

Artemisinin May Disrupt Hyphae Formation by Suppressing Biofilm-Related Genes of Candida albicans: In Vitro and In Silico Approaches
 , ,
, ,
Abstract
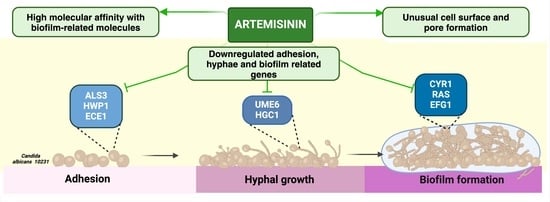
:
1. Introduction
2. Results
2.1. Artemisinin Exhibits Antifungal Activity on Candida spp.
2.2. Artemisinin Inhibits Biofilm Formation of C. albicans
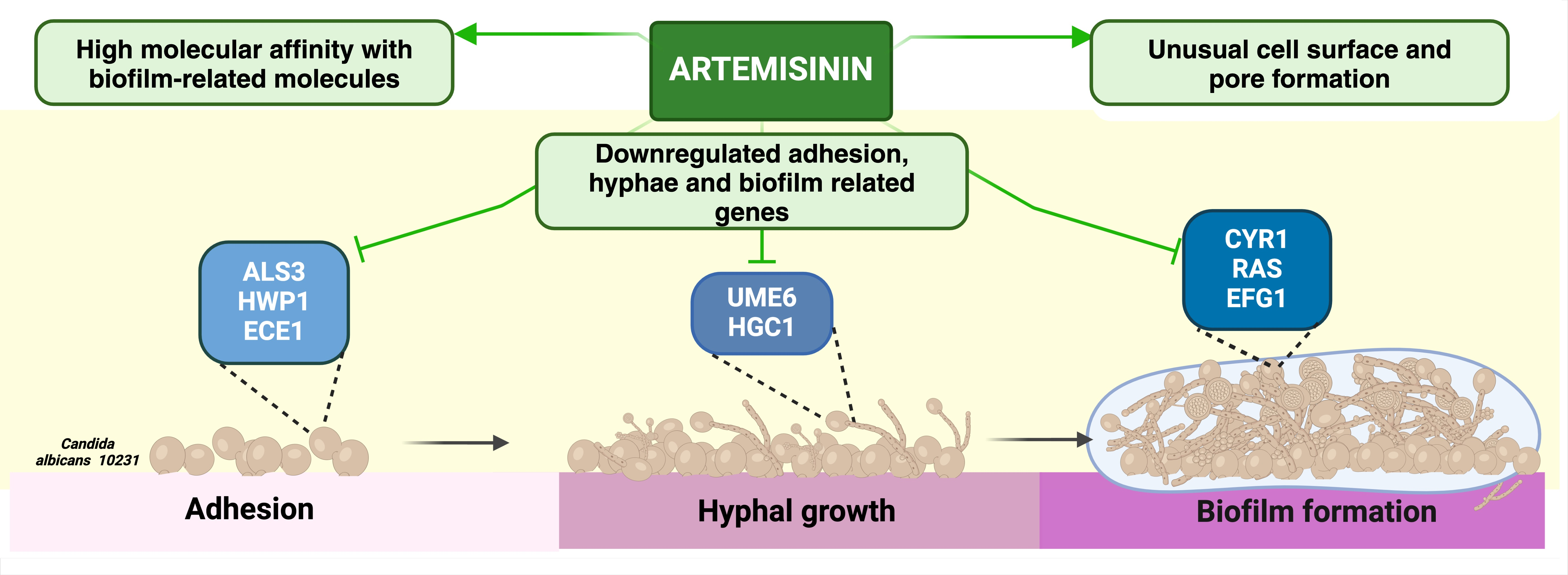
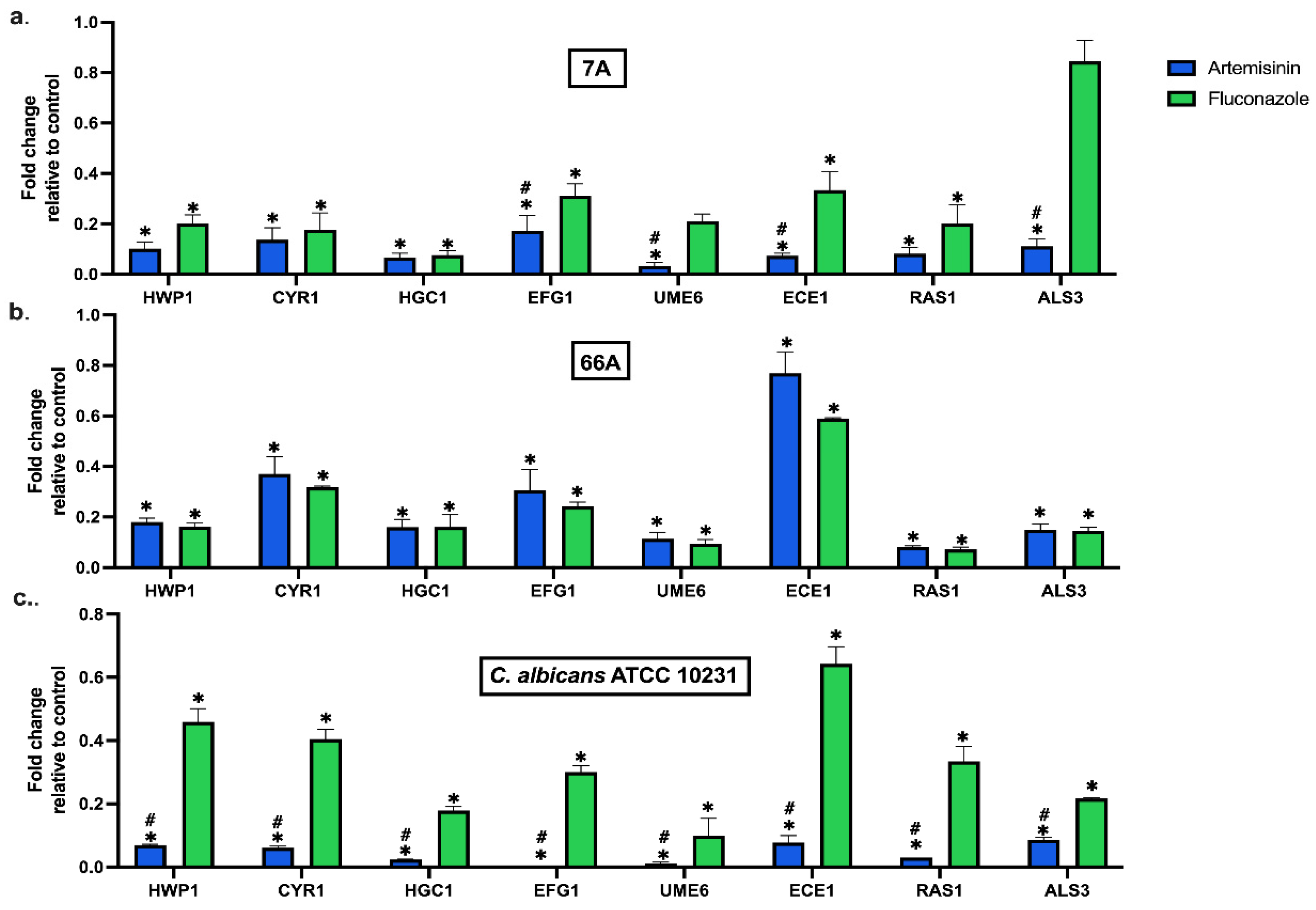
2.3. Artemisinin Affects Hyphae, Adhesion, and Biofilm-Related Gene Expression in C. albicans
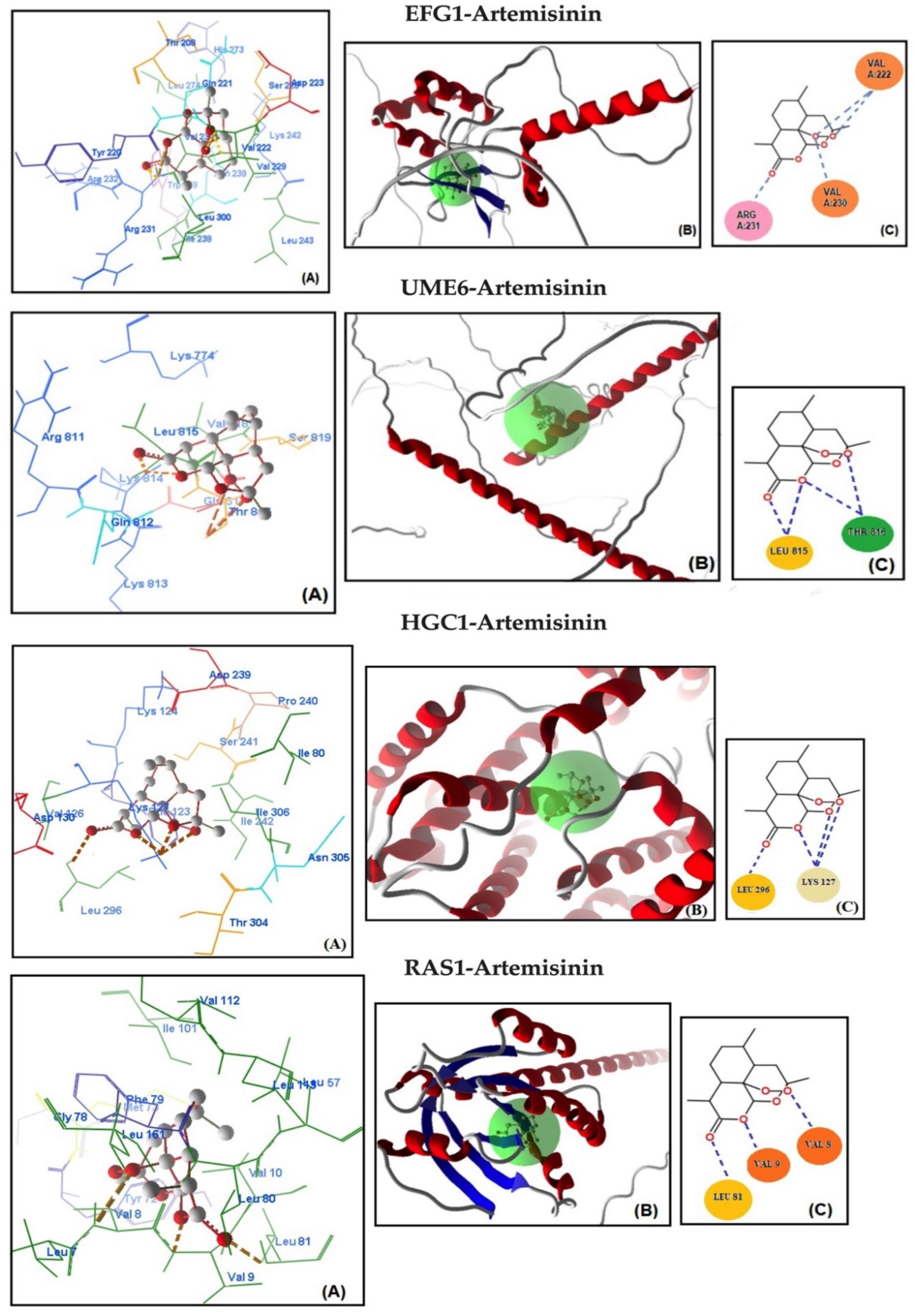
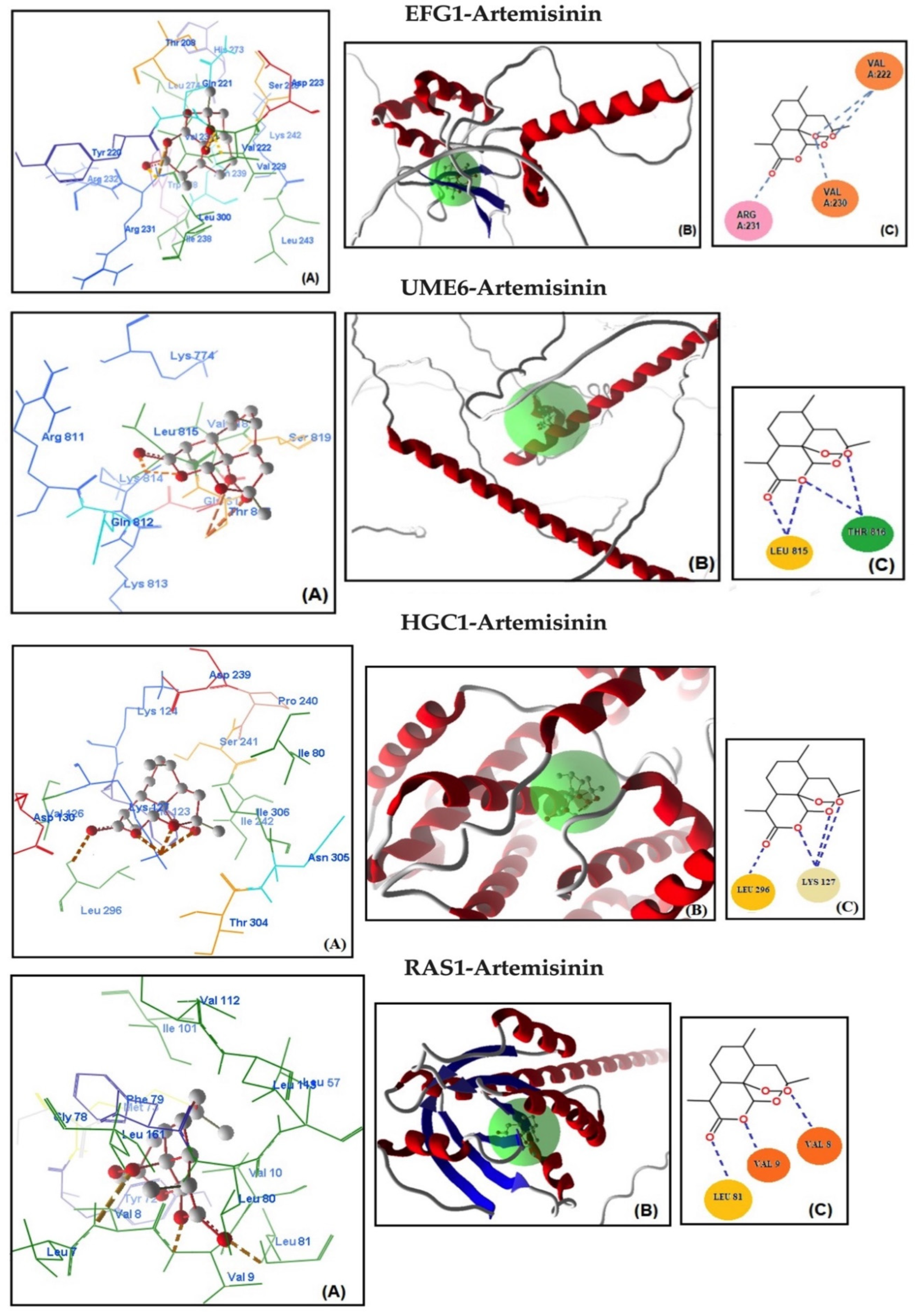
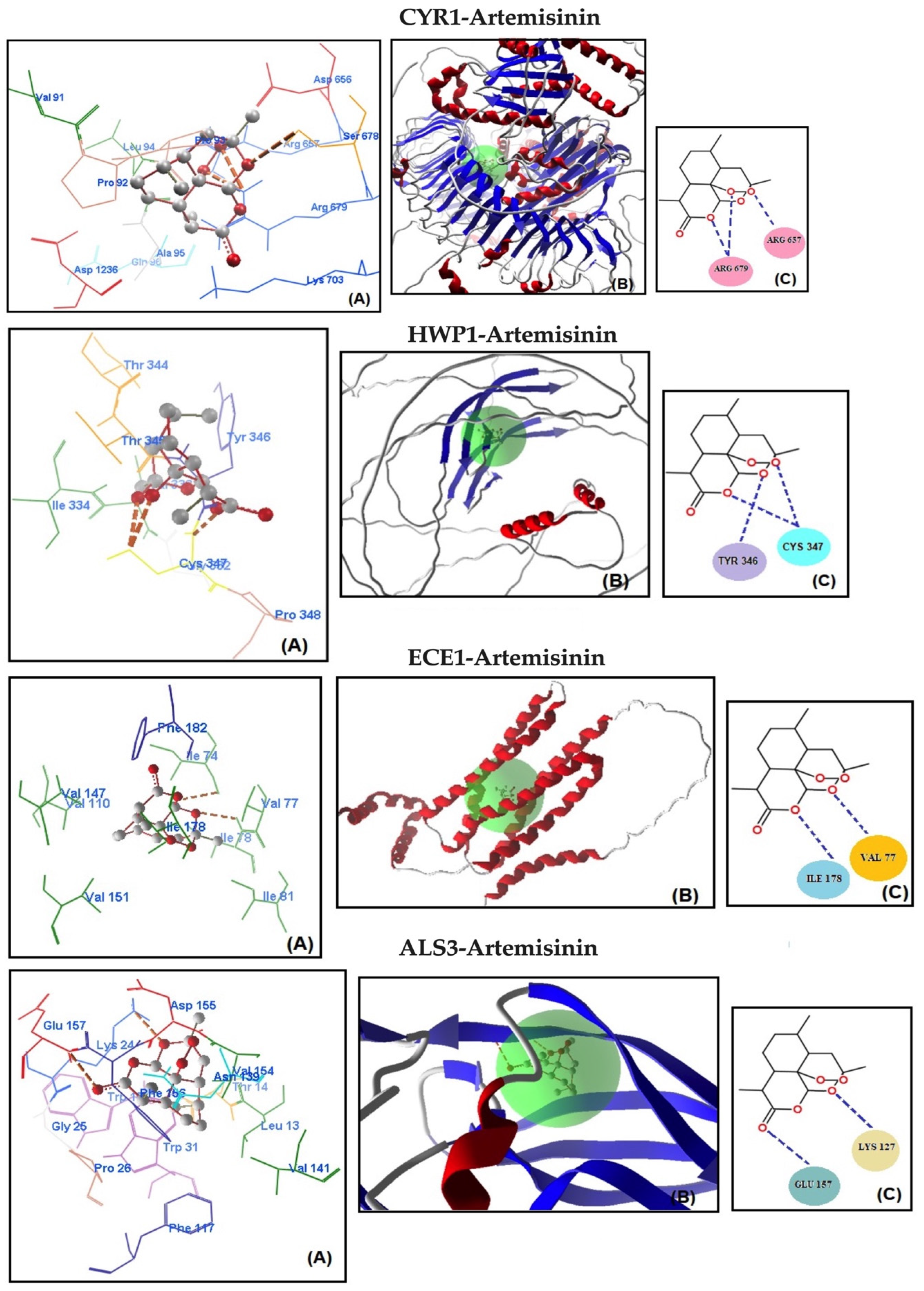
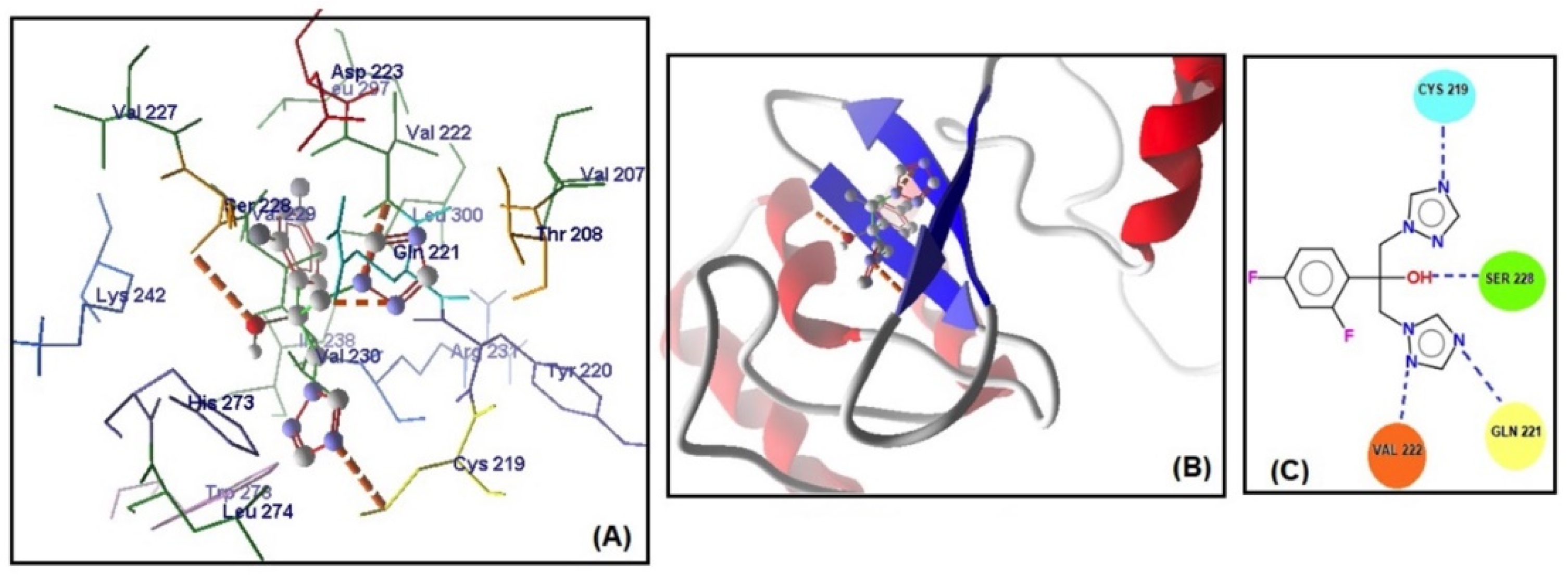
2.4. Molecular Docking Studies
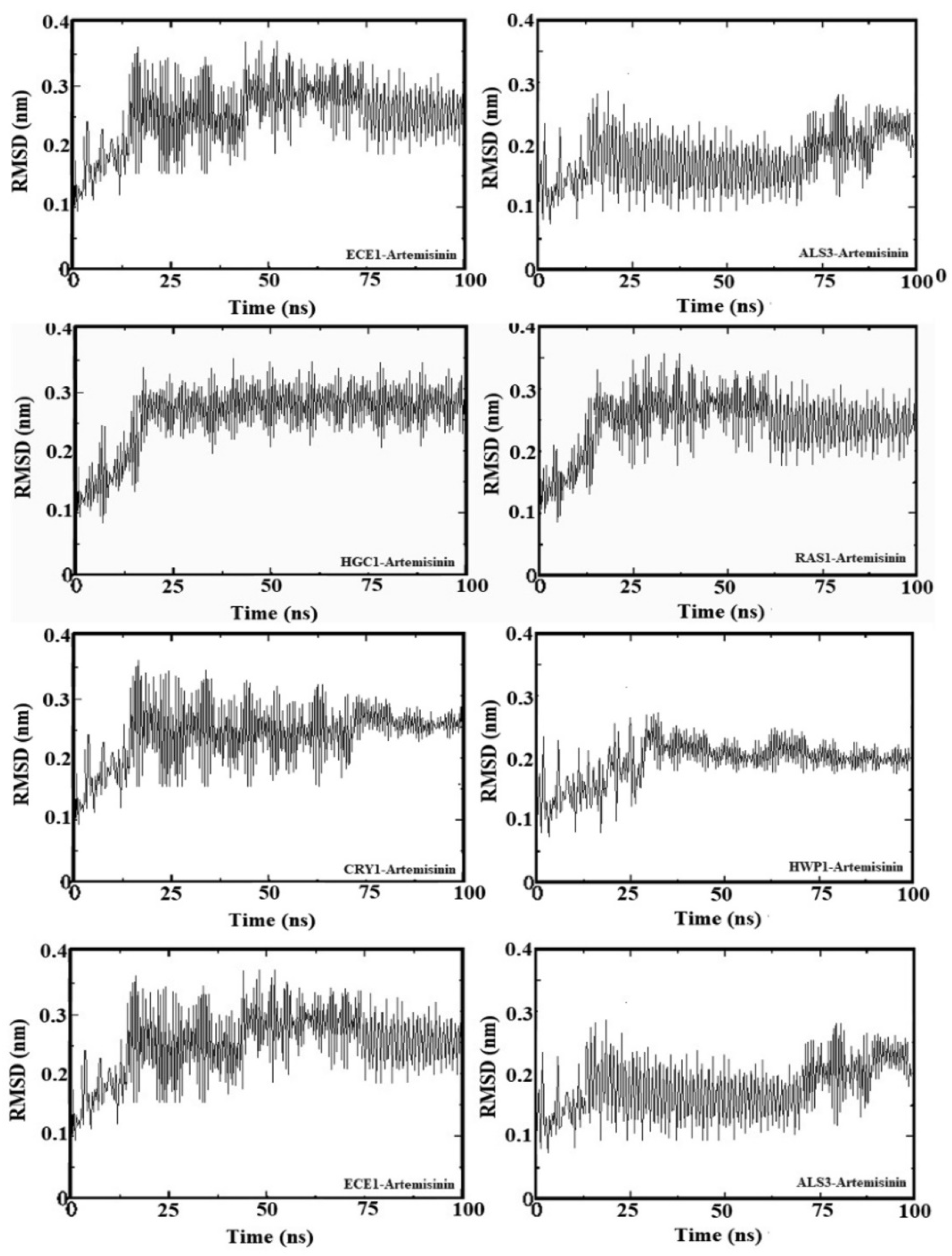
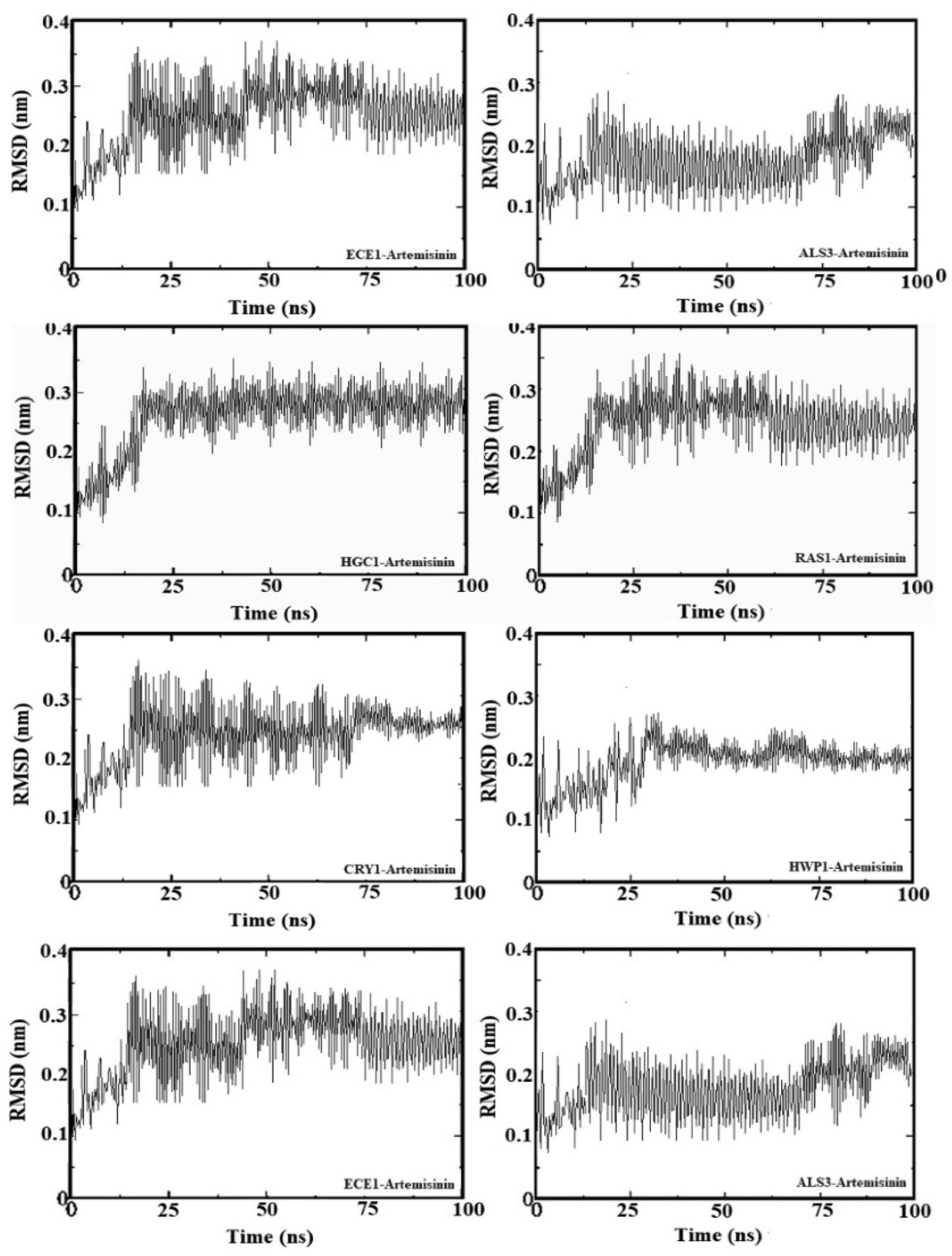
2.5. Molecular Dynamic Simulation
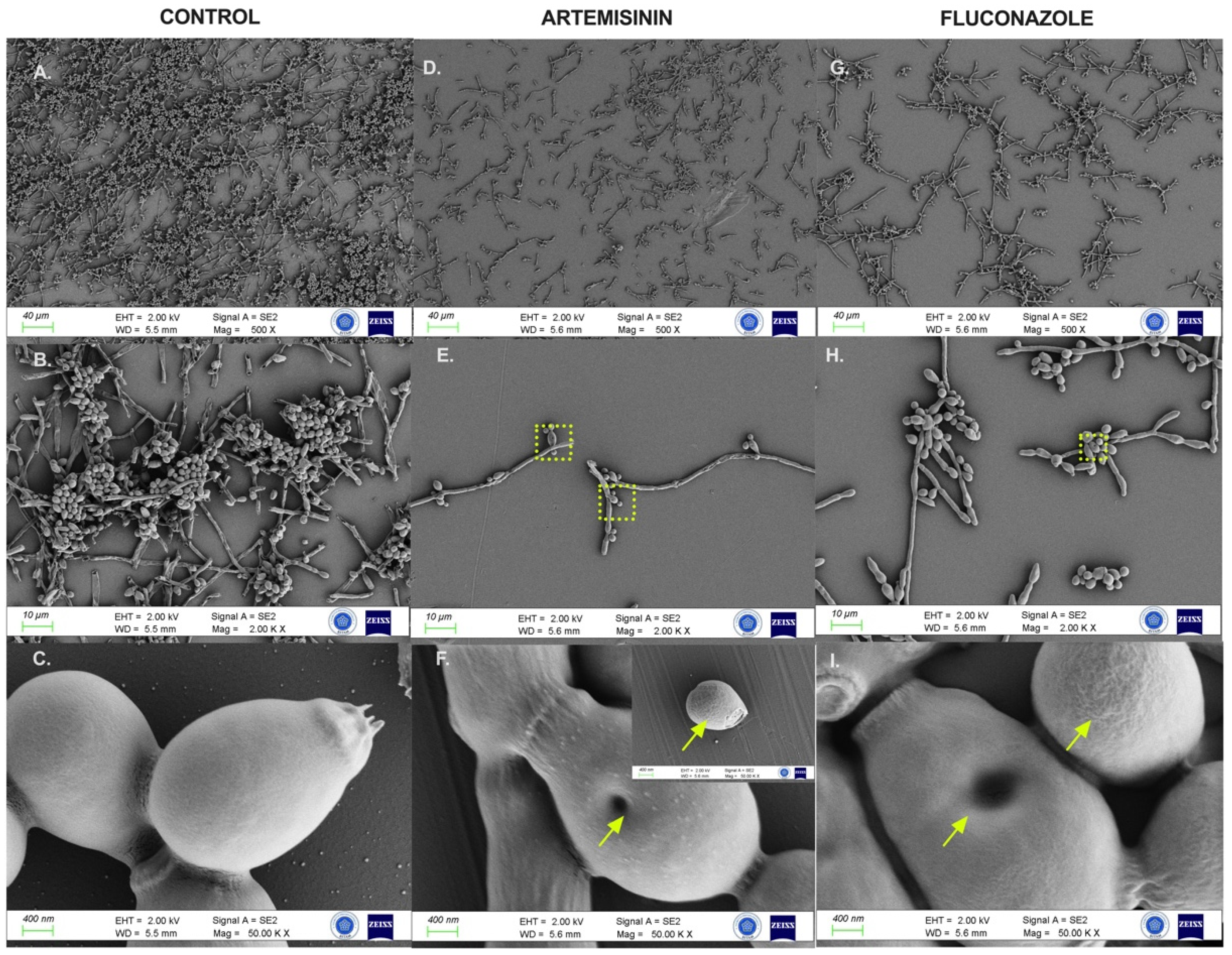
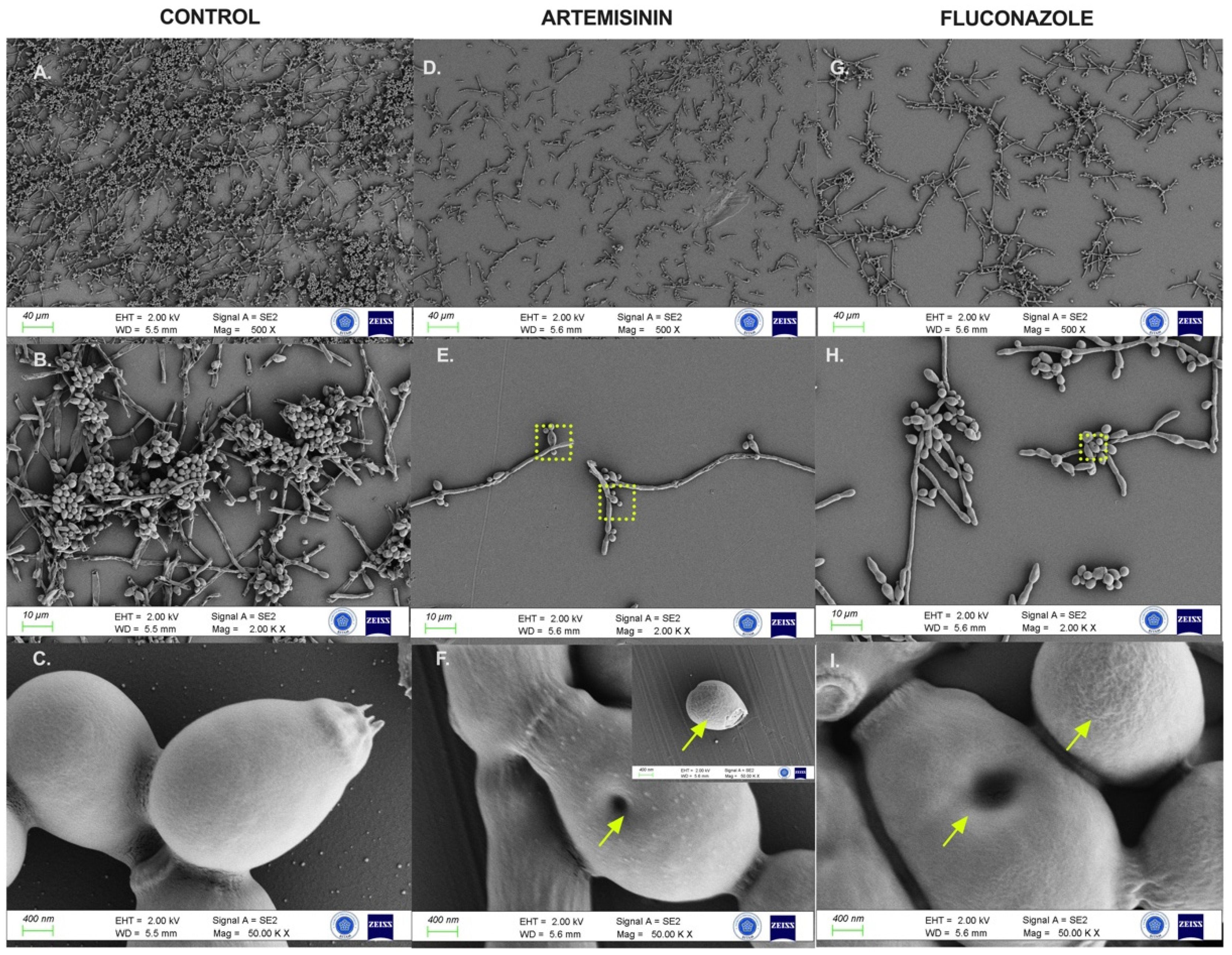
2.6. Artemisinin Impaired Cytopathological Damage to C. albicans Biofilms
3. Discussion
Limitations
4. Material and Methods
4.1. Candida Strains
4.2. Antifungal Susceptibility Testing
4.3. In Vitro Biofilm Formation Assay
4.4. Biofilm Formation and Treatment
4.5. Determination of the Biofilm and Hypha-Specific Gene Expressions by Real-Time Polymerase Chain Reaction
4.6. Molecular Docking Studies
4.7. Molecular Dynamics Simulation Studies
4.8. Field Emission Scanning Electron Microscopy
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans-The Virulence Factors and Clinical Manifestations of Infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef]
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and virulence of Candida albicans. Virulence 2022, 13, 89–121. [Google Scholar] [CrossRef]
- Ciurea, C.N.; Kosovski, I.-B.; Mare, A.D.; Toma, F.; Pintea-Simon, I.A.; Man, A. Candida and candidiasis-opportunism versus pathogenicity: A review of the virulence traits. Microorganisms 2020, 8, 857. [Google Scholar] [CrossRef]
