

Detection of Vancomycin Resistance among Methicillin-Resistant Staphylococcus aureus Strains Recovered from Children with Invasive Diseases in a Reference Pediatric Hospital
 ,
,
Abstract
:1. Introduction
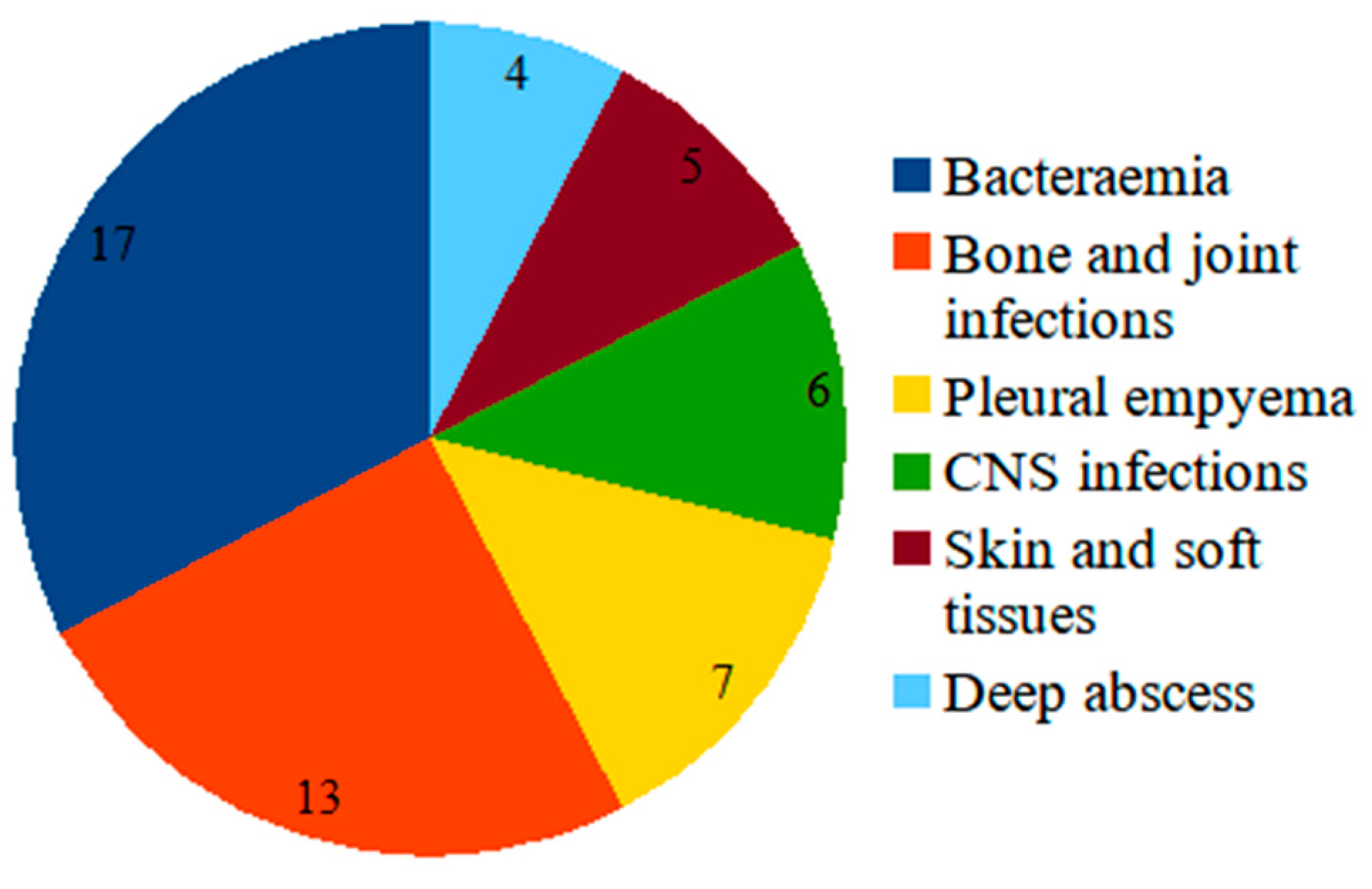
2. Results
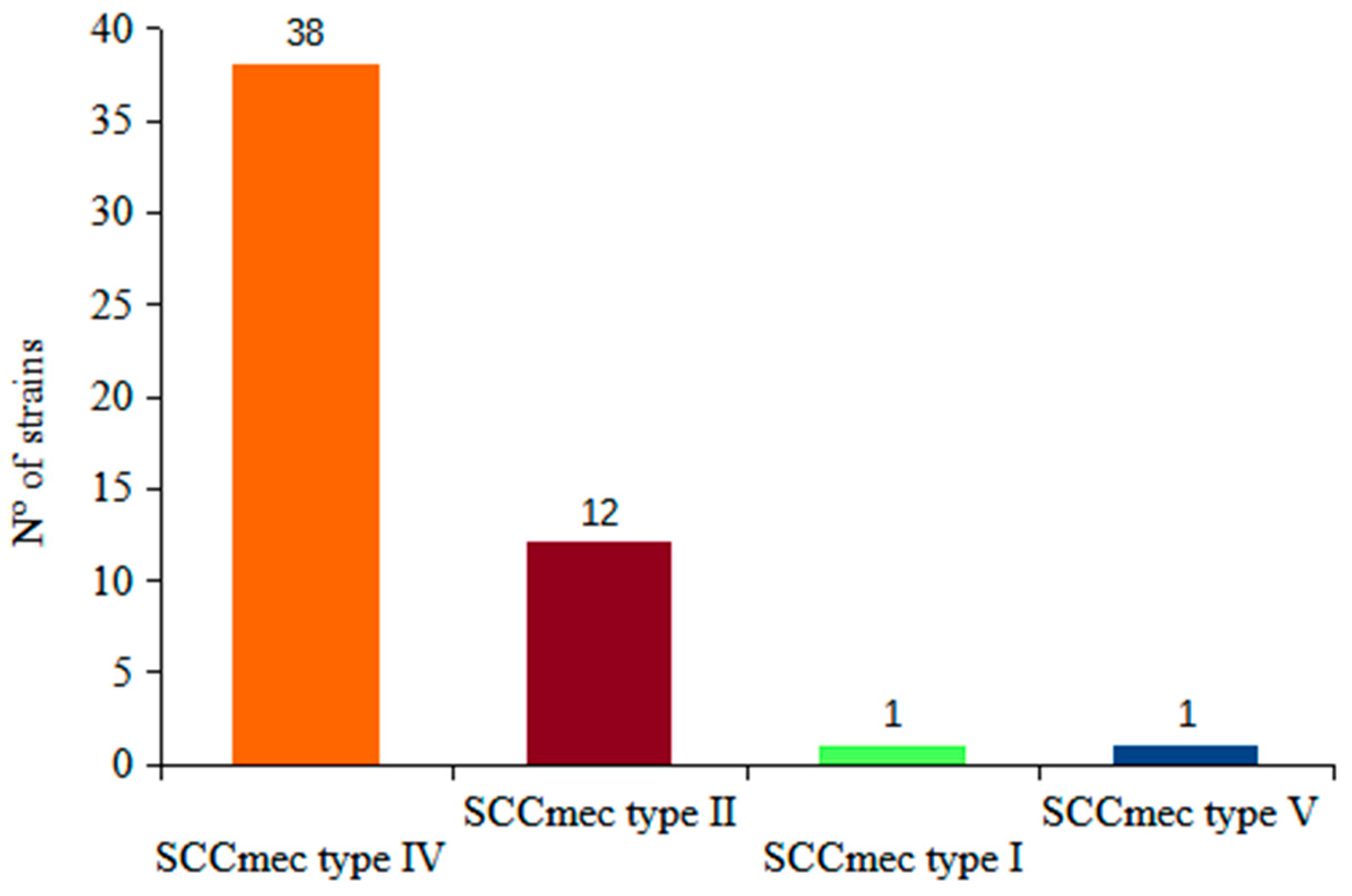
2.1. mecA, lukS-F Genes, SCCmec and MLST Genotyping
2.2. Antimicrobial Resistance Profile According to VITEK2 MICs Results
2.3. Screening for Glycopeptide Resistance and Population Analysis Profile–Area under the Curve (PAP-AUC) Assay
2.4. Microbiological Characteristics of the Suspect hVISA Strains
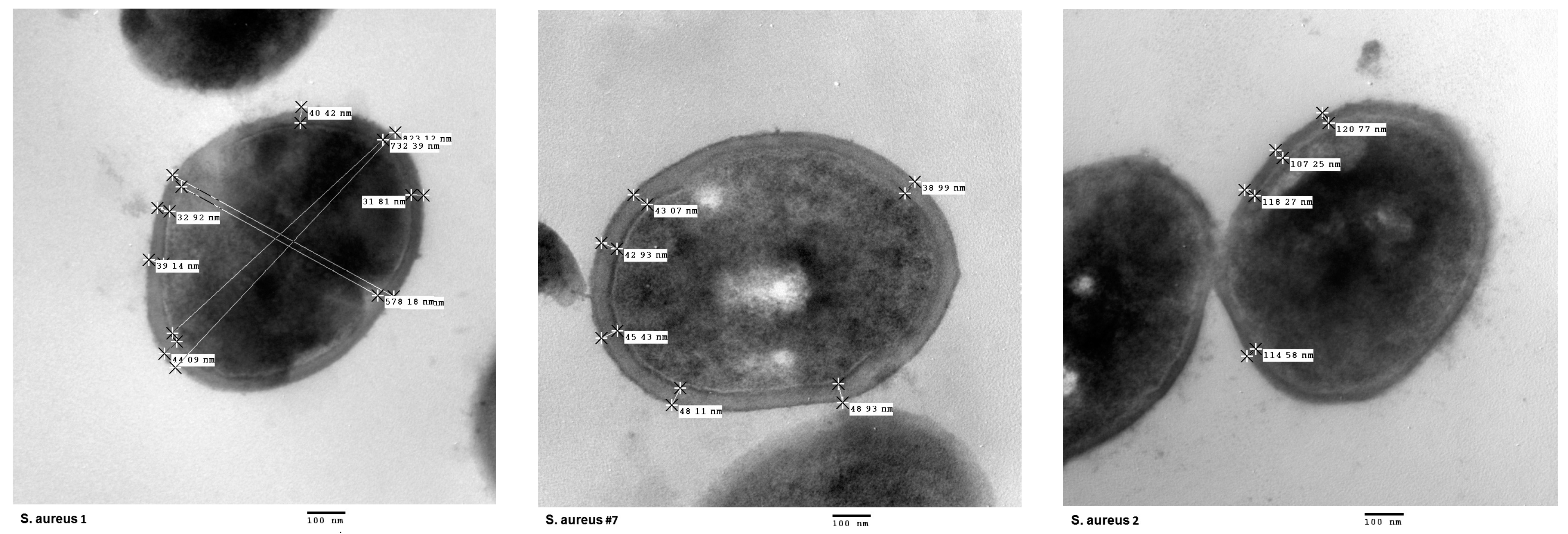
2.5. Analysis of Cell Wall Thickening
3. Discussion
4. Limitations
5. Materials and Methods
5.1. Bacterial Strains
5.2. Search for mecA, lukS-F Genes, SCCmec and Multilocus Sequence Typing (MLST)
5.3. Antimicrobial Susceptibility Testing
5.4. Screening for Resistance and Decreased Susceptibility to Glycopeptides
5.4.1. Standard E-Test
5.4.2. Macro E-Test
5.4.3. Brain–Heart Infusion–Casein–Vancomycin Plates
5.5. Study of PAP-AUC
5.6. Analysis of Cell Wall Thickening
5.7. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGuinness, W.A.; Malachowa, N.; De Leo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Hiramatsu, K. Vancomycin-Resistant Staphylococcus aureus: A new Model of Antibiotic Resistance. Lancet Infect. Dis. 2001, 1, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Moellering, R.C., Jr. The Rationale for Revising the Clinical and Laboratory Standards Institute Vancomycin Minimal Inhibitory Concentration Interpretive Criteria for Staphylococcus aureus. Clin. Infect. Dis. 2007, 44, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Kim, E.S.; Park, S.Y.; Sung, H.; Kim, M.