
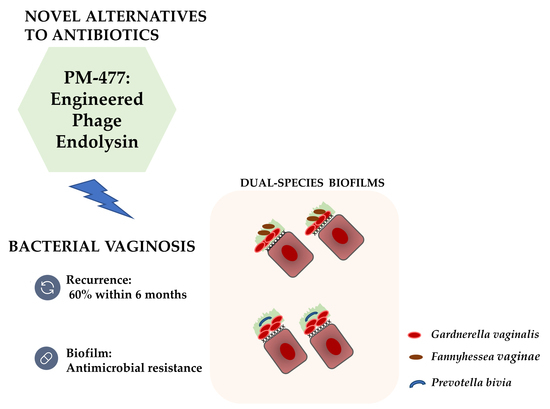
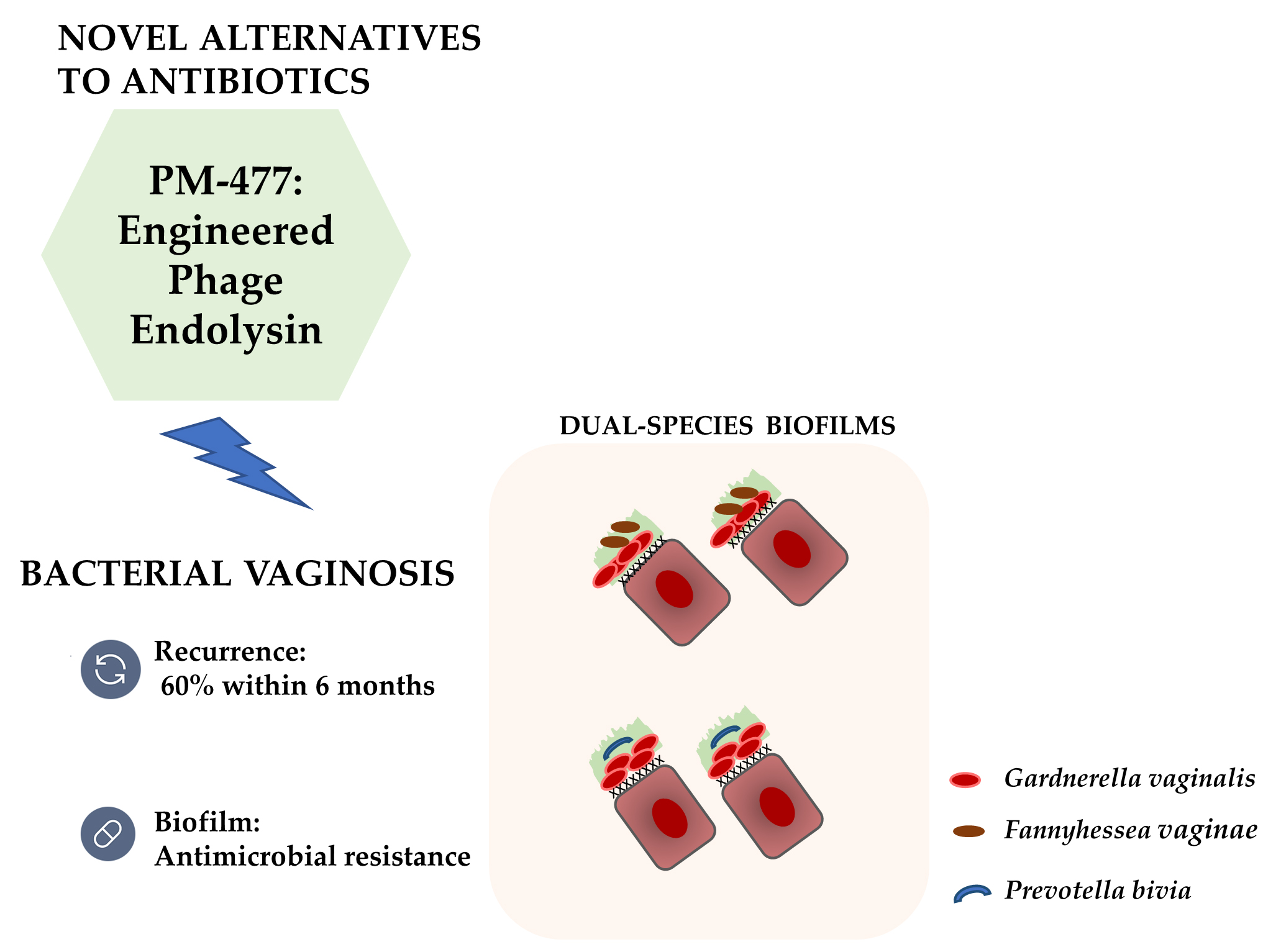
Exploiting the Anti-Biofilm Effect of the Engineered Phage Endolysin PM-477 to Disrupt In Vitro Single- and Dual-Species Biofilms of Vaginal Pathogens Associated with Bacterial Vaginosis

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Quantification of the Biomass of Single- and Dual BV-Associated Biofilms
2.2. Impact of PM-477 on Biofilm Cell Culturability
2.3. PM-477 Influences the Performance of PNA FISH
2.4. Prevalence of Bacterial Populations in the Dual-Species Biofilms before and after PM-477 Challenge
2.5. Indirect Assessment of PM-477 Activity in Biofilm Structure
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Preparation of PM-477
4.3. Dual-Species Biofilm Formation and EL Challenge
4.4. Quantification of Bacterial Biofilm by Crystal Violet (CV) Staining
4.5. Enumeration of Total Culturable Bacteria in the Biofilm Using the CFU Counting Approach
4.6. Impact of PM-477 on the Efficiency of PNA FISH Probes
4.7. Effect of PM-477 on the Bacterial Composition of Dual-Species Biofilms Assessed by Quantitative PCR (qPCR) Quantification
4.8. Confocal Laser Scanning Microscopy Analysis of Biofilm Bacterial Distribution
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reiter, S.; Spadt, S.K. Bacterial vaginosis: A primer for clinicians. Postgrad. Med. 2019, 131, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Rosca, A.S.; Castro, J.; Sousa, L.G.V.; Cerca, N. Gardnerella and vaginal health: The truth is out there. FEMS Microbiol. Rev. 2020, 44, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Isik, G.; Demirezen, Ş.; Dönmez, H.; Beksaç, M. Bacterial vaginosis in association with spontaneous abortion and recurrent pregnancy losses. J. Cytol. 2016, 33, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Dingens, A.S.; Fairfortune, T.S.; Reed, S.; Mitchell, C. Bacterial vaginosis and adverse outcomes among full-term infants: A cohort study. BMC Pregnancy Childbirth 2016, 16, 278. [Google Scholar] [CrossRef] [Green Version]
- Ravel, J.; Moreno, I.; Simón, C. Bacterial vaginosis and its association with infertility, endometritis, and pelvic inflammatory disease. Am. J. Obstet. Gynecol. 2021, 224, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Mckinnon, L.R.; Achilles, S.L.; Bradshaw, C.S.; Burgener, A.; Crucitti, T.; Fredricks, D.N.; Jaspan, H.B.; Kaul, R.; Kaushic, C.; Klatt, N.; et al. The Evolving Facets of Bacterial Vaginosis: Implications for HIV Transmission. AIDS Res. Hum. Retrovir. 