- Malinovská, Z.; Čonková, E.; Váczi, P. Biofilm Formation in Medically Important Candida Species. J. Fungi 2023, 9, 955. [Google Scholar] [CrossRef]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. Candida albicans biofilms and polymicrobial interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef]
- Alvendal, C.; Mohanty, S.; Bohm-Starke, N.; Brauner, A. Anti-biofilm activity of chlorhexidine digluconate against Candida albicans vaginal isolates. PLoS ONE 2020, 15, e0238428. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Craik, C.S.; Johnson, A.D.; Nobile, C.J. Combination of Antifungal Drugs and Protease Inhibitors Prevent Candida albicans Biofilm Formation and Disrupt Mature Biofilms. Front. Microbiol. 2020, 11, 1027. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The regulation of hyphae growth in Candida albicans. Virulence 2020, 11, 337–348. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, P.; Li, Y.; Gong, Y.; Chen, X.; Sun, S. The Synergistic Antifungal Effect and Potential Mechanism of D-Penicillamine Combined With Fluconazole against Candida albicans. Front. Microbiol. 2019, 10, 2853. [Google Scholar] [CrossRef]
- Perfect, J.R.; Ghannoum, M. Emerging Issues in Antifungal Resistance. Infect. Dis. Clin. N. Am. 2020, 34, 921–943. [Google Scholar] [CrossRef] [PubMed]
- Ajetunmobi, O.H.; Badali, H.; Romo, J.A.; Ramage, G.; Lopez-Ribot, J.L. Antifungal therapy of Candida biofilms: Past, present and future. Biofilm 2023, 5, 100126. [Google Scholar] [CrossRef] [PubMed]
- Moraes, D.C.; Ferreira-Pereira, A. Insights on the anticandidal activity of non-antifungal drugs. J. Mycol. Med. 2019, 29, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Shen, B. A New Golden Age of Natural Products Drug Discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, Y.; Li, Y.; Zhang, X.; Wong, Y.-K.; Shen, S.; Zhong, T.; Zhang, J.; Liu, Q.; Wang, J. Advances in the research on the targets of anti-malaria actions of artemisinin. Pharmacol. Ther. 2020, 216, 107697. [Google Scholar] [CrossRef]