-N.; Kim, S.-H.; Lee, S.-O.; Choi, S.-H.; Jeong, J.-Y.; Woo, J.H.; et al. Phenotypic Changes of Methicillin-Resistant Staphylococcus aureus during Vancomycin Therapy for Persistent Bacteraemia and Related Clinical Outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Peng, H.; Rao, X. Molecular Events for Promotion of Vancomycin Resistance in Vancomycin Intermediate Staphylococcus aureus. Front. Microbiol. 2016, 7, 1601. [Google Scholar] [CrossRef] [PubMed]
- Castro, B.E.; Rios, R.; Carvajal, L.P.; Vargas, M.L.; Cala, M.P.; León, L.; Hanson, B.; Dinh, A.Q.; Ortega-Recalde, O.; Seas, C.; et al. Multiomics Characterization of Methicillin-Resistant Staphylococcus aureus (MRSA) Isolates with Heterogeneous Intermediate Resistance to Vancomycin (hVISA) in Latin America. J. Antimicrob. Chemother. 2022, 78, 122–132. [Google Scholar] [CrossRef]
- Shariati, A.; Dadashi, M.; Moghadam, M.T.; van Belkum, A.; Yaslianifard, S.; Darban-Sarokhalil, D. Global Prevalence and Distribution of Vancomycin Resistant, Vancomycin Intermediate and Heterogeneously Vancomycin Intermediate Staphylococcus aureus Clinical Isolates: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 12689. [Google Scholar] [CrossRef]
- Telechea, H.; Speranza, N.; Lucas, L.; Santurio, A.; Giachetto, G.; Algorta, G.; Nanni, L.; Pírez, M.C. Antibiotic consumption and antimicrobial susceptibility evolution in the Centro Hospitalario Pereira Rossell in methicillin resistant Staphylococcus aureus era. Rev. Chil. Infectol. 2009, 26, 413–419. [Google Scholar] [CrossRef]
