural products for human health: An historical overview of the drug discovery approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef] [PubMed]
- Roling, E.E.; Klepser, M.E.; Wasson, A.; Lewis, R.E.; Ernst, E.J.; Pfaller, M.A. Antifungal activities of fluconazole, caspofungin (MK0991), and anidulafungin (LY 303366) alone and in combination against Candida spp. and Crytococcus neoformans via time-kill methods. Diagn. Microbiol. Infect. Dis. 2002, 43, 13–17. [Google Scholar] [CrossRef]
- Li, H.; Zhang, C.; Chen, Z.; Shi, W.; Sun, S. A promising approach of overcoming the intrinsic resistance of Candida krusei to fluconazole (FLC)-combining tacrolimus with FLC. FEMS Yeast Res. 2014, 14, 808–811. [Google Scholar] [CrossRef] [Green Version]
- Aligiannis, N.; Kalpotzakis, E.; Mitaku, S.; Chinou, I.B. Composition and antimicrobial activity of the essential oil of two Origanum species. J. Agric. Food Chem. 2001, 40, 4168–4170. [Google Scholar] [CrossRef]
- Webster, D.; Taschereau, P.; Belland, R.J.; Sand, C.; Rennie, R.P. Antifungal activity of medicinal plant extracts; preliminary screening studies. J. Ethnopharmacol. 2008, 115, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Diekema, D.J.; Messer, S.A.; Pfaller, M.A.; Klepser, M.E. Comparasion of E-teste, chequerboard dilution and time-kill studies for the detection or synergy or antagonism between antifungal agents tested against Candida species. J. Antimicrob. Chemother. 2002, 49, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, D.J.; Brandt, K.D.; Cugier, D.; Goldman, R. A whole-cell Candida albicans assay for the detection of inhibitors towards fungal cell wall synthesis and assembly. J. Antibiot. 1995, 48, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Lupetti, A.; Welling, M.M.; Mazzi, U.; Nibbering, P.H.; Pauwels, E.K. Technetium-99m labelled fluconazole and antimicrobial peptides for imaging of Candida albicans and Aspergillus fumigatus infection. Eur. J. Nucl. Med. Mol. Imaging 2002, 29, 674–679. [Google Scholar] [CrossRef]
- Escalante, M.; Gattuso, P.; Pérez, S.; Zacchino, S. Evidence for the mechanism of action of the antifungal phytolaccoside B isolated from Phytolacca tetramera Hauman. J. Nat. Prod. 2008, 71, 1720–1725. [Google Scholar] [CrossRef]
- Da Silva, C.R.; Neto, J.B.A.; Sidrim, J.J.C.; Magalhães, H.I.; Cavalcanti, B.C.; Brilhante, R.S.; Macedo, D.S.; de Moraes, M.O.; Lobo, M.D.; Grangeiro, T.B.; et al. Synergistic effects of amiodarone and fluconazole on Candida tropicalis resistant to fluconazole. Antimicrob. Agents Chemother. 2013, 57, 1691–1700. [Google Scholar] [CrossRef] [Green Version]
- Sandes, J.M.; Fontes, A.; Regis-da-Silva, C.G.; Castro, M.C.A.B.; Lima-Junior, C.G.; Silva, F.P.L.; Vasconcellos, M.L.A.A.; Figueiredo, R.C.B.Q. Trypanosoma cruzi Cell Death Induced by the Morita-Baylis-Hillman Adduct 3-Hydroxy-2-Methylene-3 (4-Nitrophenylpropanenitrile). PLoS ONE 2014, 9, e93936. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, J.; Gao, Y.; Lin, H.; Du, L.; Yang, S.; Long, S.; She, Z.; Cai, X.; Zhou, S.; et al. The anthracenedione compound bostrycin induces mitochondria-mediated apoptosis in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.H.; Choi, H.; Kim, A.R.; Yun, J.W.; Wo, E.R.; Lee, D.G. Hibicuslide C-induced cell death in Candida albicans involves apoptosis mechanism. J. Appl. Microbiol. 2014, 117, 1400–1411. [Google Scholar] [CrossRef]
- Ferreira, J.R.; Cavalcanti, B.C.; Da Costa, P.M.; De Arantes, F.F.P.; Alvarenga, E.S. Induction of G2/M arrest, caspase activation and apoptosis by α-santonin derivatives in HL-60 cells. Toxicol. In Vitro 2013, 27, 1458–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, H.; Zhao, L.; Li, C.; Shang, Q.; Lu, H.; Fu, Z.; Wang, L.; Jiang, Y.; Cao, Y. Inhibitory effect of Shikonin on Candida albicans growth. Biol. Pharm. Bull. 2012, 35, 1956–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Lee, D.G. Lycopene induces apoptosis in Candida albicans through reactive oxygen species production and mitochondrial dysfunction. Biochimie 2015, 115, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Tveden-Nyborg, P.; Bergmann, T.K.; Lykkesfeldt, J. Basic & Clinical Pharmacology & Toxicology Policy for Experimental and Clinical studies. Basic Clin. Pharmacol. Toxicol. 2018, 123, 233–235. [Google Scholar]
