
Distribution of Antibiotic Resistance in a Mixed-Use Watershed and the Impact of Wastewater Treatment Plants on Antibiotic Resistance in Surface Water
 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
2.1. Prevalence and Distribution of AR Contaminants in Surface Water
2.1.1. Antibiotic-Resistant Salmonella, Escherichia coli, and Enterococcus
2.1.2. ESBL-Producing Enterobacteriaceae and Carbapenem-Resistant Enterobacteriaceae (CRE)
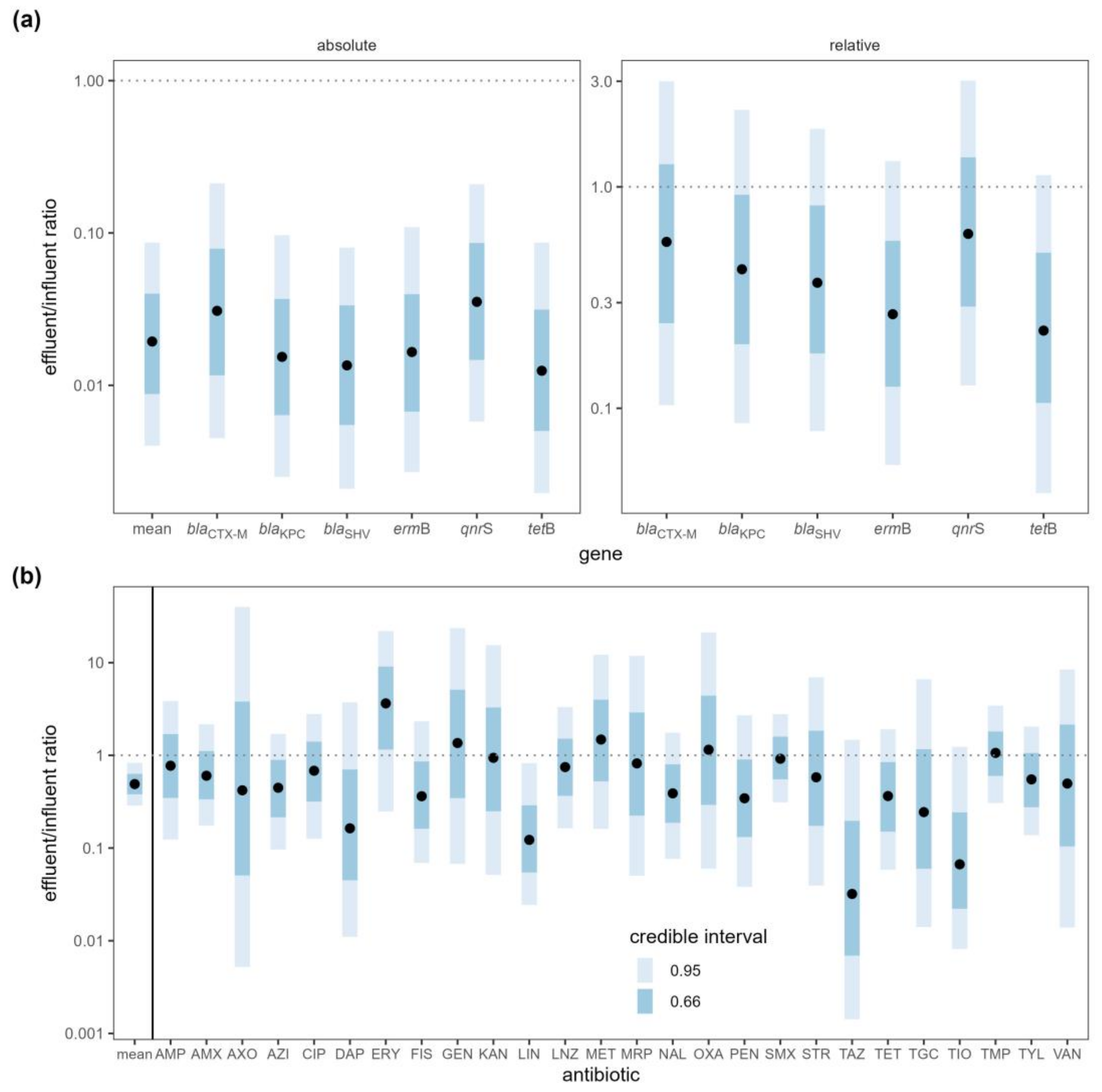
2.1.3. Antibiotic Resistance Gene Markers
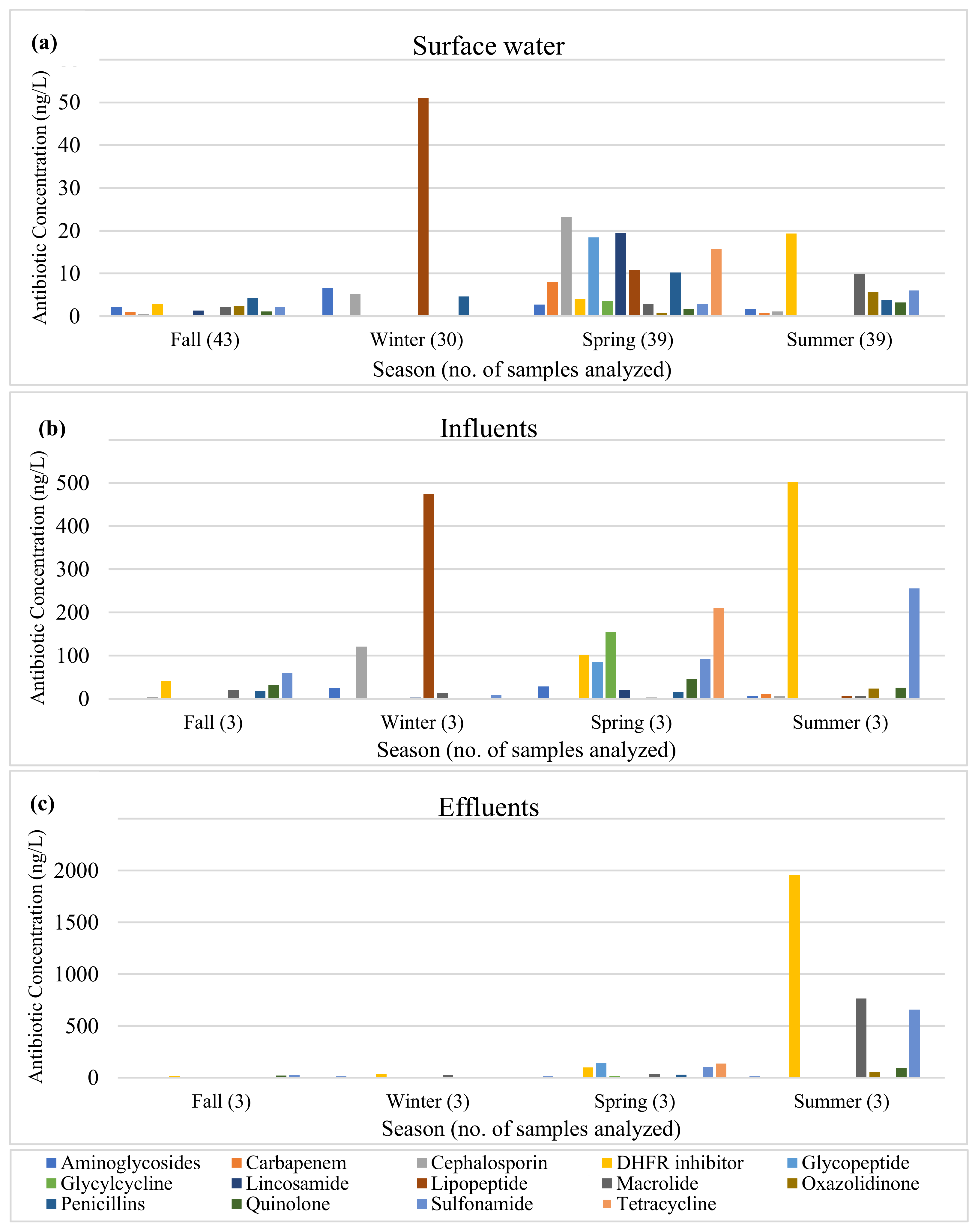
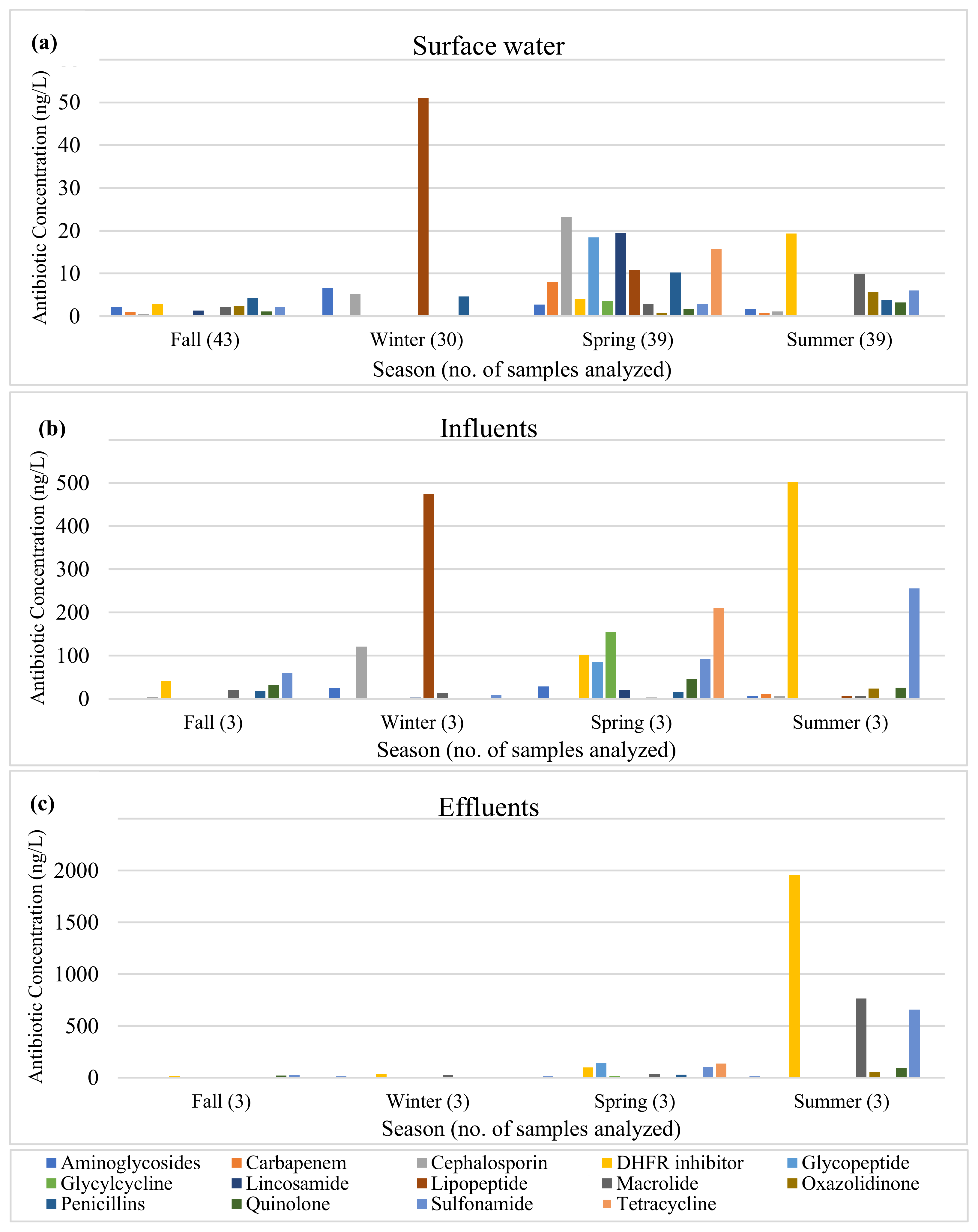
2.1.4. Antibiotics
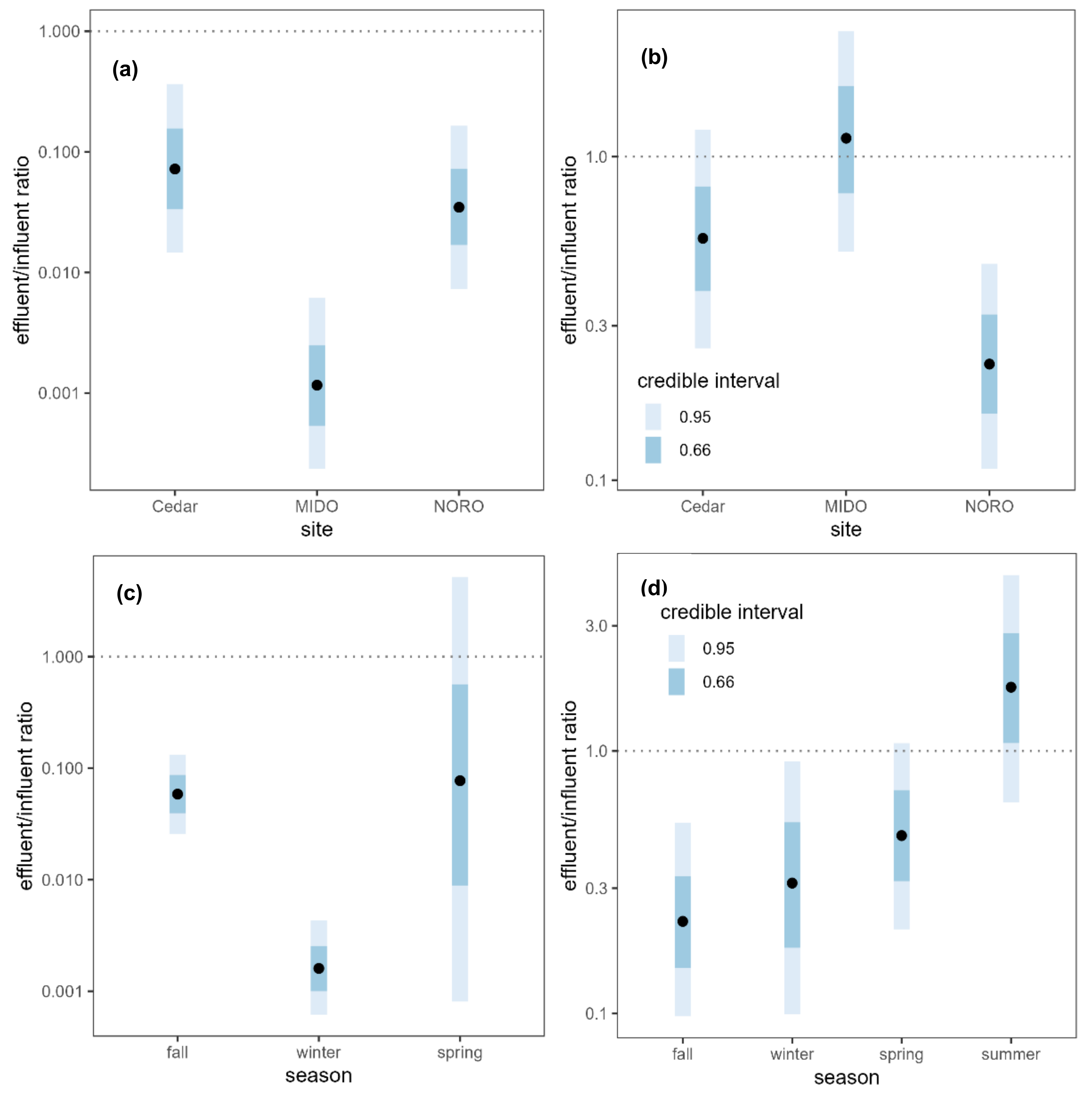
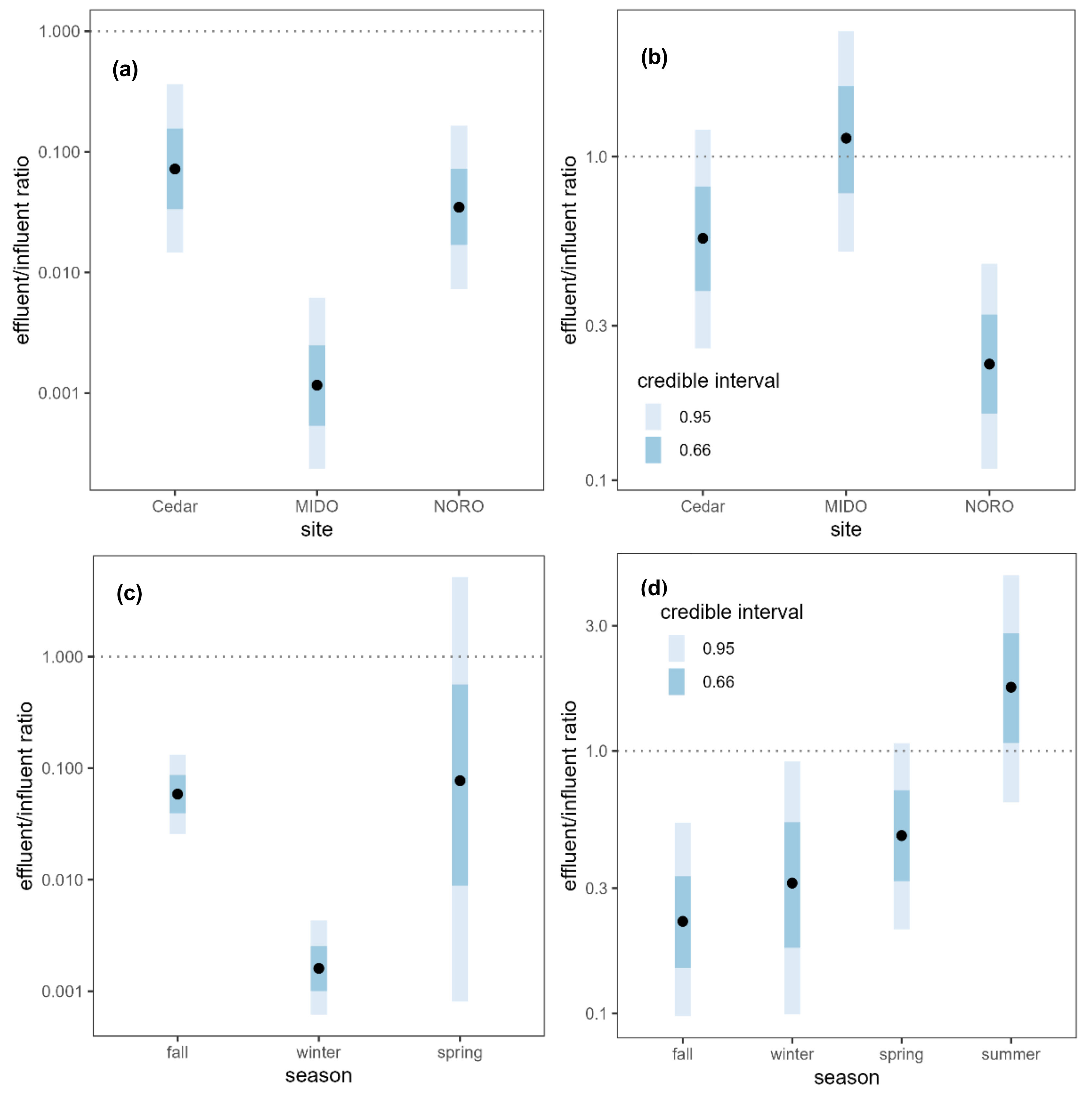
2.2. WWTPs as a Source of AR Contaminants
3. Discussion
3.1. Prevalence and Distribution of AR Contaminants in Surface Water
3.1.1. Antibiotic-Resistant Salmonella, E. coli, and Enterococcus
3.1.2. ESBL-Producing Enterobacteriaceae and CRE
3.1.3. Antibiotic Resistance Gene Markers
3.1.4. Antibiotics
3.2. WWTPs as a Source of AR Contaminants
4. Materials and Methods
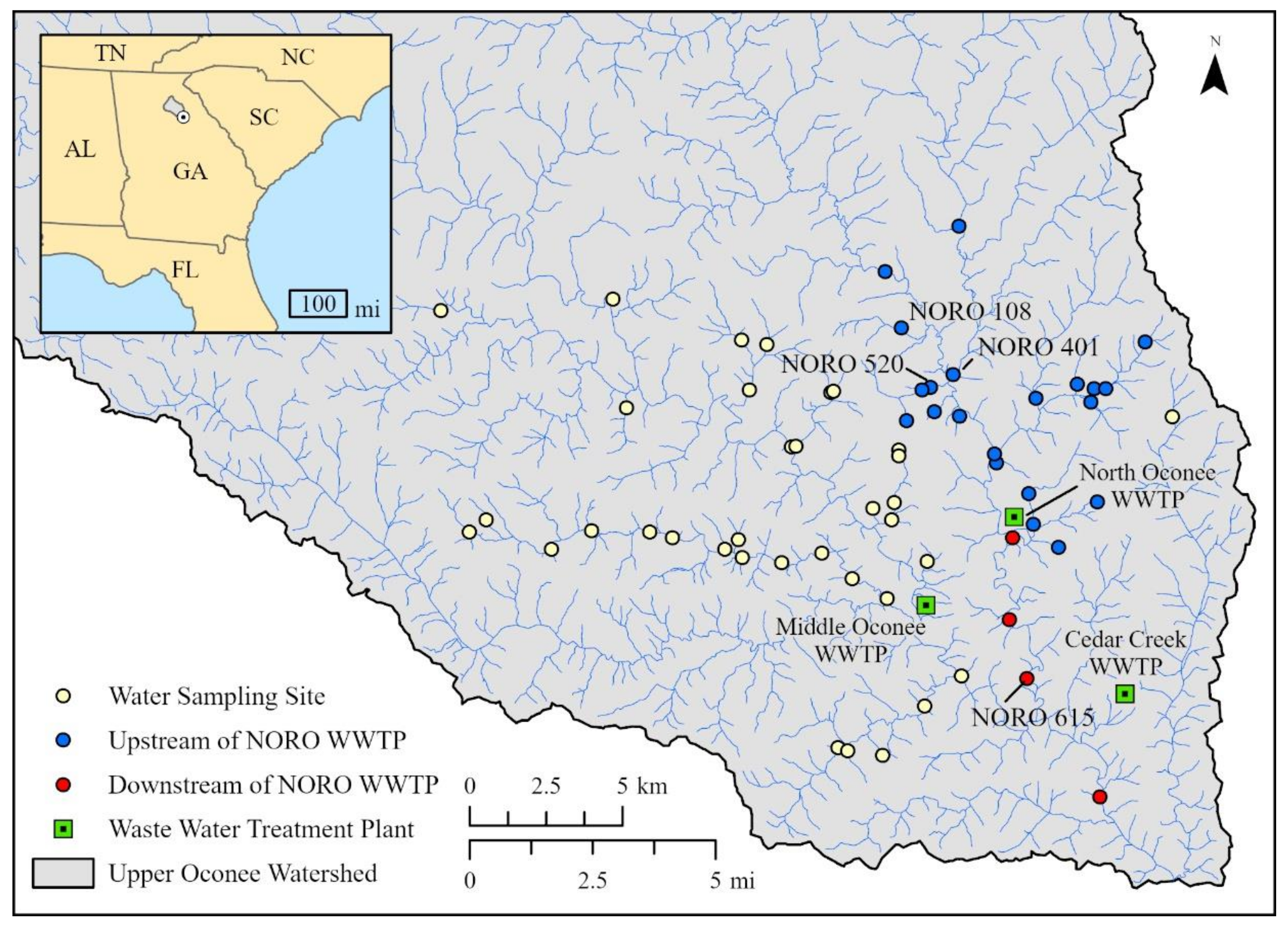
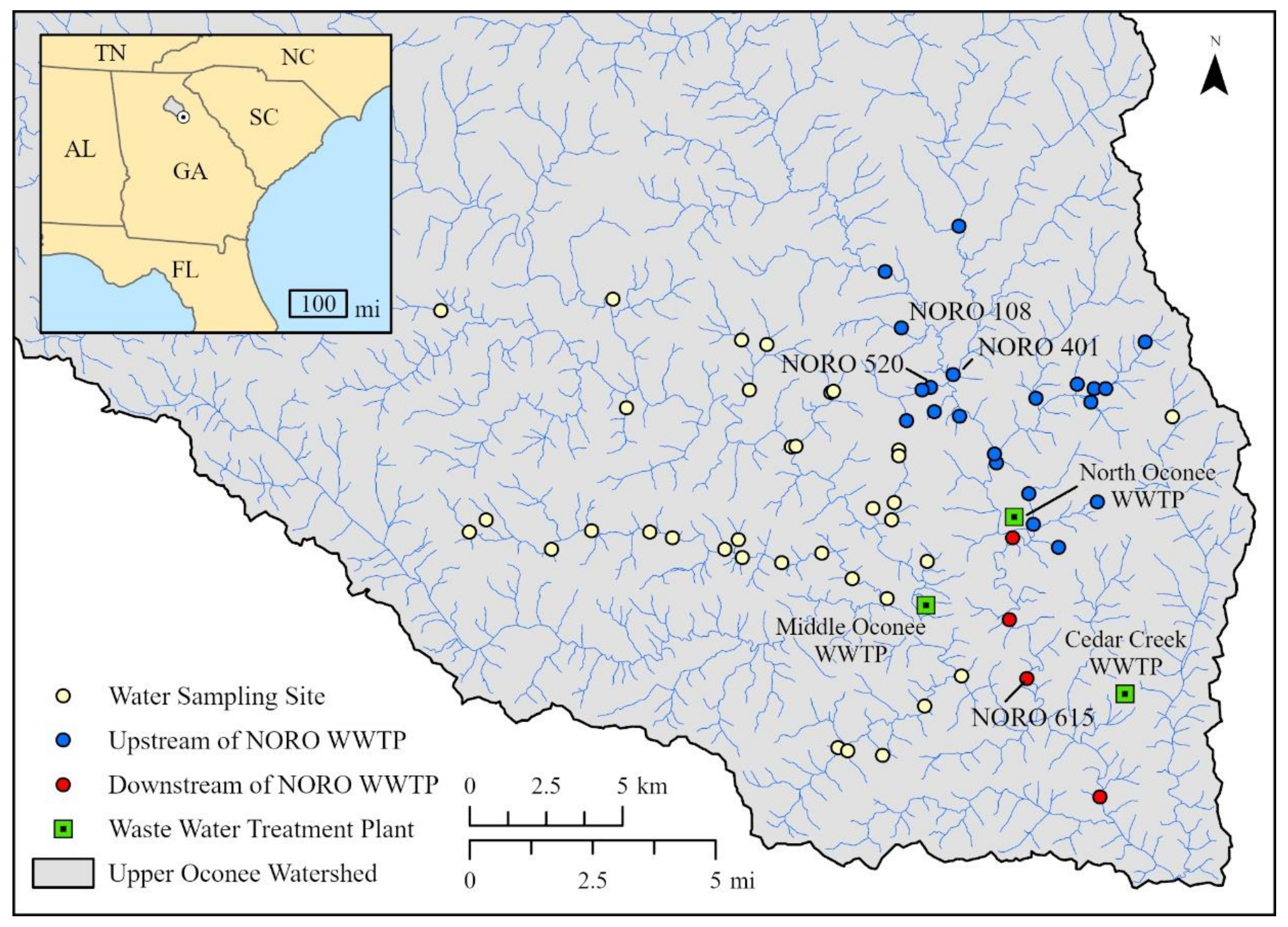
4.1. Surface Water and Wastewater Samples
4.2. Antibiotic-Resistant E. coli, Enterococcus, and Salmonella
4.2.1. Isolation and Identification of E. coli, Enterococcus, and Salmonella
4.2.2. Antimicrobial Susceptibility Testing
4.3. ESBL-Producing Enterobacteriaceae and CRE
4.3.1. Isolation and Identification of ESBL-Producing Enterobacteriaceae and CRE
4.3.2. Confirmation of ESBL Producers and CRE and Detection of β-Lactamase Genes
4.4. Antibiotic Resistance Gene Markers
4.5. Antibiotics
4.5.1. Antibiotics
4.5.2. Sample Preparation
4.5.3. LC-MS/MS Analysis
4.6. Statistical Analysis Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellington, E.M.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Kusi, J.; Ojewole, C.O.; Ojewole, A.E.; Nwi-Mozu, I. Antimicrobial Resistance Development Pathways in Surface Waters and Public Health Implications. Antibiotics 2022, 11, 821. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Kaur, R.; Verma, S.; Singh, S. Antimicrobials and Antibiotic Resistance Genes in Water Bodies: Pollution, Risk, and Control. Front. Environ. Sci. 2022, 10, 830861. [Google Scholar] [CrossRef]
- Cho, S.; Jackson, C.R.; Frye, J.G. Freshwater environment as a reservoir of extended-spectrum β-lactamase-producing Enterobacteriaceae. J. Appl. Microbiol. 2023, 134, lxad034. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, G. Detection and various environmental factors of antibiotic resistance gene horizontal transfer. Environ. Res. 2022, 212, 113267. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.S. Extended-spectrum-beta-lactamase, AmpC, and Carbapenemase issues. J. Clin. Microbiol. 2010, 48, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.D.; Laupland, K.B. Extended-spectrum β-lactamase-producing Enterobacteriaceae: An emerging public-health concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum beta-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Verlicchi, P.; Al Aukidy, M.; Zambello, E. Occurrence of pharmaceutical compounds in urban wastewater: Removal, mass load and environmental risk after a secondary treatment—A review. Sci. Total Environ. 2012, 429, 123–155. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Ying, G.G.; Pan, C.G.; Liu, Y.S.; Zhao, J.L. Comprehensive evaluation of antibiotics emission and fate in the river basins of China: Source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilback, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Fong, A.; Becket, E.; Yuan, J.; Tamae, C.; Medrano, L.; Maiz, M.; Wahba, C.; Lee, C.; Lee, K.; et al. Selective advantage of resistant strains at trace levels of antibiotics: A simple and ultrasensitive color test for detection of antibiotics and genotoxic agents. Antimicrob. Agents Chemother. 2011, 55, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, S.V.; Ostman, M.; Bengtsson-Palme, J.; Rutgersson, C.; Thoudal, M.; Sircar, T.; Blanck, H.; Eriksson, K.M.; Tysklind, M.; Flach, C.F.; et al. Minimal selective concentrations of tetracycline in complex aquatic bacterial biofilms. Sci. Total Environ. 2016, 553, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Jutkina, J.; Marathe, N.P.; Flach, C.F.; Larsson, D.G.J. Antibiotics and common antibacterial biocides stimulate horizontal transfer of resistance at low concentrations. Sci. Total Environ. 2018, 616–617, 172–178. [Google Scholar] [CrossRef]
