
Virulence of Pseudomonas aeruginosa in Cystic Fibrosis: Relationships between Normoxia and Anoxia Lifestyle
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
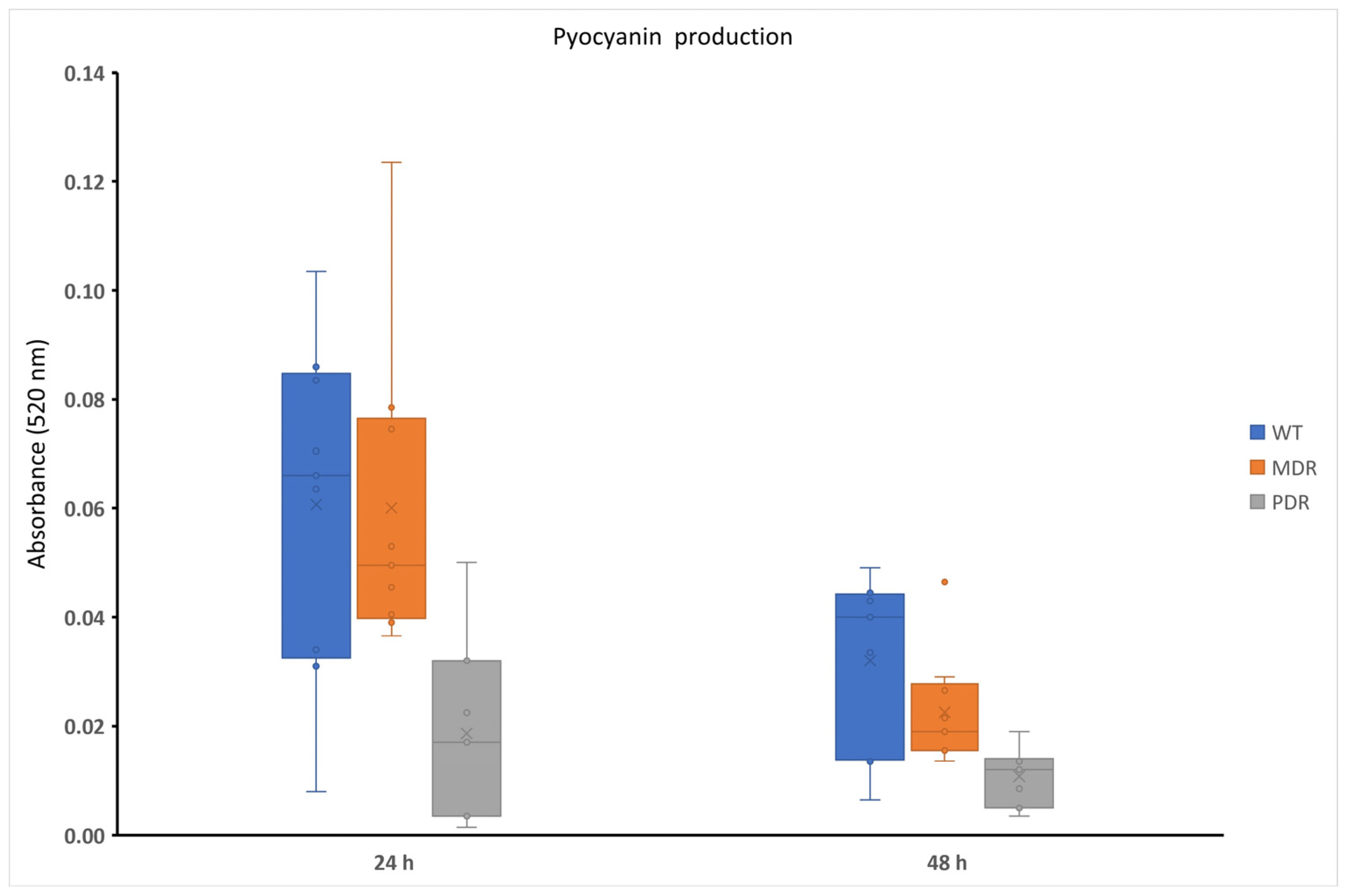
2.1. Pyocyanin Production in P. aeruginosa during CF Infection
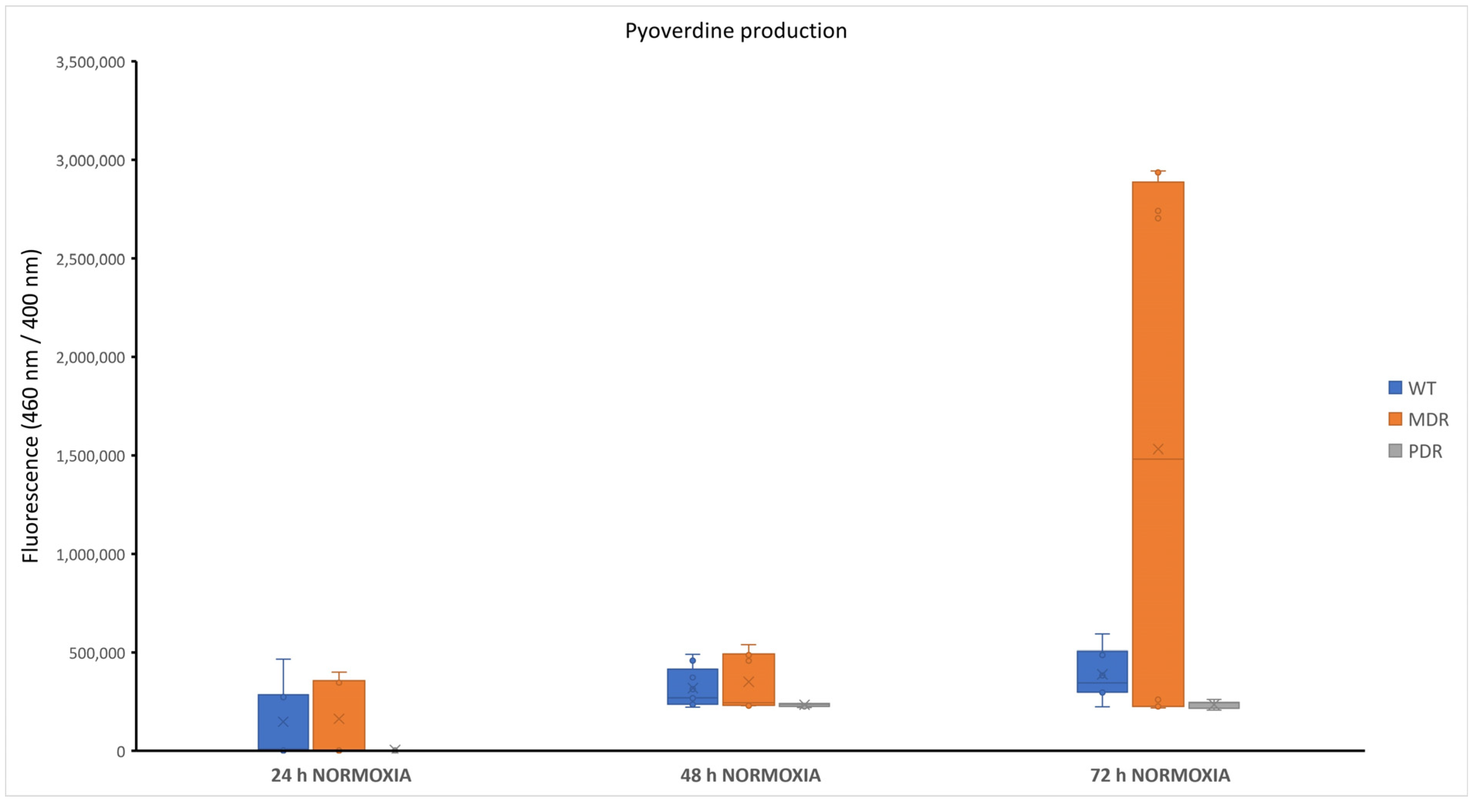
2.2. Pyoverdine Production in P. aeruginosa during CF Infection
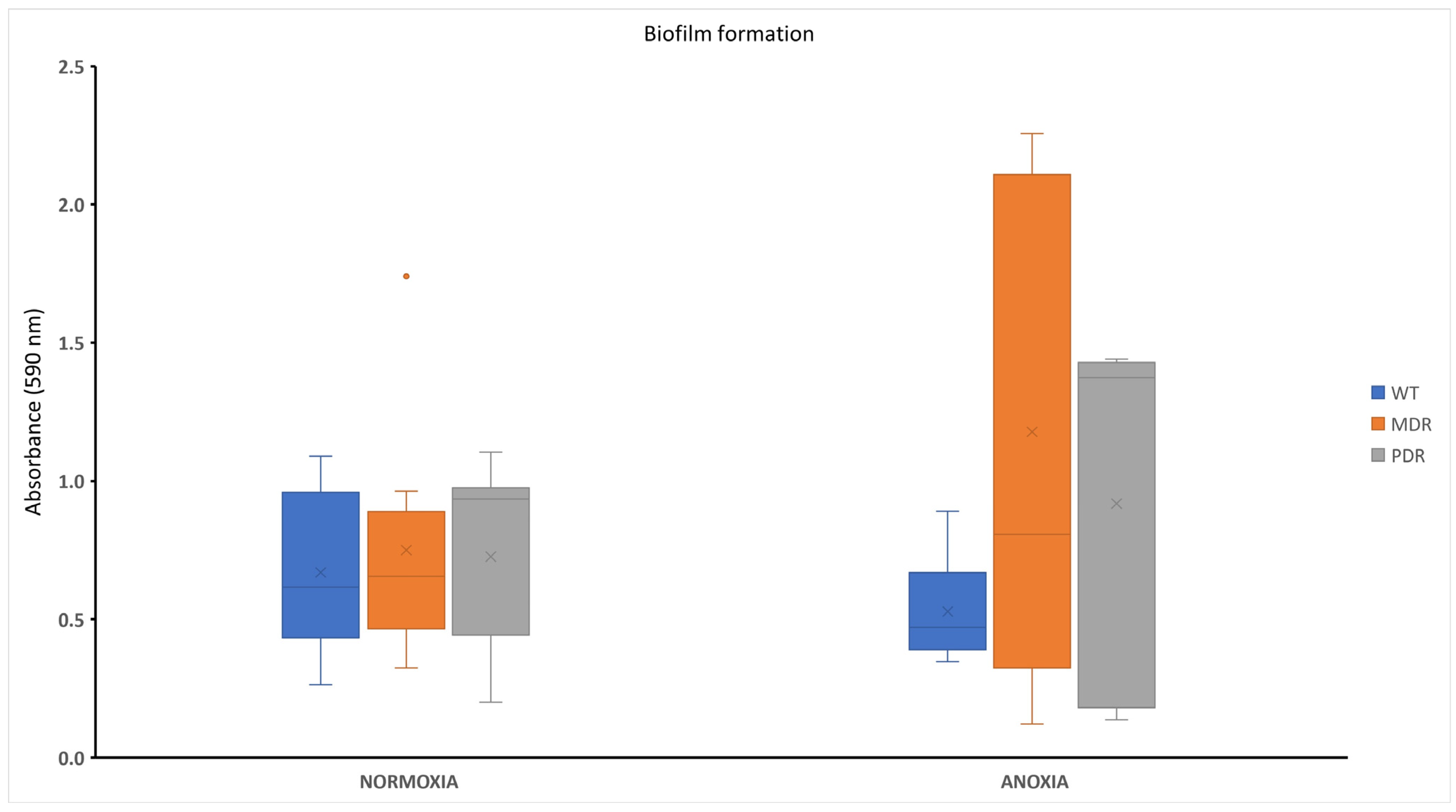
2.3. Biofilm Formation in P. aeruginosa during CF Infection
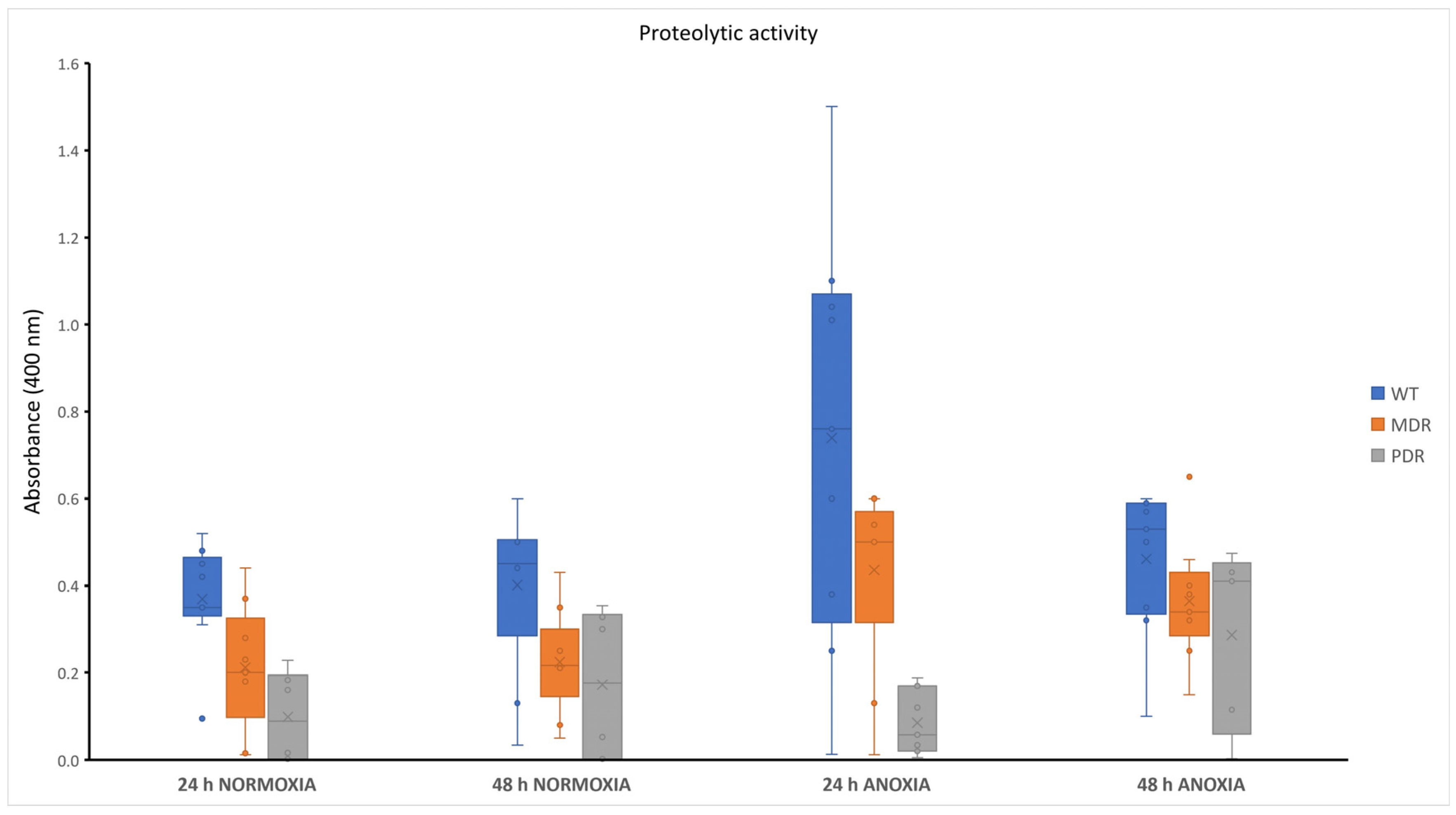
2.4. Proteolytic Content in P. aeruginosa during CF Infection
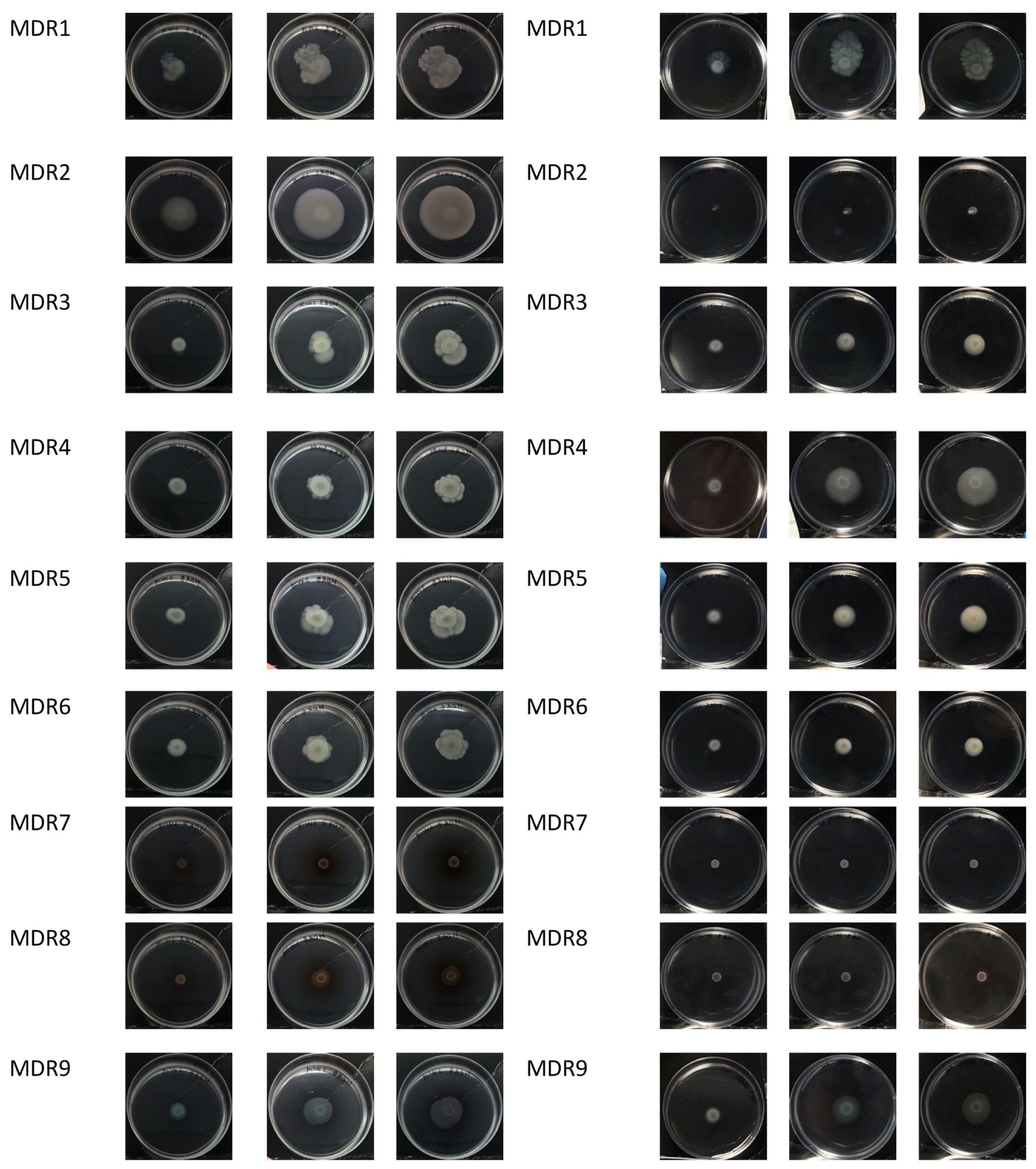
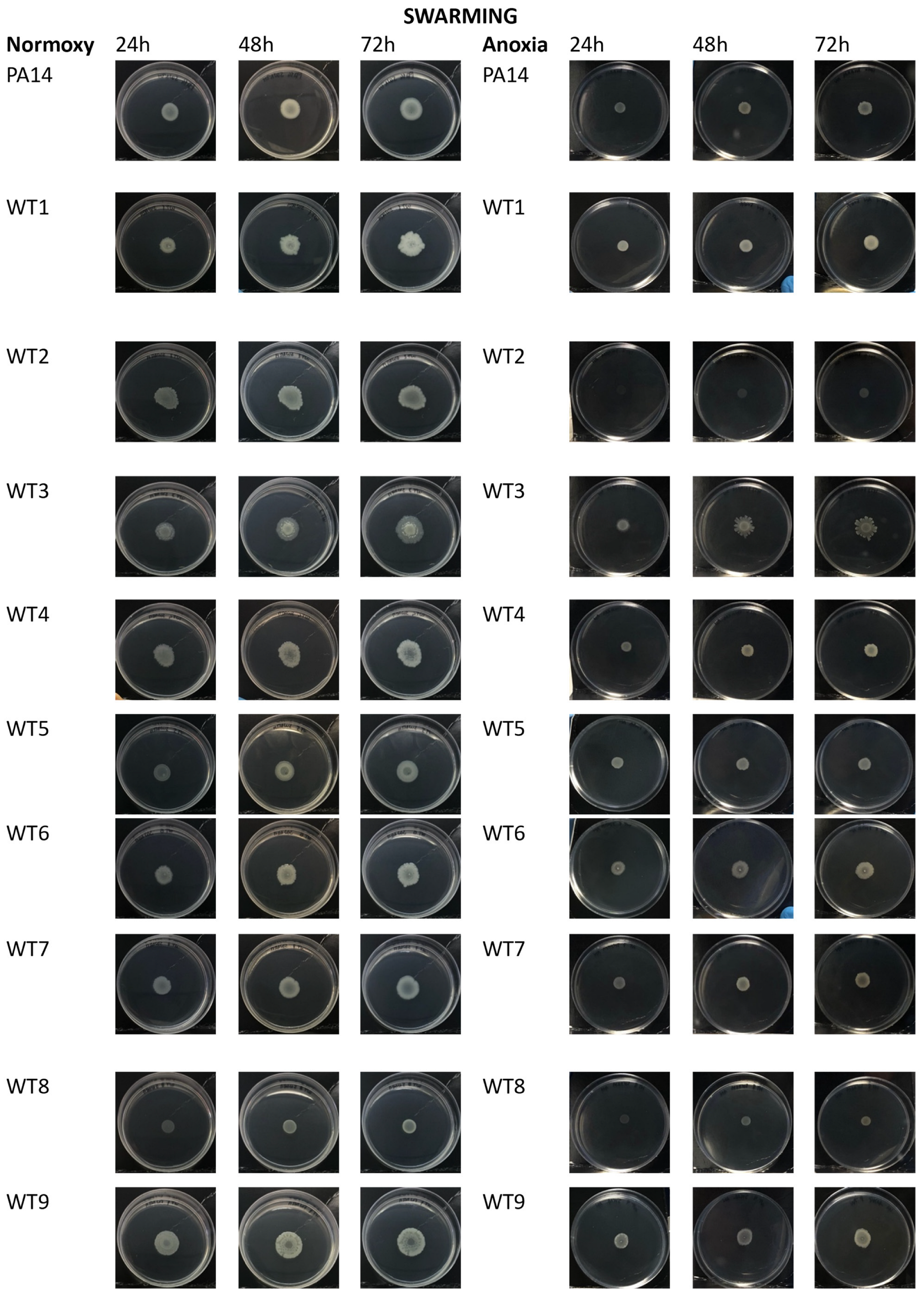
2.5. Motility in P. aeruginosa during CF Infection
3. Discussion
4. Materials and Methods
4.1. Ethics Approval and Informed Consent
4.2. Bacterial Strains and Growth Conditions
4.3. Biofilm Formation
4.4. Pyocyanin Assay
4.5. Pyoverdine Assay
4.6. Protease Assay
4.7. Zymography Assay
4.8. Motility Assays
4.8.1. Swarming Assay
4.8.2. Swimming Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef] [PubMed]
- Castellani, C.; Assael, B.M. Cystic fibrosis: A clinical view. Cell Mol. Life Sci. 2017, 74, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Cabrini, G. Innovative Therapies for Cystic Fibrosis: The Road from Treatment to Cure. Mol. Diagn. Ther. 2019, 23, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Imperlini, E.; Papa, R. Clinical Advances in Cystic Fibrosis. J. Clin. Med. 2022, 11, 6306. [Google Scholar] [CrossRef] [PubMed]