- Di Gregorio, S.; Perazzi, B.; Ordoñez, A.M.; DeGregorio, S.; Foccoli, M.; Lasala, M.B.; García, S.; Vay, C.; Famiglietti, A.; Mollerach, M. Clinical, Microbiological, and Genetic Characteristics of Heteroresistant Vancomycin-Intermediate Staphylococcus aureus Bacteremiaina Teaching Hospital. Microb. Drug Resist. 2015, 21, 25–34. [Google Scholar] [CrossRef]
- Chamon, R.C.; Marques, L.M.; Timenetsky, J.; da Costa Rachid, C.T.C.; Ferreira, R.B.R.; de Oliveira, T.L.R.; Glatthardt, T.; de Oliveira Moreira, L.; dos Santos, K.R.N. Genome Sequence of a Highly Virulent Pvl-Positive Vancomycin-Intermediate-Resistant Staphylococcus aureus Sequence Type 30. Curr. Genom. 2020, 21, 128–137. [Google Scholar] [CrossRef]
- Sola, C.; Lamberghini, R.O.; Ciarlantini, M.; Egea, A.L.; Gonzalez, P.; Diaz, E.G.; Huerta, V.; Gonzalez, J.; Corso, A.; Vilaro, M.; et al. Heterogeneous Vancomycin-Intermediate Susceptibility in a Community-Associated Methicillin-Resistant Staphylococcus aureus Epidemic Clone, in a Case of Infective Endocarditis in Argentina. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 15. [Google Scholar] [CrossRef]
- Di Gregorio, S.; Vielma, J.; Haim, M.S.; Rago, L.; Campos, J.; Kekre, M.; Abrudan, M.; Famiglietti, Á.; Canigia, L.F.; Rubinstein, G.; et al. Genomic Epidemiology of Staphylococcus aureus Isolated from Bloodstream Infections in South America during 2019 Supports Regional Surveillance. Microb. Genom. 2023, 9, 001020. [Google Scholar] [CrossRef]
- Pardo, L.; Vola, M.; Macedo-Viñas, M.; Machado, V.; Cuello, D.; Mollerach, M.; Castro, M.; Pírez, C.; Varela, G.; Algorta, G. Community-Associated Methicillin-Resistant Staphylococcus aureus in Children Treated in Uruguay. J. Infect. Dev. Ctries. 2013, 7, 10–16. [Google Scholar] [CrossRef]