- Jutkina, J.; Rutgersson, C.; Flach, C.F.; Joakim Larsson, D.G. An assay for determining minimal concentrations of antibiotics that drive horizontal transfer of resistance. Sci. Total Environ. 2016, 548–549, 131–138. [Google Scholar] [CrossRef]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.; Waldron, L.; Gillings, M.R. Potential impacts of aquatic pollutants: Sub-clinical antibiotic concentrations induce genome changes and promote antibiotic resistance. Front. Microbiol. 2015, 6, 803. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). National Antimicrobial Resistance Monitoring System—Enteric Bacteria (NARMS): 2004 Executive Report; U.S. Department of Health and Human Services, FDA: Rockville, MD, USA, 2008. [Google Scholar]
- Cho, S.; Hiott, L.M.; House, S.L.; Woodley, T.A.; McMillan, E.A.; Sharma, P.; Barrett, J.B.; Adams, E.S.; Brandenburg, J.M.; Hise, K.B.; et al. Analysis of Salmonella enterica Isolated from a Mixed-Use Watershed in Georgia, USA: Antimicrobial Resistance, Serotype Diversity, and Genetic Relatedness to Human Isolates. Appl. Environ. Microbiol. 2022, 88, e00393-22. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Hiott, L.M.; Woodley, T.A.; Frye, J.G.; Jackson, C.R. Evaluation of a new chromogenic agar for the detection of environmental Enterococcus. J. Microbiol. Methods 2020, 178, 106082. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Hiott, L.M.; McDonald, J.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Adams, E.S.; Frye, J.G.; Jackson, C.R. Diversity and antimicrobial resistance of Enterococcus from the Upper Oconee Watershed, Georgia. J. Appl. Microbiol. 2020, 128, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Nguyen, H.A.T.; McDonald, J.M.; Woodley, T.A.; Hiott, L.M.; Barrett, J.B.; Jackson, C.R.; Frye, J.G. Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA. Int. J. Environ. Res. Public. Health 2019, 16, 3761. [Google Scholar] [CrossRef] [PubMed]
- Raphael, E.; Wong, L.K.; Riley, L.W. Extended-spectrum Beta-lactamase gene sequences in gram-negative saprophytes on retail organic and nonorganic spinach. Appl. Environ. Microbiol. 2011, 77, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Tanner, W.D.; VanDerslice, J.A.; Goel, R.K.; Leecaster, M.K.; Fisher, M.A.; Olstadt, J.; Gurley, C.M.; Morris, A.G.; Seely, K.A.; Chapman, L.; et al. Multi-state study of Enterobacteriaceae harboring extended-spectrum beta-lactamase and carbapenemase genes in U.S. drinking water. Sci. Rep. 2019, 9, 3938. [Google Scholar] [CrossRef] [PubMed]
- Blaak, H.; van Hoek, A.H.; Veenman, C.; Docters van Leeuwen, A.E.; Lynch, G.; van Overbeek, W.M.; de Roda Husman, A.M. Extended spectrum ss-lactamase- and constitutively AmpC-producing Enterobacteriaceae on fresh produce and in the agricultural environment. Int. J. Food Microbiol. 2014, 168–169, 8–16. [Google Scholar] [CrossRef]
- Van Hoek, A.H.; Veenman, C.; van Overbeek, W.M.; Lynch, G.; de Roda Husman, A.M.; Blaak, H. Prevalence and characterization of ESBL- and AmpC-producing Enterobacteriaceae on retail vegetables. Int. J. Food Microbiol. 2015, 204, 1–8. [Google Scholar] [CrossRef]
- Kenzaka, T.; Tani, K. Draft Genome Sequence of Extended-Spectrum Beta-Lactamase-Producing Serratia fonticola BWK15 Isolated from Feces of Anas penelope. Genome Announc. 2017, 5, e01102-17. [Google Scholar] [CrossRef]