- Nabi, N.; Singh, S.; Saffeullah, P. An updated review on distribution, biosynthesis and pharmacological effects of artemisinin: A wonder drug. Phytochemistry 2023, 214, 113798. [Google Scholar] [CrossRef]
- Galal, A.M.; Ross, S.A.; Jacob, M.; ElSohly, M.A. Antifungal activity of artemisinin derivatives. J. Nat. Prod. 2005, 68, 1274–1276. [Google Scholar] [CrossRef]
- De Cremer, K.; Lanckacker, E.; Cools, T.L.; Bax, M.; De Brucker, K.; Cos, P.; Cammue, B.P.A.; Thevissen, K. Artemisinins, new miconazole potentiators resulting in increased activity against Candida albicans biofilms. Antimicrob. Agents Chemother. 2015, 59, 421–426. [Google Scholar] [CrossRef]
- Das, S.; Czuni, L.; Báló, V.; Papp, G.; Gazdag, Z.; Papp, N.; Kőszegi, T. Cytotoxic Action of Artemisinin and Scopoletin on Planktonic Forms and on Biofilms of Candida Species. Molecules 2020, 25, 476. [Google Scholar] [CrossRef]
- Elfawal, M.A.; Gray, O.; Dickson-Burke, C.; Weathers, P.J.; Rich, S.M. Artemisia annua and artemisinins are ineffective against human Babesia microti and six Candida sp. Longhua Chin. Med. 2021, 4, 12. [Google Scholar] [CrossRef]
- Li, H.; Chen, H.; Shi, W.; Shi, J.; Yuan, J.; Duan, C.; Fan, Q.; Liu, Y. A novel use for an old drug: Resistance reversal in Candida albicans by combining dihydroartemisinin with fluconazole. Future Microbiol. 2021, 16, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, J.; Cheong, I.; Liu, N.-N.; Wang, H. Evaluation of artemisinin derivative artemether as a fluconazole potentiator through inhibition of Pdr5. Bioorg. Med. Chem. 2021, 44, 116293. [Google Scholar] [CrossRef]
- Zhu, C.; Liao, B.; Ye, X.; Zhou, Y.; Chen, X.; Liao, M.; Cheng, L.; Zhou, X.; Ren, B. Artemisinin elevates ergosterol levels of Candida albicans to synergise with amphotericin B against oral candidiasis. Int. J. Antimicrob. Agents 2021, 58, 106394. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Yue, C.; Zeng, X.; Chen, X. Artemisinin Targets Transcription Factor PDR1 and Impairs Candida glabrata Mitochondrial Function. Antioxidants 2022, 11, 1855. [Google Scholar] [CrossRef]