- Schick, A.; Kassen, R. Rapid diversification of Pseudomonas aeruginosa in cystic fibrosis lung-like conditions. Proc. Natl. Acad. Sci. USA 2018, 115, 10714–10719. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Martín, I.; Sainz-Mejías, M.; McClean, S. Pseudomonas aeruginosa: An Audacious Pathogen with an Adaptable Arsenal of Virulence Factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, S.A.; Ahn, D.; Prince, A. Pseudomonas aeruginosa and Klebsiella pneumoniae Adaptation to Innate Immune Clearance Mechanisms in the Lung. J. Innate Immun. 2018, 10, 442–454. [Google Scholar] [CrossRef]
- Bajire, S.K.; Jain, S.; Johnson, R.P.; Shastry, R.P. 6-Methylcoumarin attenuates quorum sensing and biofilm formation in Pseudomonas aeruginosa PAO1 and its applications on solid surface coatings with polyurethane. Appl. Microbiol. Biotechnol. 2021, 105, 8647–8661. [Google Scholar] [CrossRef]
- Yaeger, L.N.; Coles, V.E.; Chan, D.C.K.; Burrows, L.L. How to kill Pseudomonas-emerging therapies for a challenging pathogen. Ann. N. Y. Acad. Sci. 2021, 1496, 59–81. [Google Scholar] [CrossRef]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell. 2002, 3, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Hassett, D.J.; Sutton, M.D.; Schurr, M.J.; Herr, A.B.; Caldwell, C.C.; Matu, J.O. Pseudomonas aeruginosa hypoxic or anaerobic biofilm infections within cystic fibrosis airways. Trends Microbiol. 2009, 17, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; McCoy, K.S. Biofilm aggregates and the host airway-microbial interface. Front. Cell Infect. Microbiol. 2022, 12, 969326. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.W.; Gray, A.R.; Brockway, B.; Lamont, I.L. Pseudomonas aeruginosa is oxygen-deprived during infection in cystic fibrosis lungs, reducing the effectiveness of antibiotics. FEMS Microbiol. Lett. 2023, 370, fnad076. [Google Scholar] [CrossRef] [PubMed]
- Rada, B.; Leto, T.L. Pyocyanin effects on respiratory epithelium: Relevance in Pseudomonas aeruginosa airway infections. Trends Microbiol. 2013, 21, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Jayaseelan, S.; Ramaswamy, D.; Dharmaraj, S. Pyocyanin: Production, applications, challenges and new insights. World J. Microbiol. Biotechnol. 2014, 30, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; Imperi, F.; Lamont, I.L. Pyoverdine siderophores: From biogenesis to biosignificance. Trends Microbiol. 2007, 15, 22–30. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef]