- Ma, X.X.; Galiana, A.; Pedreira, W.; Mowszowicz, M.; Christophersen, I.; Machiavello, S.; Lope, L.; Benaderet, S.; Buela, F.; Vicentino, W.; et al. Community-Acquired Methicillin-Resistant Staphylococcus aureus, Uruguay. Emerg. Infect. Dis. 2005, 11, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Park, J.; Chung, H.-S. Genetic Characterization of Tetracycline-Resistant Staphylococcus aureus with Reduced Vancomycin Susceptibility Using Whole-Genome Sequencing. Arch. Microbiol. 2023, 206, 24. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Peleg, A.Y.; Stinear, T.P. The Evolution of Vancomycin Intermediate Staphylococcus aureus (VISA) and Heterogenous-VISA. Infect. Genet. Evol. 2014, 21, 575–582. [Google Scholar] [CrossRef]
- Richter, S.S.; Satola, S.W.; Crispell, E.K.; Heilmann, K.P.; Dohrn, C.L.; Riahi, F.; Costello, A.J.; Diekema, D.J.; Doern, G.V. Detection of Staphylococcus aureus Isolates with Heterogeneous Intermediate-Level Resistance to Vancomycin in the United States. J. Clin. Microbiol. 2011, 49, 4203–4207. [Google Scholar] [CrossRef]
- Khatib, R.; Riederer, K.; Sharma, M.; Shemes, S.; Iyer, S.P.; Szpunar, S. Screening for Intermediately Vancomycin-Susceptible and Vancomycin-Heteroresistant Staphylococcus aureus by Use of Vancomycin-Supplemented Brain Heart Infusion Agar Biplates: Defining Growth Interpretation Criteria Based on Gold Standard Confirmation. J. Clin. Microbiol. 2015, 53, 3543–3546. [Google Scholar] [CrossRef] [PubMed]
- Satola, S.W.; Farley, M.M.; Anderson, K.F.; Patel, J.B. Comparison of Detection Methods for Heteroresistant Vancomycin-Intermediate Staphylococcus aureus, with the Population Analysis Profile Method as the Reference Method. J. Clin. Microbiol. 2020, 49, 177–183. [Google Scholar] [CrossRef]