- Liang, X.; Chen, D.; Wang, J.; Liao, B.; Shen, J.; Ye, X.; Wang, Z.; Zhu, C.; Gou, L.; Zhou Xinxuan Cheng, L.; et al. Artemisinins inhibit oral candidiasis caused by Candida albicans through the repression on its hyphal development. Int. J. Oral Sci. 2023, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Mo, W.; Shen, D.; Sun, L.; Wang, J.; Lu, S.; Gitschier, J.M.; Zhou, B. Yeast model uncovers dual roles of mitochondria in action of artemisinin. PLoS Gene. 2005, 1, e36. [Google Scholar] [CrossRef]
- Wang, J.; Xu, C.; Wong, Y.K.; Li, Y.; Liao, F.; Jiang, T.; Tu, Y. Artemisinin, the magic drug discovered from traditional Chinese medicine. Engineering 2019, 5, 32–39. [Google Scholar] [CrossRef]
- Dighal, A.; De Sarkar, S.; Gille, L.; Chatterjee, M. Can the iron content of culture media impact on the leishmanicidal effect of artemisinin? Free Radic. Res. 2021, 55, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Wilson, D.; Hube, B. Candida albicans iron acquisition within the host. FEMS Yeast Res. 2009, 9, 1000–1012. [Google Scholar] [CrossRef]
- Hameed, S.; Prasad, T.; Banerjee, D.; Chandra, A.; Mukhopadhyay, C.K.; Goswami, S.K.; Lattif, A.A.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A.; et al. Iron deprivation induces EFG1-mediated hyphal development in Candida albicans without affecting biofilm formation. FEMS Yeast Res. 2008, 8, 744–755. [Google Scholar] [CrossRef]
- Savage, K.A.; Parquet, M.C.; Allan, D.S.; Davidson, R.J.; Holbein, B.E.; Lilly, E.A.; Fidel, P.L., Jr. Iron Restriction to Clinical Isolates of Candida albicans by the Novel Chelator DIBI Inhibits Growth and Increases Sensitivity to Azoles In Vitro and In Vivo in a Murine Model of Experimental Vaginitis. Antimicrob. Agents Chemother. 2018, 62, e02576-17. [Google Scholar] [CrossRef] [PubMed]
- Davis-Hanna, A.; Piispanen, A.E.; Stateva, L.I.; Hogan, D.A. Farnesol and dodecanol effects on the Candida albicans Ras1-cAMP signalling pathway and the regulation of morphogenesis. Mol. Microbiol. 2008, 67, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Neo, S.P.; Pang, L.M.; Gao, J.; Chong, S.C.; Gunaratne, J.; Wang, Y. Comprehensive Interactome Analysis for the Sole Adenylyl Cyclase Cyr1 of Candida albicans. Microbiol. Spectr. 2022, 10, e0393422. [Google Scholar] [CrossRef] [PubMed]