- Richter, L.; Du Plessis, E.M.; Duvenage, S.; Korsten, L. Occurrence, Identification, and Antimicrobial Resistance Profiles of Extended-Spectrum and AmpC beta-Lactamase-Producing Enterobacteriaceae from Fresh Vegetables Retailed in Gauteng Province, South Africa. Foodborne Pathog. Dis. 2019, 16, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Henriques, I.; Juca Ramos, R.T.; Barauna, R.A.; de Sa, P.H.; Marinho Almeida, D.; Carneiro, A.R.; Barbosa, S.; Pereira, A.; Alves, A.; Saavedra, M.J.; et al. Draft Genome Sequence of Serratia fonticola UTAD54, a Carbapenem-Resistant Strain Isolated from Drinking Water. Genome Announc. 2013, 1, e00970-13. [Google Scholar] [CrossRef] [PubMed]
- Olson, A.B.; Silverman, M.; Boyd, D.A.; McGeer, A.; Willey, B.M.; Pong-Porter, V.; Daneman, N.; Mulvey, M.R. Identification of a progenitor of the CTX-M-9 group of extended-spectrum beta-lactamases from Kluyvera georgiana isolated in Guyana. Antimicrob. Agents Chemother. 2005, 49, 2112–2115. [Google Scholar] [CrossRef] [PubMed]
- Humeniuk, C.; Arlet, G.; Gautier, V.; Grimont, P.; Labia, R.; Philippon, A. Beta-lactamases of Kluyvera ascorbata, probable progenitors of some plasmid-encoded CTX-M types. Antimicrob. Agents Chemother. 2002, 46, 3045–3049. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.M.; El Chakhtoura, N.G.; Patel, S.; Saade, E.; Donskey, C.J.; Bonomo, R.A.; Perez, F. Carbapenem-Resistant Enterobacter cloacae in Patients from the US Veterans Health Administration, 2006–2015. Emerg. Infect. Dis. 2017, 23, 878–880. [Google Scholar] [CrossRef] [PubMed]
- Annavajhala, M.K.; Gomez-Simmonds, A.; Uhlemann, A.C. Multidrug-Resistant Enterobacter cloacae Complex Emerging as a Global, Diversifying Threat. Front. Microbiol. 2019, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, Y.; Hagiya, H.; Akeda, Y.; Aye, M.M.; Myo Win, H.P.; Sakamoto, N.; Shanmugakani, R.K.; Takeuchi, D.; Nishi, I.; Ueda, A.; et al. Dissemination of carbapenemase-producing Enterobacteriaceae harbouring blaNDM or blaIMI in local market foods of Yangon, Myanmar. Sci. Rep. 2019, 9, 14455. [Google Scholar] [CrossRef] [PubMed]
- Boyd, D.A.; Mataseje, L.F.; Davidson, R.; Delport, J.A.; Fuller, J.; Hoang, L.; Lefebvre, B.; Levett, P.N.; Roscoe, D.L.; Willey, B.M.; et al. Enterobacter cloacae Complex Isolates Harboring blaNMC-A or blaIMI-Type Class A Carbapenemase Genes on Novel Chromosomal Integrative Elements and Plasmids. Antimicrob. Agents Chemother. 2017, 61, e02578-16. [Google Scholar] [CrossRef]
- Janecko, N.; Martz, S.L.; Avery, B.P.; Daignault, D.; Desruisseau, A.; Boyd, D.; Irwin, R.J.; Mulvey, M.R.; Reid-Smith, R.J. Carbapenem-Resistant Enterobacter spp. in Retail Seafood Imported from Southeast Asia to Canada. Emerg. Infect. Dis. 2016, 22, 1675–1677. [Google Scholar] [CrossRef]
- Brouwer, M.S.M.; Rapallini, M.; Geurts, Y.; Harders, F.; Bossers, A.; Mevius, D.J.; Wit, B.; Veldman, K.T. Enterobacter cloacae Complex Isolated from Shrimps from Vietnam Carrying blaIMI-1 Resistant to Carbapenems but Not Cephalosporins. Antimicrob. Agents Chemother. 2018, 62, e00398-18. [Google Scholar] [CrossRef]
- Aubron, C.; Poirel, L.; Ash Ronald, J.; Nordmann, P. Carbapenemase-producing Enterobacteriaceae, U.S. rivers. Emerg. Infect. Dis. 2005, 11, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Fedorka-Cray, P.J.; Jackson-Hall, M.C.; Hiott, L.M. Effect of media, temperature and culture conditions on the species population and antibiotic resistance of enterococci from broiler chickens. Lett. Appl. Microbiol. 2005, 41, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Damashek, J.; Westrich, J.R.; McDonald, J.M.B.; Teachey, M.E.; Jackson, C.R.; Frye, J.G.; Lipp, E.K.; Capps, K.A.; Ottesen, E.A. Non-point source fecal contamination from aging wastewater infrastructure is a primary driver of antibiotic resistance in surface waters. Water Res. 2022, 222, 118853. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Outpatient Antibiotic Prescriptions—United States. 2017. Available online: https://www.cdc.gov/antibiotic-use/community/pdfs/Annual-Report-2017-H.pdf (accessed on 2 July 2022).