- Katayama, Y.; Azechi, T.; Miyazaki, M.; Takata, T.; Sekine, M.; Matsui, H.; Hanaki, H.; Yahara, K.; Sasano, H.; Asakura, K.; et al. Prevalence of Slow-Growth Vancomycin Non susceptibility in Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2017, 61, e00452-17. [Google Scholar] [CrossRef]
- Wootton, M.; Howe, R.A.; Hillman, R.; Walsh, T.R.; Bennett, P.M.; MacGowan, A.P. A Modified Population Analysis Profile (PAP) Method to Detect Hetero-Resistance to Vancomycin in Staphylococcus aureus in a UK Hospital. J. Antimicrob. Chemother. 2001, 47, 399–403. [Google Scholar] [CrossRef]
- Investigation and Control of Vancomycin-Resistant Staphylococcus aureus (VRSA): 2015 Update. Available online: https://www.cdc.gov/hai/pdfs/vrsa-investigation-guide-05_12_2015.pdf (accessed on 4 January 2024).
- Silveira, A.C.d.O.; Sambrano, G.E.; Paim, T.G.d.S.; Caierão, J.; de Cordova, C.M.M.; d’Azevedo, P.A. Is Pre diffusion Test an Alternative to Improve Accuracy in Screening hVISA Strains and to Detect Susceptibility to Glycopeptides/Lipopeptides? Diagn. Microbiol. Infect. Dis. 2014, 79, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR Strategy for Rapid Identification of Structural Types and Variants of the Mec Element in Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of Multiplex PCRs for Staphylococcal Cassette Chromosome mec Type Assignment: Rapid Identification System for Mec, Ccr, and Major Differences in Junkyard Regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [PubMed]
- SCCmec International Working Group on the Staphylococcal Cassette Cromosome Elements (IWG-SCC). Available online: https://www.sccmec.org/index.php/en/ (accessed on 24 February 2024).