- Glazier, V.E. EFG1, everyone’s favorite gene in Candida albicans: A comprehensive literature review. Front. Cell Infect. Microbiol. 2022, 12, 855229. [Google Scholar] [CrossRef]
- Zeidler, U.; Lettner, T.; Lassnig, C.; Müller, M.; Lajko, R.; Hintner, H.; Breitenbach, M.; Bito, A. UME6 is a crucial downstream target of other transcriptional regulators of true hyphal development in Candida albicans. FEMS Yeast Res. 2009, 9, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, P.L.; Kadosh, D. Candida albicans Ume6, a filament-specific transcriptional regulator, directs hyphal growth via a pathway involving Hgc1 cyclin-related protein. Eukaryot. Cell 2010, 9, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V.; Feldman, M.; Smoum, R.; Mechoulam, R.; Steinberg, D. Anandamide prevents the adhesion of filamentous Candida albicans to cervical epithelial cells. Sci. Rep. 2020, 10, 13728. [Google Scholar] [CrossRef] [PubMed]
- Shinobu-Mesquita, C.S.; Martins, E.; Júnior, J.B.; de Souza Bonfim-Mendonça, P.; Felipe, M.S.S.; Kioshima, É.S.; Svidzinski, T.I.E. In vitro and in vivo activity of a possible novel antifungal small molecule against Candida albicans. J. Mycol. Med. 2020, 30, 100939. [Google Scholar] [CrossRef]
- Orsi, C.F.; Borghi, E.; Colombari, B.; Neglia, R.G.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida albicans hyphal wall protein 1 (HWP1) genotype on biofilm production and fungal susceptibility to microglial cells. Microb. Pathog. 2014, 69–70, 20–27. [Google Scholar] [CrossRef]
- König, A.; Hube, B.; Kasper, L. The Dual Function of the Fungal Toxin Candidalysin during Candida albicans-Macrophage Interaction and Virulence. Toxins 2020, 12, 469. [Google Scholar] [CrossRef]