- Centers for Disease Control and Prevention (CDC). Outpatient Antibiotic Prescriptions—United States. 2018. Available online: https://www.cdc.gov/antibiotic-use/community/pdfs/Annual-Report-2018-H.pdf (accessed on 2 July 2022).
- Stockton, B.; Jones, N. Antibiotics in Agriculture: The Blurred Line between Growth Promotion and Disease Prevention. Available online: https://www.thebureauinvestigates.com/stories/2018-09-19/growth-promotion-or-disease-prevention-the-loophole-in-us-antibiotic-regulations (accessed on 24 May 2022).
- Hanson, M.L.; Knapp, C.W.; Graham, D.W. Field assessment of oxytetracycline exposure to the freshwater macrophytes Egeria densa Planch. and Ceratophyllum demersum L. Environ. Pollut. 2006, 141, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, R.; Ternes, T.; Haberer, K.; Kratz, K.L. Occurrence of antibiotics in the aquatic environment. Sci. Total Environ. 1999, 225, 109–118. [Google Scholar] [CrossRef]
- Le Page, G.; Gunnarsson, L.; Snape, J.; Tyler, C.R. Integrating human and environmental health in antibiotic risk assessment: A critical analysis of protection goals, species sensitivity and antimicrobial resistance. Environ. Int. 2017, 109, 155–169. [Google Scholar] [CrossRef]
- Suda, K.J.; Hicks, L.A.; Roberts, R.M.; Hunkler, R.J.; Taylor, T.H. Trends and seasonal variation in outpatient antibiotic prescription rates in the United States, 2006 to 2010. Antimicrob. Agents Chemother. 2014, 58, 2763–2766. [Google Scholar] [CrossRef]
- Kummerer, K. Antibiotics in the aquatic environment—A review—Part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef]
- Kolpin, D.W.; Meyer, M.T. Pharmaceuticals, hormones, and other organic wastewater contaminants in U.S. streams, 1999–2000: A National Reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Carlson, K. Quantification of human and veterinary antibiotics in water and sediment using SPE/LC/MS/MS. Anal. Bioanal. Chem. 2007, 387, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Kleywegt, S.; Pileggi, V.; Yang, P.; Hao, C.; Zhao, X.; Rocks, C.; Thach, S.; Cheung, P.; Whitehead, B. Pharmaceuticals, hormones and bisphenol A in untreated source and finished drinking water in Ontario, Canada—Occurrence and treatment efficiency. Sci. Total Environ. 2011, 409, 1481–1488. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Larsson, D.G. Concentrations of antibiotics predicted to select for resistant bacteria: Proposed limits for environmental regulation. Environ. Int. 2016, 86, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Olivares-Rieumont, S.; Knapp, C.W.; Lima, L.; Werner, D.; Bowen, E. Antibiotic resistance gene abundances associated with waste discharges to the Almendares River near Havana, Cuba. Environ. Sci. Technol. 2011, 45, 418–424. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Y.; Wang, H.; Guo, C.; Qiu, H.; He, Y.; Zhang, Y.; Li, X.; Meng, W. Occurrence of antibiotics and antibiotic resistance genes in a sewage treatment plant and its effluent-receiving river. Chemosphere 2015, 119, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sanchez-Melsio, A.; Borrego, C.M.; Barcelo, D.; Balcazar, J.L. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef]
- An, X.L.; Su, J.Q.; Li, B.; Ouyang, W.Y.; Zhao, Y.; Chen, Q.L.; Cui, L.; Chen, H.; Gillings, M.R.; Zhang, T.; et al. Tracking antibiotic resistome during wastewater treatment using high throughput quantitative PCR. Environ. Int. 2018, 117, 146–153. [Google Scholar] [CrossRef]
- Alexander, J.; Hembach, N.; Schwartz, T. Evaluation of antibiotic resistance dissemination by wastewater treatment plant effluents with different catchment areas in Germany. Sci. Rep. 2020, 10, 8952. [Google Scholar] [CrossRef]
- Li, J.; Cheng, W.; Xu, L.; Strong, P.J.; Chen, H. Antibiotic-resistant genes and antibiotic-resistant bacteria in the effluent of urban residential areas, hospitals, and a municipal wastewater treatment plant system. Environ. Sci. Pollut. Res. Int. 2015, 22, 4587–4596. [Google Scholar] [CrossRef]