- La Rosa, R.; Rossi, E.; Feist, A.M.; Johansen, H.K.; Molin, S. Compensatory evolution of Pseudomonas aeruginosa’s slow growth phenotype suggests mechanisms of adaptation in cystic fibrosis. Nat. Commun. 2021, 12, 3186. [Google Scholar] [CrossRef]
- Montemari, A.L.; Marzano, V.; Essa, N.; Levi Mortera, S.; Rossitto, M.; Gardini, S.; Selan, L.; Vrenna, G.; Onetti Muda, A.; Putignani, L.; et al. A Shaving Proteomic Approach to Unveil Surface Proteins Modulation of Multi-Drug Resistant Pseudomonas aeruginosa Strains Isolated From Cystic Fibrosis Patients. Front. Med. 2022, 9, 818669. [Google Scholar] [CrossRef]
- Li, D.Y.; Han, J.T.; Zhang, M.Y.; Yan, X.; Zhang, N.; Ge, H.; Wang, Z.; He, Y.X. The Two-Component System RstA/RstB Regulates Expression of Multiple Efflux Pumps and Influences Anaerobic Nitrate Respiration in Pseudomonas fluorescens. mSystems 2021, 6, e0091121. [Google Scholar] [CrossRef] [PubMed]
- Pallett, R.; Leslie, L.J.; Lambert, P.A.; Milic, I.; Devitt, A.; Marshall, L.J. Anaerobiosis influences virulence properties of Pseudomonas aeruginosa cystic fibrosis isolates and the interaction with Staphylococcus aureus. Sci. Rep. 2019, 9, 6748. [Google Scholar] [CrossRef] [PubMed]