- Bahn, Y.S. Exploiting fungal virulence-regulating transcription factors as novel antifungal drug targets. PLoS Pathog. 2015, 11, e1004936. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Mermer, A.; Bayrak, H.A.; Alyar, S.; Alagumuthu, M. Synthesis, DFT calculations, biological investigation, molecular docking studies of β-lactam derivatives. J. Mol. Struct. 2020, 1208, 127891. [Google Scholar] [CrossRef]
- Bilen, E.; Özdemir Özmen, Ü.; Çete, S.; Alyar, S.; Yaşar, A. Bioactive sulfonyl hydrazones with alkyl derivative: Characterization, ADME properties, molecular docking studies and investigation of inhibition on choline esterase enzymes for the diagnosis of Alzheimer’s disease. Chem. Biol. Interact. 2022, 1, 109956. [Google Scholar] [CrossRef] [PubMed]
- Trost, A.; Graf, B.; Eucker, J.; Sezer, O.; Possinger, K.; Göbel, U.B.; Adam, T. Identification of clinically relevant yeasts by PCR/RFLP. J. Microbiol. Methods 2004, 56, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Costa-De-Oliveira, S.; Martins, M.L.; Pina-Vaz, C.; Rodrigues, A.G.; Ludovico, P.; Rodrigues, F. Multiplex PCR identification of eight clinically relevant Candida species. Med. Mycol. 2007, 45, 619–627. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference method for broth dilution antifungal susceptibility testing of yeasts: Approved standard. In CLSI Document M27-A3, 3rd ed.; PA Clinical and Laboratory Standards Institute: Wayne, UK, 2008. [Google Scholar]
- Komaki, N.; Watanabe, T.; Ogasawara, A.; Sato, N.; Mikami, T.; Matsumoto, T. Antifungal mechanism of hinokitiol against Candida albicans. Biol. Pharm. Bull. 2008, 31, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tan, F.; Miao, H.; Wang, H.; Cao, Y. Effect of shikonin against Candida albicans biofilms. Front. Microbiol. 2019, 10, 1085. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Aydin, M.; Ozturk, A.; Duran, T.; Ozmen, U.O.; Sumlu, E.; Ayan, E.B.; Korucu, E.N. In vitro antifungal and antibiofilm activities of novel sulfonyl hydrazone derivatives against Candida spp. J. Mycol. Med. 2023, 33, 101327. [Google Scholar] [CrossRef]