- Mao, D.; Yu, S.; Rysz, M.; Luo, Y.; Yang, F.; Li, F.; Hou, J.; Mu, Q.; Alvarez, P.J. Prevalence and proliferation of antibiotic resistance genes in two municipal wastewater treatment plants. Water Res. 2015, 85, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Burgmann, H.; Sorum, H.; Norstrom, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Sui, Q.; Zhang, J.; Tong, J.; Chen, M.; Wei, Y. Seasonal variation and removal efficiency of antibiotic resistance genes during wastewater treatment of swine farms. Environ. Sci. Pollut. Res. Int. 2017, 24, 9048–9057. [Google Scholar] [CrossRef] [PubMed]
- Osinska, A.; Korzeniewska, E.; Harnisz, M.; Felis, E.; Bajkacz, S.; Jachimowicz, P.; Niestepski, S.; Konopka, I. Small-scale wastewater treatment plants as a source of the dissemination of antibiotic resistance genes in the aquatic environment. J. Hazard. Mater. 2020, 381, 121221. [Google Scholar] [CrossRef]
- Jiao, Y.N.; Zhou, Z.C.; Chen, T.; Wei, Y.Y.; Zheng, J.; Gao, R.X.; Chen, H. Biomarkers of antibiotic resistance genes during seasonal changes in wastewater treatment systems. Environ. Pollut. 2018, 234, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Aydin, M.E.; Ulvi, A.; Kilic, H. Antibiotics in hospital effluents: Occurrence, contribution to urban wastewater, removal in a wastewater treatment plant, and environmental risk assessment. Environ. Sci. Pollut. Res. Int. 2019, 26, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Du, M.; Jiang, H.; Zhang, D.; Lin, L.; Ye, H.; Zhang, X. Occurrence, seasonal variation and removal efficiency of antibiotics and their metabolites in wastewater treatment plants, Jiulongjiang River Basin, South China. Environ. Sci. Process Impacts 2015, 17, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, Y.; Wang, N.; Wan, P.; Peng, Z.; Zhao, H.; Wang, W.; Xiong, L.; Zhang, S.; Liu, R. Seasonal variability of the correlation network of antibiotics, antibiotic resistance determinants, and bacteria in a wastewater treatment plant and receiving water. J. Environ. Manage 2022, 317, 115362. [Google Scholar] [CrossRef]
- Zielinski, W.; Korzeniewska, E.; Harnisz, M.; Drzymala, J.; Felis, E.; Bajkacz, S. Wastewater treatment plants as a reservoir of integrase and antibiotic resistance genes—An epidemiological threat to workers and environment. Environ. Int. 2021, 156, 106641. [Google Scholar] [CrossRef]
- Sabri, N.A.; Schmitt, H.; Van der Zaan, B.; Gerritsen, H.W.; Zuidema, T.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. J. Environ. Chem. Eng. 2020, 8, 102245. [Google Scholar] [CrossRef]
- Lekunberri, I.; Villagrasa, M.; Balcazar, J.L.; Borrego, C.M. Contribution of bacteriophage and plasmid DNA to the mobilization of antibiotic resistance genes in a river receiving treated wastewater discharges. Sci. Total Environ. 2017, 601–602, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Laird, E.; Gentry, T.J.; Brooks, J.P.; Karthikeyan, R. Increased Antimicrobial and Multidrug Resistance Downstream of Wastewater Treatment Plants in an Urban Watershed. Front. Microbiol. 2021, 12, 657353. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed]
- Capps, K.A.; Bateman McDonald, J.M.; Gaur, N.; Parsons, R. Assessing the Socio-Environmental Risk of Onsite Wastewater Treatment Systems to Inform Management Decisions. Environ. Sci. Technol. 2020, 54, 14843–14853. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Humayoun, S.B.; Adams, E.S.; Jackson, C.R.; Frye, J.G. Prevalence and characterization of Escherichia coli isolated from the Upper Oconee Watershed in Northeast Georgia. PLoS ONE 2018, 13, e0197005. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.; Greune, L.; Heusipp, G.; Karch, H.; Fruth, A.; Tschape, H.; Schmidt, M.A. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl. Environ. Microbiol. 2007, 73, 3380–3390. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B. Use of a genus- and species-specific multiplex PCR for identification of enterococci. J. Clin. Microbiol. 2004, 42, 3558–3565. [Google Scholar] [CrossRef] [PubMed]