- Britigan, B.E.; Roeder, T.L.; Rasmussen, G.T.; Shasby, D.M.; McCormick, M.L.; Cox, C.D. Interaction of the Pseudomonas aeruginosa secretory products pyocyanin and pyochelin generates hydroxyl radical and causes synergistic damage to endothelial cells. Implications for Pseudomonas-associated tissue injury. J. Clin. Investig. 1992, 90, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.C.; Klepac-Ceraj, V.; Lorenzi, M.M.; Grotzinger, H.; Martin, T.R.; Newman, D.K. Phenazine content in the cystic fibrosis respiratory tract negatively correlates with lung function and microbial complexity. Am. J. Respir. Cell Mol. Biol. 2012, 47, 738–745. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Perraud, Q. Pseudomonas aeruginosa and its multiple strategies to access iron. Environ. microbiol. 2023, 25, 811–831. [Google Scholar] [CrossRef] [PubMed]
- Touati, D. Iron and oxidative stress in bacteria. Arch. Biochem. Biophys. 2000, 373, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.J.; Khan, F.; Khan, S.; Tabassum, N.; Mehta, S.; Kim, Y.M. Pseudomonas aeruginosa virulence attenuation by inhibiting siderophore functions. Appl. Microbiol. Biotechnol. 2023, 107, 1019–1038. [Google Scholar] [CrossRef] [PubMed]
- Konovalova, A.; Søgaard-Andersen, L.; Kroos, L. Regulated proteolysis in bacterial development. FEMS Microbiol. Rev. 2014, 38, 493–522. [Google Scholar] [CrossRef]
- Dong, S.; Chen, H.; Zhou, Q.; Liao, N. Protein degradation control and regulation of bacterial survival and pathogenicity: The role of protein degradation systems in bacteria. Mol. Biol. Rep. 2021, 48, 7575–7585. [Google Scholar] [CrossRef]