2019, 35, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzny, C.A.; Taylor, C.M.; Swords, W.E.; Tamhane, A.; Chattopadhyay, D.; Cerca, N.; Schwebke, J.R. An updated conceptual model on the pathogenesis of bacterial vaginosis. J. Infect. Dis. 2019, 220, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.P.; Goodfellow, M.; Göker, M. Genome-based taxonomic classification of the phylum actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [Green Version]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Ladhoff, A.; Swidsinski, S.; Hale, L.P.; Lochs, H. Adherent Biofilms in Bacterial Vaginosis. Obstet. Gynecol. 2005, 106, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- De Backer, E.; Verhelst, R.; Verstraelen, H.; Alqumber, M.A.; Burton, J.P.; Tagg, J.R.; Temmerman, M.; Vaneechoutte, M. Quantitative determination by real-time PCR of four vaginal Lactobacillus species, Gardnerella vaginalis and Atopobium vaginae indicates an inverse relationship between L. gasseri and L. iners. BMC Microbiol. 2007, 7, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menard, J.P.; Mazouni, C.; Salem-Cherif, I.; Fenollar, F.; Raoult, D.; Boubli, L.; Gamerre, M.; Bretelle, F. High vaginal concentrations of Atopobium vaginae and Gardnerella vaginalis in women undergoing preterm labor. Obstet. Gynecol. 2010, 115, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Mendling, W.; Palmeira-de-Oliveira, A.; Biber, S.; Prasauskas, V. An update on the role of Atopobium vaginae in bacterial vaginosis: What to consider when choosing a treatment? A mini review. Arch. Gynecol. Obstet. 2019, 300, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, M.J.; Masztal, A.; Aldridge, K.E.; Fortenberry, J.D.; Fidel, P.L.; Martin, D.H. Association of Atopobium vaginae, a recently described metronidazole resistant anaerobe, with bacterial vaginosis. BMC Infect. Dis. 2004, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, C.; Amsel, R.; Eschenbach, D.; Schoenknecht, F.; Holmes, K. Anaerobic bacteria in nonspecific vaginitis. N. Engl. J. Med. 1980, 303, 601–607. [Google Scholar] [CrossRef]
- Gibbs, R.; McDuffi Jr, R.; Kunze, M.; Barr, J.M.; Wolf, D.; Sze, C.-I.; Shikes, R.; Sherman, M. Experimental intrauterine infection with Prevotella bivia in New Zealand White rabbits. Am. J. Obstet. Gynecol. 2004, 190, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Randis, T.M.; Ratner, A.J. Gardnerella and Prevotella: Co-Conspirators in the Pathogenesis of Bacterial Vaginosis. J. Infect. Dis. 2019, 220, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Bocking, A.; Hill, J.E.; Money, D.M. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome 2018, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- Muzny, C.A.; Blanchard, E.; Taylor, C.M.; Aaron, K.J.; Talluri, R.; Griswold, M.E.; Redden, D.T.; Luo, M.; Welsh, D.A.; Van Der Pol, W.J.; et al. Identification of Key Bacteria Involved in the Induction of Incident Bacterial Vaginosis: A Prospective Study. J. Infect. Dis. 2018, 218, 966–978. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.S.; Pirotta, M.; de Guingand, D.; Hocking, J.S.; Morton, A.N.; Garland, S.M.; Fehler, G.; Morrow, A.; Walker, S.; Vodstrcil, L.A.; et al. Efficacy of oral metronidazole with vaginal clindamycin or vaginal probiotic for bacterial vaginosis: Randomised placebo-controlled double-blind trial. PLoS ONE 2012, 7, e34540. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.S.; Morton, A.N.; Hocking, J.; Garland, S.M.; Morris, M.B.; Moss, L.M.; Horvath, L.B.; Kuzevska, I.; Fairley, C.K. High recurrence rates of bacterial vaginosis over the course of 12 months after oral metronidazole therapy and factors associated with recurrence. J. Infect. Dis. 2006, 193, 1478–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Swidsinski, S.; Dörffel, Y.; Scholze, J.; Lochs, H.; Verstraelen, H. An adherent Gardnerella vaginalis biofilm persists on the vaginal epithelium after standard therapy with oral metronidazole. Am. J. Obstet. Gynecol. 2008, 198, 97.e1–97.e6. [Google Scholar] [CrossRef]
- Swidsinski, A.; Dörffel, Y.; Loening-Baucke, V.; Schilling, J.; Mendling, W. Response of Gardnerella vaginalis biofilm to 5 days of moxifloxacin treatment. FEMS Immunol. Med. Microbiol. 2011, 61, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landlinger, C.; Tisakova, L.; Oberbauer, V.; Schwebs, T.; Muhammad, A.; Latka, A.; Van Simaey, L.; Vaneechoutte, M.; Guschin, A.; Resch, G.; et al. Engineered Phage Endolysin Eliminates Gardnerella Biofilm without Damaging Beneficial Bacteria in Bacterial Vaginosis Ex Vivo. Pathogens 2021, 10, 54. [Google Scholar] [CrossRef]
- Rosca, A.; Castro, J.; Sousa, L.; França, A.; Vanecchoute, M.; Cerca, N. In vitro interactions within a biofilm containing three species found in bacterial vaginosis support the higher antimicrobial tolerance associated with BV recurrence. J. Antimicrob. Chemother. 2022; in press. [Google Scholar]
- Rosca, A.S.; Castro, J.; Cerca, N. Evaluation of different culture media to support in vitro growth and biofilm formation of bacterial vaginosis-associated anaerobes. PeerJ 2020, 8, e9917. [Google Scholar] [CrossRef]
- Castro, J.; Rosca, A.S.; Cools, P.; Vaneechoutte, M.; Cerca, N. Gardnerella vaginalis Enhances Atopobium vaginae Viability in an in vitro Model. Front. Cell. Infect. Microbiol. 2020, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.; Azevedo, N.F.; Santos, S.; Keevil, C.W.; Vieira, M.J. Discriminating multi-species populations in biofilms with peptide nucleic acid fluorescence in situ hybridization (PNA FISH). PLoS ONE 2011, 6, e14786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.S.; Guimarães, N.; Madureira, P.; Azevedo, N.F. Optimization of a peptide nucleic acid fluorescence in situ hybridization (PNA-FISH) method for the detection of bacteria and disclosure of a formamide effect. J. Biotechnol. 