- Leader, B.T.; Frye, J.G.; Hu, J.; Fedorka-Cray, P.J.; Boyle, D.S. High-throughput molecular determination of salmonella enterica serovars by use of multiplex PCR and capillary electrophoresis analysis. J. Clin. Microbiol. 2009, 47, 1290–1299. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI supplement M100; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Sjolund-Karlsson, M.; Joyce, K.; Blickenstaff, K.; Ball, T.; Haro, J.; Medalla, F.M.; Fedorka-Cray, P.; Zhao, S.; Crump, J.A.; Whichard, J.M. Antimicrobial susceptibility to azithromycin among Salmonella enterica isolates from the United States. Antimicrob. Agents Chemother. 2011, 55, 3985–3989. [Google Scholar] [CrossRef]
- Zhao, S.; White, D.G.; McDermott, P.F.; Friedman, S.; English, L.; Ayers, S.; Meng, J.; Maurer, J.J.; Holland, R.; Walker, R.D. Identification and expression of cephamycinase bla(CMY) genes in Escherichia coli and Salmonella isolates from food animals and ground meat. Antimicrob. Agents Chemother. 2001, 45, 3647–3650. [Google Scholar] [CrossRef]
- Bonnet, R.; Recule, C.; Baraduc, R.; Chanal, C.; Sirot, D.; De Champs, C.; Sirot, J. Effect of D240G substitution in a novel ESBL CTX-M-27. J. Antimicrob. Chemother. 2003, 52, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Fernando, D.M.; Tun, H.M.; Poole, J.; Patidar, R.; Li, R.; Mi, R.; Amarawansha, G.E.A.; Fernando, W.G.D.; Khafipour, E.; Farenhorst, A.; et al. Detection of Antibiotic Resistance Genes in Source and Drinking Water Samples from a First Nations Community in Canada. Appl. Environ. Microbiol. 2016, 82, 4767–4775. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, M.R.; Grant, J.M.; Plewes, K.; Roscoe, D.; Boyd, D.A. New Delhi metallo-beta-lactamase in Klebsiella pneumoniae and Escherichia coli, Canada. Emerg. Infect. Dis. 2011, 17, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Feria, C.; Ferreira, E.; Correia, J.D.; Goncalves, J.; Canica, M. Patterns and mechanisms of resistance to beta-lactams and beta-lactamase inhibitors in uropathogenic Escherichia coli isolated from dogs in Portugal. J. Antimicrob. Chemother. 2002, 49, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Colom, K.; Perez, J.; Alonso, R.; Fernandez-Aranguiz, A.; Larino, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of blaTEM, bla(SHV) and blaOXA-1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Brinas, L.; Zarazaga, M.; Saenz, Y.; Ruiz-Larrea, F.; Torres, C. Beta-lactamases in ampicillin-resistant Escherichia coli isolates from foods, humans, and healthy animals. Antimicrob. Agents Chemother. 2002, 46, 3156–3163. [Google Scholar] [CrossRef] [PubMed]
- Voets, G.M.; Fluit, A.C.; Scharringa, J.; Cohen Stuart, J.; Leverstein-van Hall, M.A. A set of multiplex PCRs for genotypic detection of extended-spectrum beta-lactamases, carbapenemases, plasmid-mediated AmpC beta-lactamases and OXA beta-lactamases. Int. J. Antimicrob. Agents 2011, 37, 356–359. [Google Scholar] [CrossRef]
- Yousefi, S.; Farajnia, S.; Nahaei, M.R.; Akhi, M.T.; Ghotaslou, R.; Soroush, M.H.; Naghili, B.; Jazani, N.H. Detection of metallo-beta-lactamase-encoding genes among clinical isolates of Pseudomonas aeruginosa in northwest of Iran. Diagn. Microbiol. Infect. Dis. 2010, 68, 322–325. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decre, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 9 August 2022).
- Stan Development Team. Stan Modeling Language Users Guide and Reference Manual, 2.28. 2021. Available online: https://mc-stan.org (accessed on 9 August 2022).
- Gabry, J.; Češnovar, R.; Johnson, A. cmdstanr: R Interface to ‘CmdStan’. 2022. Available online: https://mc-stan.org/cmdstanr/ (accessed on 9 August 2022).
- Bürkner, P.C. Advanced Bayesian Multilevel Modeling with the R Package brms. R J. 2018, 10, 395–411. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.8.2. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 9 August 2022).
- Kay, M. Tidybayes: Tidy Data and Geoms for Bayesian Models. R Package Version 3.0.2. 2022. Available online: http://mjskay.github.io/tidybayes/ (accessed on 9 August 2022).