- Artini, M.; Imperlini, E.; Buonocore, F.; Relucenti, M.; Porcelli, F.; Donfrancesco, O.; Tuccio Guarna Assanti, V.; Fiscarelli, E.V.; Papa, R.; Selan, L. Anti-Virulence Potential of a Chionodracine-Derived Peptide against Multidrug-Resistant Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients. Int. J. Mol. Sci. 2022, 23, 13494. [Google Scholar] [CrossRef]
- Artini, M.; Vrenna, G.; Trecca, M.; Tuccio Guarna Assanti, V.; Fiscarelli, E.V.; Papa, R.; Selan, L. Serratiopeptidase Affects the Physiology of Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Int. J. Mol. Sci. 2022, 23, 12645. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.J.; Vasil, A.I.; Johnson, Z.; Wilson, M.J.; Cunliffe, H.E.; Lamont, I.L.; Vasil, M.L. Characterization of an endoprotease (PrpL) encoded by a PvdS-regulated gene in Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Kessler, E.; Safrin, M. Elastinolytic and proteolytic enzymes. Methods Mol. Biol. 2014, 1149, 135–169. [Google Scholar] [PubMed]
- Yang, R.; Guan, Y.; Zhou, J.; Sun, B.; Wang, Z.; Chen, H.; He, Z.; Jia, A. Phytochemicals from Camellia nitidissima chi flowers reduce the pyocyanin production and motility of Pseudomonas aeruginosa PAO1. Front. Microbiol. 2018, 8, 2640. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Fluoroquinilones | Penicillins | Monobactams | Cephalosporins | Aminoglycosides | Carbapenems | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CIP | LEV | TZP | ATM | CAZ | FEP | CZA | C/T | AK | TOB | IM | MRP | |
| 5 µg | 5 µg | 30–6 µg | 30 µg | 10 µg | 30 µg | 10–4 µg | 30–10 µg | 30 µg | 10 µg | 10 µg | 10 µg | |
| PA14 | S | S | S | S | S | S | - | - | S | S | S | S |
| WT1 | I | R | I | I | I | I | - | - | S | S | I | S |
| WT2 | I | I | I | I | I | I | - | - | S | S | I | S |
| WT3 | I | I | I | - | I | I | S | S | S | S | I | S |
| WT4 | I | I | I | - | I | I | S | S | S | S | I | S |
| WT5 | I | I | S | - | I | I | - | - | S | I | - | S |
| WT6 | I | I | I | - | I | I | S | S | S | S | I | S |
| WT7 | I | I | I | I | I | I | - | - | S | S | I | S |
| WT8 | I | I | I | - | I | I | S | S | S | S | I | S |
| WT9 | I | I | I | - | I | I | S | S | S | S | I | S |
| MDR1 | R | R | R | R | R | R | R | R | R | R | R | R |
| MDR2 | I | R | R | R | R | R | R | R | R | R | R | R |
| MDR3 | I | R | R | R | R | R | S | R | R | R | R | I |
| MDR4 | I | R | R | R | R | R | S | S | R | R | R | I |
| MDR5 | R | R | R | R | R | R | S | S | R | R | R | R |
| MDR6 | R | R | S | S | S | S | - | - | R | R | R | R |
| MDR7 | R | R | R | R | R | R | S | S | R | R | R | R |
| MD8 | R | R | R | R | R | R | S | R | S | S | R | I |