- Matos, F.J.A. Introdução à Fitoquímica Experimental, 3rd ed.; Editora da Universidade Federal do Ceará: Fortaleza, CE, Brazil, 2009. [Google Scholar]
- Guerra, F.Q.S.; Mendes, J.M.; Sousa, J.P.D. Increasing antibiotic activity against a multidrug-resistant Acinetobacter spp. by essential oils of Citrus limon and Cinnamomum zeylanicum. Nat. Prod. Res. 2012, 26, 2235–2238. [Google Scholar] [CrossRef]
- Leite, M.C.; de Brito Bezerra, A.P.; de Sousa, J.P.; de Oliveira Lima, E. Investigating the antifungal activity and mechanism(s) of geraniol against Candida albicans strains. Med. Mycol. 2015, 53, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Neto, J.B.; Da Silva, C.R.; Neta, M.A.S.; Campos, R.S.; Siebra, J.T.; Silva, R.A.; Gaspar, D.M.; Magalhães, H.I.; de Moraes, M.O.; Lobo, M.D.; et al. Antifungal activity of naphthoquinoidal compounds in vitro against fluconazole resistant strains of different Candida species: A special emphasis on mechanisms of action on Candida tropicalis. PLoS ONE 2014, 9, e93698. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Cinnamic Derivatives | Flavonoids | ||||||
|---|---|---|---|---|---|---|---|
| OGME | 6.600 | 25.377; | 27.223 | ||||
| FEM1 | 5.650; | 6.443 | 25.293 | ||||
| FEM2 | 6.543; | 9.860; | 15.150 | 10.117; | 25.263 | ||
| FEM3 | 10.070 | 25.263 | |||||
| FEM4 | - | 25.267; | 25.637; | 26.127; | 26.277 | ||
| FEM5 | - | 25.263; | 27.177 | ||||
| Species/Compounds | C. albicans | C. parapsilosis | C. tropicalis | C. krusei | |||
|---|---|---|---|---|---|---|---|
| ATCC14053 | URM5901 | ATCC22019 | URM4970 | URM7048 | URM6551 | ATCC6258 | |
| OGHE | - | - | - | - | 400 | 100 | - |
| OGCE | - | 400 | 200 | 100 | 100 | - | 100 |
| OGAcOEt | - | 200 | - | 400 | 100 | - | 100 |
| OGME | 400 | 6.25 | 6.25 | 12.5 | 6.25 | - | 3.12 |
| FEM1 | - | - | 400 | 400 | 400 | 200 | 200 |
| FEM2 | - | - | - | - | - | - | - |
| FEM3 | - | - | - | - | - | - | - |
| FEM4 | - | - | - | - | - | - | - |
| FEM5 | 400 | 400 | 400 | 400 | 50 | 200 | 25 |
| FLC | - | 16 | 1 | 8 | 4 | 1 | NA |
| KET | 0.06 | - | 0.06 | 0.06 | 8 | 0.06 | 2 |
| AMB | 0.06 | 0.25 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| Species/Compounds | C. albicans ATCC14053 | C. krusei ATCC6258 | C. parapsilosis ATCC22019 | |||
|---|---|---|---|---|---|---|
| MIC | FIC Index | MIC | FIC Index | CIM | FIC Index | |
| OGME | 400 | 3.12 | 6.25 | |||
| AMB | 0.06 | 0.06 | 0.06 | |||
| KET | 0.06 | 2 | 0.06 | |||
| FLC | 16 | NA | NA | 1 | ||
| OGME/AMB | 12.5/0.06 | 1.03 (I) | 1.56/0.03 | 1 (I) | 3.12/0.06 | 1.5 (I) |
| OGME/ KET | 6.25/0.03 | 0.51 (A) | 0.78/0.03 | 0.26 (S) | 0.78/0.03 | 0.62 (A) |
| OGME/FLC | 6.25/0.12 | 0.02 (S) | NA | NA | 3.12/0.06 | 0.56 (A) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anjos, M.N.V.; Araújo-Neto, L.N.d.; Silva Buonafina, M.D.; Pereira Neves, R.; Souza, E.R.d.; Bezerra, I.C.F.; Ferreira, M.R.A.; Soares, L.A.L.; Coutinho, H.D.M.; Martins, N.; et al. Ocotea glomerata (Nees) Mez Extract and Fractions: Chemical Characterization, Anti-Candida Activity and Related Mechanism of Action. Antibiotics 2020, 9, 394. https://doi.org/10.3390/antibiotics9070394
Anjos MNV, Araújo-Neto LNd, Silva Buonafina MD, Pereira Neves R, Souza ERd, Bezerra ICF, Ferreira MRA, Soares LAL, Coutinho HDM, Martins N, et al. Ocotea glomerata (Nees) Mez Extract and Fractions: Chemical Characterization, Anti-Candida Activity and Related Mechanism of Action. Antibiotics. 2020; 9(7):394. https://doi.org/10.3390/antibiotics9070394
Chicago/Turabian StyleAnjos, Mayara Nunes Vitor, Luiz Nascimento de Araújo-Neto, Maria Daniela Silva Buonafina, Rejane Pereira Neves, Edson Rubhens de Souza, Isabelle Cristinne Ferraz Bezerra, Magda Rhayanny Assunção Ferreira, Luiz Alberto Lira Soares, Henrique Douglas Melo Coutinho, Natália Martins, and et al. 2020. "Ocotea glomerata (Nees) Mez Extract and Fractions: Chemical Characterization, Anti-Candida Activity and Related Mechanism of Action" Antibiotics 9, no. 7: 394. https://doi.org/10.3390/antibiotics9070394