- Lina, G.; Piémont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.-O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine Leukocidin—Producing Staphylococcus aureus in Primary Skin Infections and Pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterization of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed.; CLSIsupplementM100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023; pp. 93–105. [Google Scholar]
- Cázares-Domínguez, V.; Cruz-Córdova, A.; Ochoa, S.A.; Escalona, G.; Arellano-Galindo, J.; Rodríguez-Leviz, A.; Hernández-Castro, R.; López-Villegas, E.O.; Xicohtencatl-Cortes, J. Vancomycin Tolerant, Methicillin-Resistant Staphylococcus aureus Reveals the Effects of Vancomycin on Cell Wall Thickening. PLoS ONE 2015, 10, e0118791. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain Number | AUC Strain | AUC Strain/AUCμ3 | VITEK® VA TE | E-Test VA TE | Macro E-Test VA TE | BHI424 VA (CFU) | BHI448 VA (CFU) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 16.05 | 0.799 | 1 | 0.5 | 1 | 0.5 | 3 | 3 | 1 | 1 |
| 2 | 16.3 | 0.811 | 0.5 | 0.5 | 1 | 0.75 | 3 | 3 | 1 | 4 |
| 3 | 17.55 | 0.873 | 0.5 | 0.5 | 1 | 1.5 | 1 | 2 | 0 | 0 |
| 4 | 15.25 | 0.759 | 0.5 | 0.5 | 1 | 0.25 | 1.5 | 1.5 | 0 | 1 |
| 5 | 13.98 | 0.685 | 1 | 0.5 | 1 | 0.75 | 1.5 | 1.5 | 0 | 1 |
| 6 | 16.72 | 0.832 | 1 | 0.5 | 1 | 0.75 | 2 | 2 | 0 | 1 |
| 7 | 19.01 | 0.946 | 1 | 0.5 | 1 | 0.75 | 2 | 1.5 | 0 | 1 |
| 8 | 14.48 | 0.720 | 1 | 0.5 | 1 | 1 | 2 | 3 | 1 | 1 |
| 9 | 17.15 | 0.853 | 1 | 0.5 | 1.5 | 0.38 | 2 | 3 | 0 | 0 |
| 10 | 9.83 | 0.489 | 0.5 | 0.5 | 1 | 0.75 | 3 | 2 | 1 | 2 |
| 11 | 16.48 | 0.822 | 1 | 0.5 | 1.5 | 1 | 2 | 2 | 0 | 0 |
| 12 | 16.01 | 0.797 | 1 | 0.5 | 1.5 | 0.5 | 2 | 2 | 0 | 0 |
| 13 | 9.46 | 0.471 | 0.5 | 0.5 | 1 | 0.5 | 3 | 3 | 1 | 1 |
| 14 | 16.4 | 0.816 | 1 | 0.5 | 0.75 | 1 | 2 | 3 | 2 | 2 |
| 15 | 17.55 | 0.873 | 1 | 0.5 | 1 | 0.75 | 1.5 | 2 | 1 | 1 |
| μ3 | 20.09 | 1 | ND | ND | ND | ND | ND | ND | ND | ND |
| Strain Number | AUC Strain/AUCμ3 | Source | lukS-F Gene | SCCmec Type | Resistance Phenotype | Sequence Type /Clonal Complex |
|---|---|---|---|---|---|---|
| 1 | 0.799 | bacteremia | − | II | OX, CIP, cMLSb | ST-868/CC-5 |
| 2 | 0.811 | adenophlegmon | + | IV | OX | ST-8/CC-8 |
| 3 | 0.873 | arthritis | − | IV | OX | ND |
| 4 | 0.759 | bacteremia | − | II | OX, CIP, cMLSb | ST-5/CC-5 |
| 5 | 0.685 | bacteremia | − | IV | OX, iMLSb | ST-30/CC-30 |
| 6 | 0.832 | bacteremia | + | IV | OX | ND |
| 7 | 0.946 | bacteremia | + | IV | OX | ST-30/CC-30 |
| 8 | 0.720 | pneumonia | − | II | OX, CIP, cMLSb, STX, GN | ST-5/CC-5 |