| MDR9 | R | R | I | R | S | R | S | S | R | R | I | R |
| PDR2 | R | R | R | I | R | R | R | R | R | R | R | R |
| PDR3 | R | R | R | R | R | R | - | - | R | R | R | R |
| PDR4 | R | R | R | R | R | R | R | R | R | R | R | R |
| PDR5 | R | R | R | R | R | R | R | R | R | R | R | R |
| PDR6 | R | R | R | R | R | R | R | R | R | R | R | R |
| PDR7 | R | R | R | R | R | R | R | R | R | R | R | R |
| PDR8A | R | R | R | R | R | R | R | R | R | R | R | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papa, R.; Imperlini, E.; Trecca, M.; Paris, I.; Vrenna, G.; Artini, M.; Selan, L. Virulence of Pseudomonas aeruginosa in Cystic Fibrosis: Relationships between Normoxia and Anoxia Lifestyle. Antibiotics 2024, 13, 1. https://doi.org/10.3390/antibiotics13010001
Papa R, Imperlini E, Trecca M, Paris I, Vrenna G, Artini M, Selan L. Virulence of Pseudomonas aeruginosa in Cystic Fibrosis: Relationships between Normoxia and Anoxia Lifestyle. Antibiotics. 2024; 13(1):1. https://doi.org/10.3390/antibiotics13010001
Chicago/Turabian StylePapa, Rosanna, Esther Imperlini, Marika Trecca, Irene Paris, Gianluca Vrenna, Marco Artini, and Laura Selan. 2024. "Virulence of Pseudomonas aeruginosa in Cystic Fibrosis: Relationships between Normoxia and Anoxia Lifestyle" Antibiotics 13, no. 1: 1. https://doi.org/10.3390/antibiotics13010001