2014, 187, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosca, A.S.; Castro, J.; França, Â.; Vaneechoutte, M.; Cerca, N. Gardnerella vaginalis Dominates Multi-Species Biofilms in Both Pre-Conditioned and Competitive In Vitro Biofilm Formation Models. Microb. Ecol. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Keer, J.T.; Birch, L. Molecular methods for the assessment of bacterial viability. J. Microbiol. Methods 2003, 53, 175–183. [Google Scholar] [CrossRef]
- Latka, A.; Van Simaey, L.; Reynders, M.; Cools, P.; Rogier, T.; Lebbe, B.; Corsini, L.; Landlinger, C.; Vaneechoutte, M. Optimization of Propidium Monoazide qPCR (Viability-qPCR) to Quantify the Killing by the Gardnerella-Specific Endolysin PM-477, Directly in Vaginal Samples from Women with Bacterial Vaginosis. Antibiotics 2022, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Muzny, C.A.; Kardas, P. A Narrative Review of Current Challenges in the Diagnosis and Management of Bacterial Vaginosis. Sex. Transm. Dis. 2020, 47, 441–446. [Google Scholar] [CrossRef]
- Bilardi, J.; Walker, S.; McNair, R.; Mooney-Somers, J.; Temple-Smith, M.; Bellhouse, C.; Fairley, C.; Chen, M.; Bradshaw, C. Women’s management of recurrent bacterial vaginosis and experiences of clinical care: A qualitative study. PLoS ONE 2016, 11, e0151794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redelinghuys, M.J.; Geldenhuys, J.; Jung, H.; Kock, M.M. Bacterial Vaginosis: Current Diagnostic Avenues and Future Opportunities. Front. Cell. Infect. Microbiol. 2020, 10, 354. [Google Scholar] [CrossRef]
- Verstraelen, H.; Swidsinski, A. The biofilm in bacterial vaginosis: Implications for epidemiology, diagnosis and treatment. Curr. Opin. Infect. Dis. 2013, 26, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.S.; Sobel, J.D. Current Treatment of Bacterial Vaginosis—Limitations and Need for Innovation. J. Infect. Dis. 2016, 214, S14–S20. [Google Scholar] [CrossRef] [Green Version]
- Marrazzo, J.M.; Dombrowski, J.C.; Wierzbicki, M.R.; Perlowski, C.; Pontius, A.; Dithmer, D.; Schwebke, J. Safety and efficacy of a novel vaginal anti-infective, TOL-463, in the treatment of bacterial vaginosis and vulvovaginal candidiasis: A randomized, single-blind, phase 2, controlled trial. Clin. Infect. Dis. 2019, 68, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Lima, Â.; Sousa, L.G.V.; Rosca, A.S.; Muzny, C.A.; Cerca, N. Crystal Violet Staining Alone Is Not Adequate to Assess Synergism or Antagonism in Multi-Species Biofilms of Bacteria Associated with Bacterial Vaginosis. Front. Cell. Infect. Microbiol. 2022, 11, 795797. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Mendling, W.; Dörffel, Y.; Schilling, J.; Halwani, Z.; Jiang, X.; Verstraelen, H.; Swidsinski, S. Infection through structured polymicrobial Gardnerella biofilms (StPM-GB). Histol. Histopathol. 