| 9 | 0.853 | arthritis | − | V | OX | ST-8/CC-8 |
| 10 | 0.489 | osteomyelitis | + | IV | OX, iMLSb | ND |
| 11 | 0.822 | endocarditis | − | IV | OX | ST-30/CC-30 |
| 12 | 0.797 | osteomyelitis | + | II | OX | ST-577/CC-121 |
| 13 | 0.471 | arthritis | + | IV | OX | ND |
| 14 | 0.816 | arthritis | − | V | OX | ND |
| 15 | 0.873 | pneumonia | − | IV | OX, iMLSb | ST-434/CC-30 |
| Primer Name | SCCmec Amplicon | Oligonucleotide Sequence (5′-3′) | Amplicon Size |
|---|---|---|---|
| mI-1 | mec complex A | AATGGCGAAAAAGCACAACA | 480 bp |
| mI-2 | mec complex A | GACTTGATTGTTTCCTCTGTT | |
| mcR-2 | mec complex A | CGCTCAGAAATTTGTTGTGC | 1597 bp |
| mcR-3 | mec complex A | ATCTCCACGTTAATTCCATT | |
| IS1272 | mec complex B | ATTTTGGGTTTCACTCGGAT | 565 bp |
| mecR1 | mec complex B | CAAATATTAAAGAACGTGTT | |
| mA7 | mec complex C | ATATACCAAACCCGACAACTACA | 804 bp |
| iS2 | mec complex C | TGAGGTTATTCAGATATTTCGATGT | |
| ccr-β2 | ccrAB1 | ATTGCCTTGATAATAGCCITCT | 700 bp |
| ccr-α2 | ccrAB1 | AACCTATATCATCAATCAGTACGT | |
| ccr-β2 | ccrAB2 | ATTGCCTTGATAATAGCCITCT | 1000 bp |
| ccr-α3 | ccrAB2 | TAAAGGCATCAATGCACAAACACT | |
| ccr-β2 | ccrAB3 | ATTGCCTTGATAATAGCCITCT | 1600 bp |
| ccr-α4 | ccrAB3 | GCTCAAAAGCAAGCAATAGAAT | |
| ccrC-F2 | ccrC | GTACTCGTTACAATGTTTGG | 449 bp |
| ccrC-R2 | ccrC | ATAATGGCTTCATGCTTACC |
| SCCmecType | CcrComplex | MecComplex |
|---|---|---|
| I | A1B1 | B |
| II | A2B2 | A |
| III | A3B3 | A |
| IV | A2B2 | B |
| V | C1 | C2 |
| VI | A4B4 | B |
| VII | C1 | C1 |
| VIII | A4B4 | A |
| IX | A1B1 | C2 |
| X | A1B6 | C1 |
| XI | A1B3 | E |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo, L.; Mota, M.I.; Parnizari, A.; Varela, A.; Algorta, G.; Varela, G. Detection of Vancomycin Resistance among Methicillin-Resistant Staphylococcus aureus Strains Recovered from Children with Invasive Diseases in a Reference Pediatric Hospital. Antibiotics 2024, 13, 298. https://doi.org/10.3390/antibiotics13040298
Pardo L, Mota MI, Parnizari A, Varela A, Algorta G, Varela G. Detection of Vancomycin Resistance among Methicillin-Resistant Staphylococcus aureus Strains Recovered from Children with Invasive Diseases in a Reference Pediatric Hospital. Antibiotics. 2024; 13(4):298. https://doi.org/10.3390/antibiotics13040298
Chicago/Turabian StylePardo, Lorena, María Inés Mota, Andrés Parnizari, Adriana Varela, Gabriela Algorta, and Gustavo Varela. 2024. "Detection of Vancomycin Resistance among Methicillin-Resistant Staphylococcus aureus Strains Recovered from Children with Invasive Diseases in a Reference Pediatric Hospital" Antibiotics 13, no. 4: 298. https://doi.org/10.3390/antibiotics13040298