- Wiley, J.; Hedeker, D. Brmsmargins: Bayesian Marginal Effects for ‘brms’ Models. R Package Version 0.2.0. 2022. Available online: https://CRAN.R-project.org/package=brmsmargins (accessed on 9 August 2022).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Season (No. of Samples) | Salmonella | E. coli | Enterococcus | |||
|---|---|---|---|---|---|---|---|
| % of Positive Samples (No. of Isolates) | No. of AR Isolates (No. of MDR Isolates) 1 | % of Positive Samples (No. of Isolates) | No. of AR Isolates (No. of MDR Isolates) 1 | % of Positive Samples (No. of Isolates) | No. of AR Isolates (No. of MDR Isolates) 1 | ||
| Surface water | 2017 fall (43) | 72.1 (78) | 27 (27) | 100 (43) | 3 (0) | 100 (43) | 43 (2) |
| 2018 winter (41) | 48.8 (57) | 0 (0) | 100 (41) | 4 (2) | 97.6 (40) | 40 (4) | |
| 2018 spring (42) | 59.5 (74) | 2 (0) | 100 (42) | 2 (1) | 100 (42) | 42 (1) | |
| 2018 summer (44) | 93.2 (94) | 0 (0) | 100 (44) | 2 (1) | 100 (44) | 44 (1) | |
| Influent | 2017 fall (3) | 100.0 (5) | 2 (0) | 100.0 (3) | 2 (2) | 100 (3) | 3 (0) |
| 2018 winter (3) | 100.0 (5) | 2 (0) | 100.0 (3) | 0 (0) | 100 (3) | 3 (1) | |
| 2018 spring (3) | 100.0 (7) | 2 (0) | 100 (3) | 0 (0) | 100 (3) | 3 (1) | |
| 2018 summer (3) | 100.0 (6) | 0 (0) | 100 (3) | 0 (0) | 100 (3) | 3 (0) | |
| Effluent | 2017 fall (3) | 0 (0) | 0 (0) | 66.7 (2) | 0 (0) | 100 (3) | 3 (1) |
| 2018 winter (2) | 0 (0) | 0 (0) | 100.0 (2) | 1 (1) | 100 (2) | 2 (0) | |
| 2018 spring (3) | 0 (0) | 0 (0) | 100 (3) | 1 (0) | 100 (3) | 3 (1) | |
| 2018 summer (3) | 33.3 (2) | 0 (0) | 66.7 (2) | 1 (1) | 66.7 (2) | 2 (1) | |
| Season | Source (No. of Samples) | Organisms | No. of β-Lactamase-Positive Enterobacteriaceae | No. of ESBL-Producing Enterobacteriaceae 1 | No. of CRE 1 |
|---|---|---|---|---|---|
| 2017 fall | water (42) | Escherichia coli | 3 | 2 | 0 |
| influent (3) | Enterobacter spp. | 2 | 0 | 1 | |
| effluent (3) | Escherichia coli | 1 | 1 | 0 | |
| 2018 winter | water (42) | Escherichia coli | 1 | 1 | 0 |
| Serratia fonticola | 19 | 19 | 2 | ||
| influent (3) | Citrobacter freundii | 1 | 0 | 0 | |
| Escherichia coli | 2 | 1 | 0 | ||
| effluent (3) | Enterobacter asburiae | 1 | 0 | 1 | |
| 2018 spring | water (41) | Escherichia coli | 3 | 2 | 0 |
| Klebsiella oxytoca | 1 | 0 | 0 | ||
| Klebsiella pneumoniae | 3 | 2 | 1 | ||
| Serratia fonticola | 40 | 1 | 2 | ||
| influent (3) | Escherichia coli | 1 | 1 | 0 | |
| Kluyvera ascorbata | 1 | 0 | 0 | ||
| Kluyvera cryocrescens | 2 | 2 | 0 | ||
| effluent (3) | Citrobacter braakii/freundii | 1 | 0 | 0 | |
| Klebsiella pneumoniae | 3 | 2 | 0 | ||
| 2018 summer | water (43) | Serratia fonticola | 38 | 0 | 2 |
| Enterobacter cloacae complex | 7 | 0 | 7 | ||
| Escherichia coli | 4 | 3 | 0 | ||
| Klebsiella pneumoniae | 1 | 1 | 0 | ||
| influent (3) | Enterobacter asburiae | 1 | 0 | 0 | |
| Enterobacter cloacae complex | 1 | 0 | 0 | ||
| Escherichia coli | 3 | 3 | 0 | ||
| Klebsiella pneumoniae | 3 | 1 | 0 | ||
| Kluyvera cryocrescens | 1 | 0 | 0 | ||
| effluent (3) | Enterobacter cloacae complex | 2 | 0 | 2 | |
| Klebsiella pneumoniae | 1 | 0 | 0 |
| Season | Source (No. of Samples) | ermB | tetB | blaKPC | ||||||
| No. of Positive Samples (%) | Maximum (Copies/mL) | Average (Copies/mL) | No. of Positive Samples (%) | Maximum (Copies/mL) | Average (Copies/mL) | No. of Positive Samples (%) | Maximum (Copies/mL) | Average (Copies/mL) | ||
| 2017 fall | surface water (38) | 23 (60.5) | 1533.8 | 57.2 | 10 (26.3) | 127.0 | 4.0 | 9 (23.7) | 13.5 | 1.6 |
| influent (3) | 3 (100) | 189,008.9 | 125,110.9 | 3 (100) | 4591.6 | 2524.8 | 3 (100) | 299,148.9 | 153,214.4 | |
| effluent (3) | 3 (100) | 33,052.9 | 11,394.3 | 3 (100) | 909.1 | 311.7 | 3 (100) | 31,100.5 | 11,630.9 | |
| 2018 winter | surface water (38) | 8 (21.6) | 355.0 | 11.3 | 2 (5.6) | 0.7 | 0.0 | 5 (13.5) | 49.9 | 2.0 |
| influent (3) | 3 (100) | 338,120.7 | 213,492.4 | 3 (100) | 15,694.5 | 8238.5 | 3 (100) | 362,358.8 | 142,942.9 | |
| effluent (3) | 3 (100) | 277.9 | 182.7 | 2 (66.7) | 5.3 | 2.1 | 3 (100) | 197.5 | 115.7 | |
| 2018 spring | surface water (34) | 8 (23.5) | 41.0 | 1.9 | 1 (2.9) | 1.6 | 0.0 | 2 (5.9) | 76.2 | 2.4 |
| influent (3) | 3 (100) | 98,933.2 | 46,165.2 | 3 (100) | 3219.8 | 1216.4 | 3 (100) | 73,007.5 | 48,650.3 | |
| 2018 summer | surface water (40) | 11 (27.5) | 347.5 | 11.9 | 2 (5.0) | 1.0 | 0.1 | 7 (17.5) | 377.5 | 9.8 |
| Season | Source (No. of Samples) | blaSHV | qnrS | blaCTX-M | ||||||
| No. of positive samples (%) | Maximum (copies/mL) | Average (copies/mL) | No. of positive samples (%) | Maximum (copies/mL) | Average (copies/mL) | No. of positive samples (%) | Maximum (copies/mL) | Average (copies/mL) | ||
| 2017 fall | surface water (38) | 9 (23.7) | 325.0 | 10.7 | 8 (21.1) | 308.4 | 12.5 | 3 (7.9) | 455.2 | 15.9 |
| influent (3) | 3 (100) | 37,087.9 | 24,498.7 | 3 (100) | 624,275.8 | 418,281.0 | 3 (100) | 6,740.0 | 2633.0 | |
| effluent (3) | 3 (100) | 7423.3 | 2637.1 | 3 (100) | 138,990.3 | 50,771.0 | 2 (66.7) | 1499.5 | 502.4 | |
| 2018 winter | surface water (38) | 2 (5.4) | 2.0 | 0.1 | 8 (21.1) | 582.6 | 19.5 | 0 (0.0) | 0.0 | 0.0 |
| influent (3) | 3 (100) | 40,126.5 | 26,100.6 | 3 (100) | 605,863.5 | 322,060.7 | 3 (100) | 26,050.7 | 13,449.5 | |
| effluent (3) | 3 (100) | 37.1 | 16.6 | 3 (100) | 1784.4 | 942.9 | 1 (33.3) | 51.8 | 17.3 | |
| 2018 spring | surface water (34) | 1 (2.9) | 2.0 | 0.1 | 3 (9.4) | 122.4 | 4.6 | 0 (0.0) | 0.0 | 0.0 |
| influent (3) | 3 (100) | 34,286.7 | 13,942.2 | 3 (100) | 291,963.4 | 137,383.9 | 3 (100) | 26,165.9 | 9461.5 | |
| 2018 summer | surface water (40) | 2 (5.0) | 7.7 | 0.2 | 8 (20.0) | 703.0 | 21.5 | 0 (0.0) | 0.0 | 0.0 |
| Season | Source (No. of Samples) | Antibiotics 1 | |||||||||||||
| AMX | AMP | AZI | AXO | TAZ | TIO | CIP | DAP | ERY | GEN | KAN | LIN | LNZ | |||
| 2017 fall | water (43) | maximum concentrations (ng/L) | 45.2 | 44.6 | 106.6 | 0.0 | 10.9 | 34.9 | 13.5 | 0.0 | 26.1 | 0.0 | 0.0 | 39.9 | 37.7 |
| average concentrations (ng/L) | 2.1 | 4.4 | 3.9 | 0.0 | 0.3 | 1.4 | 1.6 | 0.0 | 1.1 | 0.0 | 0.0 | 1.3 | 2.4 | ||
| no. of times detected | 14 | 15 | 7 | 0 | 1 | 21 | 28 | 0 | 19 | 0 | 0 | 14 | 9 | ||
| wastewater (6) | maximum concentrations (ng/L) | 2.1 | 52.8 | 120.6 | 0.0 | 0.0 | 38.0 | 123.7 | 16.3 | 20.8 | 0.0 | 0.0 | 1.3 | 0.6 | |
| average concentrations (ng/L) | 0.4 | 13.9 | 29.2 | 0.0 | 0.0 | 6.7 | 49.8 | 2.7 | 11.2 | 0.0 | 0.0 | 0.6 | 0.2 | ||