2014, 29, 567–587. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Rosca, A.S.; Muzny, C.A.; Cerca, N. Atopobium vaginae and Prevotella bivia are able to incorporate and influence gene expression in a pre-formed Gardnerella vaginalis biofilm. Pathogens 2021, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.; Almeida, C.; Salgueiro, D.; Henriques, A.; Vaneechoutte, M.; Haesebrouck, F.; Vieira, M.J.; Rodrigues, L.; Azevedo, N.F.; Cerca, N. Fluorescence in situ Hybridization method using Peptide Nucleic Acid probes for rapid detection of Lactobacillus and Gardnerella spp. BMC Microbiol. 2013, 13, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, L.; Jespers, V.; Dahchour, N.; Mwambarangwe, L.; Musengamana, V.; Vaneechoutte, M.; Crucitti, T. Unravelling the Bacterial Vaginosis-Associated Biofilm: A Multiplex Gardnerella vaginalis and Atopobium vaginae Fluorescence In Situ Hybridization Assay Using Peptide Nucleic Acid Probes. PLoS ONE 2015, 10, e0136658. [Google Scholar] [CrossRef] [PubMed]
- Bourne, R. ImageJ. In Fundamentals of Digital Imaging in Medicine; Springer: London, UK, 2010; Volume 9, pp. 185–188. [Google Scholar] [CrossRef]
- Magalhães, A.P.; França, Â.; Pereira, M.O.; Cerca, N. RNA-based qPCR as a tool to quantify and to characterize dual-species biofilms. Sci. Rep. 2019, 9, 13639. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Probe Efficiency (%) |
|---|---|
| G. vaginalis biofilm | 92.08 * |
| G. vaginalis biofilm after PM-477 exposure (16 µL/mL) | 38.90 * |
| F. vaginae biofilm | 91.59 |
| F. vaginae biofilm after PM-477 exposure (16 µL/mL) | 89.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, J.; Sousa, L.G.V.; França, Â.; Podpera Tisakova, L.; Corsini, L.; Cerca, N. Exploiting the Anti-Biofilm Effect of the Engineered Phage Endolysin PM-477 to Disrupt In Vitro Single- and Dual-Species Biofilms of Vaginal Pathogens Associated with Bacterial Vaginosis. Antibiotics 2022, 11, 558. https://doi.org/10.3390/antibiotics11050558
Castro J, Sousa LGV, França Â, Podpera Tisakova L, Corsini L, Cerca N. Exploiting the Anti-Biofilm Effect of the Engineered Phage Endolysin PM-477 to Disrupt In Vitro Single- and Dual-Species Biofilms of Vaginal Pathogens Associated with Bacterial Vaginosis. Antibiotics. 2022; 11(5):558. https://doi.org/10.3390/antibiotics11050558
Chicago/Turabian StyleCastro, Joana, Lúcia G. V. Sousa, Ângela França, Lenka Podpera Tisakova, Lorenzo Corsini, and Nuno Cerca. 2022. "Exploiting the Anti-Biofilm Effect of the Engineered Phage Endolysin PM-477 to Disrupt In Vitro Single- and Dual-Species Biofilms of Vaginal Pathogens Associated with Bacterial Vaginosis" Antibiotics 11, no. 5: 558. https://doi.org/10.3390/antibiotics11050558