- Bitencourt-Ferreira, G.; Azevedo, W.F.d. Molegro virtual docker for docking. In Docking Screens for Drug Discovery; Springer: Berlin/Heidelberg, Germany, 2019; pp. 149–167. [Google Scholar]
- Frisch, M.J.E.A. Gaussian 09, Revision d. 01; Gaussian. Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; MacKerell, A.D., Jr. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Li, X.; Liu, Q.; Lu, J.; Wang, T.; Zhang, Q. Antifungal and Antibiofilm Efficacy of Paeonol Treatment Against Biofilms Comprising Candida albicans and/or Cryptococcus neoformans. Front. Cell Infect. Microbiol. 2022, 12, 884793. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antifungal Activity MIC Range (μg/mL) | ||
|---|---|---|
| Candida Species (n) | Artemisinin | FLC |
| C. albicans (28) | 320–1280 | 0.5–4 |
| C. krusei (4) | 5 | 32–64 |
| C. tropicalis (4) | 160–320 | 1–4 |
| C. kefyr (3) | 80 | 1–4 |
| C. lusitaniae (2) | 160–640 | 1–4 |
| C. guilliermondii (2) | 20 | 2 |
| C. albicans ATCC 10231 | 640 | 0.5 |
| C. krusei ATCC 6258 | 10 | 128 |
| C. tropicalis NRRLY-12968 | 320 | 8 |
| Genes | MolDock Score | Steric Interaction | H.Bond kcal/mol | H.Bond Residue Lenght | Artemisinin–Genes * | Total Energy kcal/mol | ΔGbind kcal/mol |
|---|---|---|---|---|---|---|---|
| EFG1 | −129.466 | 119.340 | −8.838 | Arg 231 (2.862 Å) | C=O–NH | −128.187 | −9.02 |
| Val 222 (2.699 Å) | C-O-O–NH | ||||||
| Val 222 (2.628 Å) | C-O-O–NH | ||||||
| Val 230 (2.262 Å) | C-O-O–NH | ||||||
| UME6 | −126.667 | −116.532 | −7.845 | Leu 815 (2.820 Å) | C=O–NH | −127.199 | −8.79 |
| Leu 815 (2.820 Å) | C-O–NH | ||||||
| Thr 816 (3.221 Å) | C-O–NH | ||||||
| Thr 816 (3.331 Å) | C-O-O–NH | ||||||
| HGC1 | −122.299 | −114.834 | −7.118 | Leu 296 (2.334 Å) | C=O–NH | −125.250 | −8.67 |
| Lys 127 (2.331 Å) | C-O–NH3 | ||||||
| Lys 127 (3.099 Å) | C-O–NH3 | ||||||
| Lys 127 (2.334 Å) | C-O-O–NH3 | ||||||
| RAS1 | −119.998 | −111.367 | −6.123 | Leu 81 (2.462 Å) | C=O–NH | −122.980 | −8.02 |
| Val 9 (2.262 Å) | C-O–NH | ||||||
| Val 8 (2.862 Å) | C-O-O–NH | ||||||