| no. of times detected | 1 | 5 | 6 | 0 | 0 | 3 | 6 | 1 | 5 | 0 | 0 | 6 | 3 | ||
| 2018 winter | water (30) | maximum concentrations (ng/L) | 14.6 | 41.0 | 0.4 | 6.6 | 152.3 | 19.9 | 1.8 | 1472.8 | 0.1 | 122.0 | 0.1 | 3.8 | 0.1 |
| average concentrations (ng/L) | 1.8 | 4.5 | 0.0 | 1.0 | 11.4 | 3.4 | 0.1 | 51.1 | 0.0 | 16.5 | 0.0 | 0.1 | 0.0 | ||
| no. of times detected | 8 | 6 | 2 | 6 | 4 | 11 | 8 | 5 | 4 | 13 | 1 | 3 | 3 | ||
| wastewater (6) | maximum concentrations (ng/L) | 91.0 | 18.3 | 22.9 | 21.9 | 686.0 | 37.8 | 4.6 | 1292.6 | 86.3 | 199.6 | 0.0 | 4.6 | 1.5 | |
| average concentrations (ng/L) | 15.2 | 3.1 | 6.8 | 3.6 | 162.0 | 15.5 | 0.8 | 237.1 | 23.0 | 48.1 | 0.0 | 1.4 | 0.3 | ||
| no. of times detected | 1 | 1 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 2 | 0 | 3 | 2 | ||
| 2018 spring | water (39) | maximum concentrations (ng/L) | 67.6 | 293.3 | 31.9 | 750.5 | 524.3 | 19.8 | 14.9 | 167.8 | 21.9 | 37.9 | 12.0 | 259.7 | 13.7 |
| average concentrations (ng/L) | 3.8 | 15.4 | 0.9 | 49.1 | 17.3 | 3.4 | 3.1 | 10.8 | 1.4 | 1.0 | 0.3 | 19.4 | 0.9 | ||
| no. of times detected | 12 | 14 | 19 | 6 | 3 | 17 | 19 | 7 | 14 | 1 | 1 | 12 | 18 | ||
| wastewater (6) | maximum concentrations (ng/L) | 6.9 | 216.9 | 46.7 | 0.0 | 0.0 | 14.7 | 67.1 | 0.0 | 113.2 | 0.0 | 0.0 | 59.0 | 4.5 | |
| average concentrations (ng/L) | 1.5 | 74.9 | 18.8 | 0.0 | 0.0 | 2.5 | 30.4 | 0.0 | 36.6 | 0.0 | 0.0 | 9.8 | 2.0 | ||
| no. of times detected | 2 | 4 | 5 | 0 | 0 | 1 | 4 | 0 | 3 | 0 | 0 | 1 | 4 | ||
| 2018 summer | water (39) | maximum concentrations (ng/L) | 39.9 | 4.6 | 26.9 | 8.6 | 23.8 | 12.7 | 13.6 | 2.9 | 66.5 | 23.7 | 24.9 | 0.1 | 27.3 |
| average concentrations (ng/L) | 12.6 | 0.6 | 6.3 | 0.5 | 1.8 | 1.0 | 1.9 | 0.3 | 13.2 | 3.0 | 1.9 | 0.0 | 5.8 | ||
| no. of times detected | 31 | 8 | 10 | 6 | 3 | 9 | 11 | 7 | 20 | 5 | 3 | 1 | 10 | ||
| wastewater (6) | maximum concentrations (ng/L) | 43.6 | 4.3 | 326.5 | 0.0 | 31.0 | 25.7 | 29.7 | 19.1 | 3550.6 | 23.8 | 75.7 | 0.0 | 69.2 | |
| average concentrations (ng/L) | 10.9 | 0.7 | 121.6 | 0.0 | 9.4 | 6.5 | 8.9 | 3.2 | 978.7 | 4.0 | 23.1 | 0.0 | 40.0 | ||
| no. of times detected | 2 | 1 | 3 | 0 | 2 | 2 | 2 | 1 | 3 | 1 | 3 | 0 | 6 | ||
| Season | Source (No. of Samples) | Antibiotics 1 | |||||||||||||
| MRP | MET | NAL | OXA | PEN | SMX | FIS | STR | TGC | TMP | TET | TYL | VAN | |||
| 2017 fall | water (43) | maximum concentrations (ng/L) | 19.6 | 31.5 | 13.6 | 80.9 | 40.9 | 66.7 | 25.4 | 44.9 | 0.0 | 40.9 | 0.0 | 22.1 | 0.0 |
| average concentrations (ng/L) | 0.9 | 2.1 | 0.6 | 10.1 | 2.4 | 3.2 | 1.3 | 6.4 | 0.0 | 2.9 | 0.0 | 1.4 | 0.0 | ||
| no. of times detected | 10 | 14 | 9 | 12 | 13 | 25 | 16 | 16 | 0 | 42 | 0 | 18 | 0 | ||
| wastewater (6) | maximum concentrations (ng/L) | 0.6 | 6.2 | 14.3 | 163.3 | 26.8 | 173.5 | 43.5 | 0.0 | 0.0 | 70.7 | 0.0 | 2.0 | 0.0 | |
| average concentrations (ng/L) | 0.1 | 1.6 | 4.1 | 37.0 | 5.4 | 72.6 | 10.0 | 0.0 | 0.0 | 29.2 | 0.0 | 0.6 | 0.0 | ||
| no. of times detected | 1 | 2 | 3 | 2 | 3 | 6 | 4 | 0 | 0 | 6 | 0 | 3 | 0 | ||
| 2018 winter | water (30) | maximum concentrations (ng/L) | 3.9 | 0.6 | 0.6 | 502.6 | 0.4 | 4.0 | 0.1 | 26.5 | 0.0 | 0.7 | 0.3 | 0.8 | 0.2 |
| average concentrations (ng/L) | 0.3 | 0.0 | 0.0 | 17.0 | 0.0 | 0.2 | 0.0 | 3.5 | 0.0 | 0.2 | 0.0 | 0.2 | 0.0 | ||
| no. of times detected | 6 | 6 | 2 | 2 | 1 | 7 | 10 | 5 | 0 | 20 | 4 | 20 | 1 | ||
| wastewater (6) | maximum concentrations (ng/L) | 0.0 | 2.1 | 0.2 | 10.6 | 0.0 | 35.0 | 0.0 | 28.2 | 0.0 | 91.5 | 2.8 | 91.6 | 0.0 | |
| average concentrations (ng/L) | 0.0 | 0.4 | 0.0 | 1.8 | 0.0 | 18.5 | 0.0 | 9.2 | 0.0 | 16.1 | 1.1 | 26.1 | 0.0 | ||
| no. of times detected | 0 | 1 | 1 | 1 | 0 | 6 | 0 | 2 | 0 | 4 | 4 | 6 | 0 | ||
| 2018 spring | water (39) | maximum concentrations (ng/L) | 220.7 | 50.0 | 13.7 | 593.2 | 19.7 | 29.0 | 60.5 | 159.3 | 46.5 | 16.6 | 44.8 | 63.6 | 238.0 |
| average concentrations (ng/L) | 8.1 | 4.3 | 0.4 | 23.6 | 3.9 | 1.2 | 4.7 | 6.9 | 3.5 | 4.0 | 15.8 | 6.0 | 18.4 | ||
| no. of times detected | 13 | 17 | 2 | 5 | 17 | 9 | 12 | 9 | 5 | 36 | 23 | 20 | 8 | ||
| wastewater (6) | maximum concentrations (ng/L) | 11.8 | 42.0 | 148.0 | 0.0 | 99.7 | 362.3 | 85.8 | 256.7 | 288.4 | 168.9 | 490.0 | 12.8 | 262.8 | |
| average concentrations (ng/L) | 2.7 | 8.2 | 25.8 | 0.0 | 28.3 | 170.1 | 23.0 | 62.9 | 84.9 | 100.1 | 173.5 | 3.3 | 112.1 | ||
| no. of times detected | 2 | 2 | 4 | 0 | 4 | 6 | 4 | 2 | 3 | 6 | 5 | 3 | 3 | ||
| 2018 summer | water (39) | maximum concentrations (ng/L) | 9.9 | 21.6 | 81.8 | 0.0 | 23.8 | 98.4 | 2.8 | 0.0 | 0.0 | 87.5 | 0.2 | 24.5 | 0.0 |
| average concentrations (ng/L) | 0.7 | 2.2 | 4.5 | 0.0 | 3.7 | 11.6 | 0.5 | 0.0 | 0.0 | 19.3 | 0.0 | 9.8 | 0.0 | ||
| no. of times detected | 4 | 12 | 4 | 0 | 12 | 32 | 11 | 0 | 0 | 37 | 1 | 33 | 0 | ||
| wastewater (6) | maximum concentrations (ng/L) | 31.8 | 28.2 | 328.5 | 0.0 | 0.0 | 2193.5 | 19.0 | 0.0 | 0.0 | 5777.0 | 20.9 | 134.0 | 0.0 | |
| average concentrations (ng/L) | 5.3 | 4.7 | 113.8 | 0.0 | 0.0 | 909.6 | 3.2 | 0.0 | 0.0 | 1228.0 | 3.7 | 55.2 | 0.0 | ||
| no. of times detected | 1 | 1 | 4 | 0 | 0 | 6 | 1 | 0 | 0 | 6 | 2 | 6 | 0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.; Hiott, L.M.; Read, Q.D.; Damashek, J.; Westrich, J.; Edwards, M.; Seim, R.F.; Glinski, D.A.; Bateman McDonald, J.M.; Ottesen, E.A.; et al. Distribution of Antibiotic Resistance in a Mixed-Use Watershed and the Impact of Wastewater Treatment Plants on Antibiotic Resistance in Surface Water. Antibiotics 2023, 12, 1586. https://doi.org/10.3390/antibiotics12111586
Cho S, Hiott LM, Read QD, Damashek J, Westrich J, Edwards M, Seim RF, Glinski DA, Bateman McDonald JM, Ottesen EA, et al. Distribution of Antibiotic Resistance in a Mixed-Use Watershed and the Impact of Wastewater Treatment Plants on Antibiotic Resistance in Surface Water. Antibiotics. 2023; 12(11):1586. https://doi.org/10.3390/antibiotics12111586
Chicago/Turabian StyleCho, Sohyun, Lari M. Hiott, Quentin D. Read, Julian Damashek, Jason Westrich, Martinique Edwards, Roland F. Seim, Donna A. Glinski, Jacob M. Bateman McDonald, Elizabeth A. Ottesen, and et al. 2023. "Distribution of Antibiotic Resistance in a Mixed-Use Watershed and the Impact of Wastewater Treatment Plants on Antibiotic Resistance in Surface Water" Antibiotics 12, no. 11: 1586. https://doi.org/10.3390/antibiotics12111586