| CYR1 | −116.250 | −109.577 | −5.844 | Arg 657 (2.822 Å) | C-O-O–NH | −120.190 | −7.86 |
| Arg 679 (2.851 Å) | C-O-O–NH2 | ||||||
| Arg 679 (2.334 Å) | C-O–NH2 | ||||||
| HWP1 | −115.699 | −108.834 | −4.891 | Cys 347 (2.875 Å) | C-O–NH | −118.384 | −7.67 |
| Cys 347 (3.099 Å) | C-O-O–NH | ||||||
| Tyr 346 (2.226 Å) | C-O–NH | ||||||
| ECE1 | −114.577 | −88.834 | −3.867 | Val 77 (2.569 Å) | C-O–NH | −113.823 | −7.02 |
| Ile 178 (2.628 Å) | C-O–NH | ||||||
| ALS3 | −113.367 | −79.834 | −3.144 | Lys 24 (2.367 Å) | C-O–NH3 | −110.144 | −6.86 |
| Glu 157 (2.331 Å) | C=O–NH |
| Genes | Forward Primer Sequence (5′→3′) | Reverse Primer Sequence (5′→3′) |
|---|---|---|
| HWP1 | GCTCCTGCTCCTGAAATGAC | CTGGAGCAATTGGTGAGGTT |
| CYR1 | CCAACAAACGACCAAAAGGT | TCTTGAACTGCCAGACGATG |
| HGC1 | GCTTCCTGCACCTCATCAAT | AGCACGAGAACCAGCGATAC |
| EFG1 | GCCTCGAGCACTTCCACTGT | TTTTTTCATCTTCCCACATGGTAGT |
| UME6 | ACCACCACTACCACCACCAC | TATCCCCATTTCCAAGTCCA |
| ECE1 | TTGCTAATGCCGTCGTCAGA | GAACGACCATCTCTCTTGGCAT |
| RAS1 | TGGATGTTGTGTTATTGTTTGAGC | GTCTTGAATTGTTCATCTTCTCCCA |
| ALS3 | TCGTCCTCATTACACCAACCA | TGAAGTTGCAGATGGGGCTT |
| 18S rRNA | AGAAACGGCTACCACATCCA | AGCCCAAGGTTCAACTACGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sumlu, E.; Aydin, M.; Korucu, E.N.; Alyar, S.; Nsangou, A.M. Artemisinin May Disrupt Hyphae Formation by Suppressing Biofilm-Related Genes of Candida albicans: In Vitro and In Silico Approaches. Antibiotics 2024, 13, 310. https://doi.org/10.3390/antibiotics13040310
Sumlu E, Aydin M, Korucu EN, Alyar S, Nsangou AM. Artemisinin May Disrupt Hyphae Formation by Suppressing Biofilm-Related Genes of Candida albicans: In Vitro and In Silico Approaches. Antibiotics. 2024; 13(4):310. https://doi.org/10.3390/antibiotics13040310
Chicago/Turabian StyleSumlu, Esra, Merve Aydin, Emine Nedime Korucu, Saliha Alyar, and Ahmed Moustapha Nsangou. 2024. "Artemisinin May Disrupt Hyphae Formation by Suppressing Biofilm-Related Genes of Candida albicans: In Vitro and In Silico Approaches" Antibiotics 13, no. 4: 310. https://doi.org/10.3390/antibiotics13040310