
Artificial Cell Encapsulation for Biomaterials and Tissue Bio-Nanoengineering: History, Achievements, Limitations, and Future Work for Potential Clinical Applications and Transplantation


 , ,
, ,  , , , ,
, , , ,  ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology
3. Diabetes Mellitus
3.1. Overview
3.2. Treatments and Limitations
4. β-Cell Biology and Mechanism of Insulin Release
5. Insulinoma Cell Lines and Their Derivatives
6. Islet Transplantation
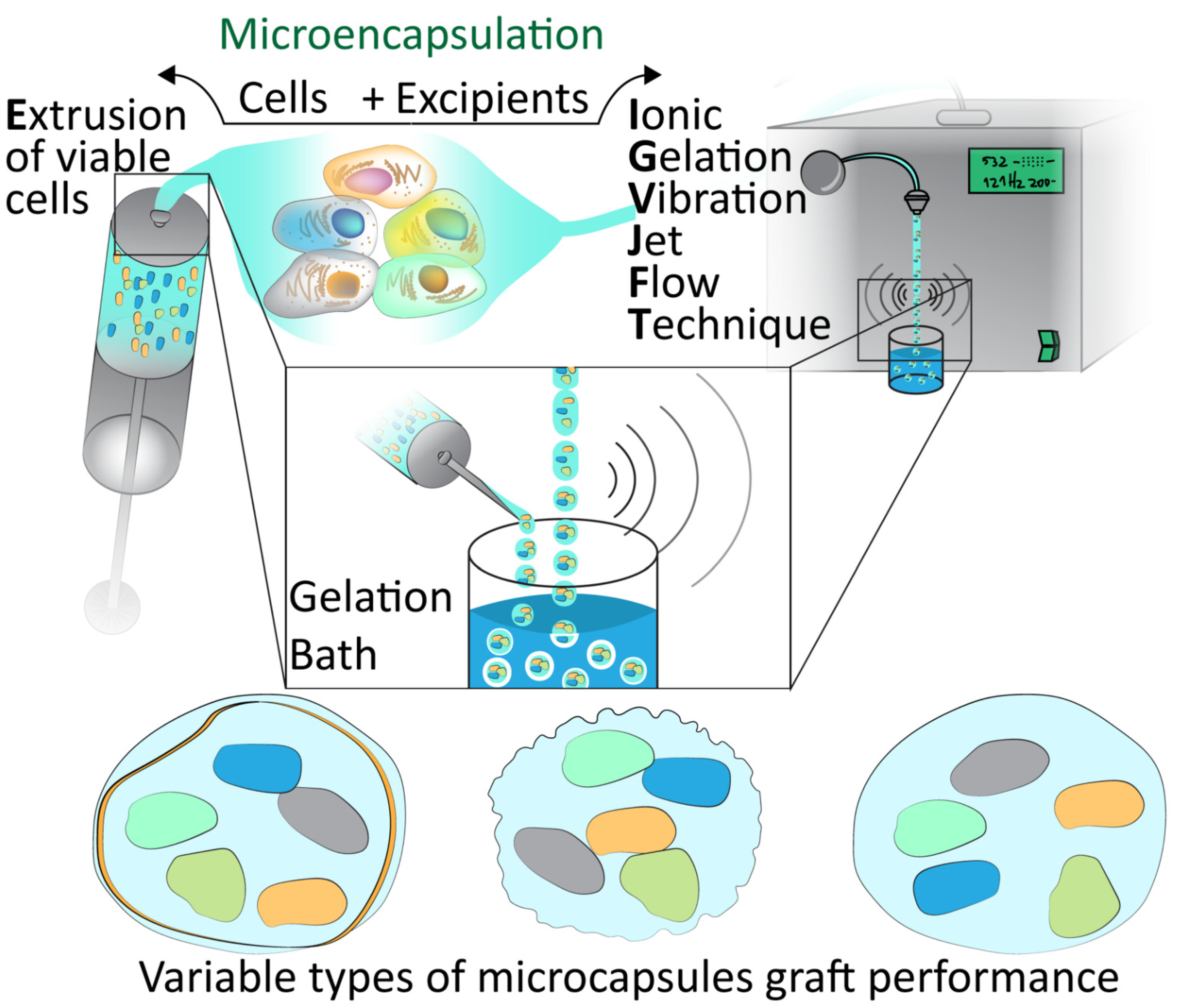
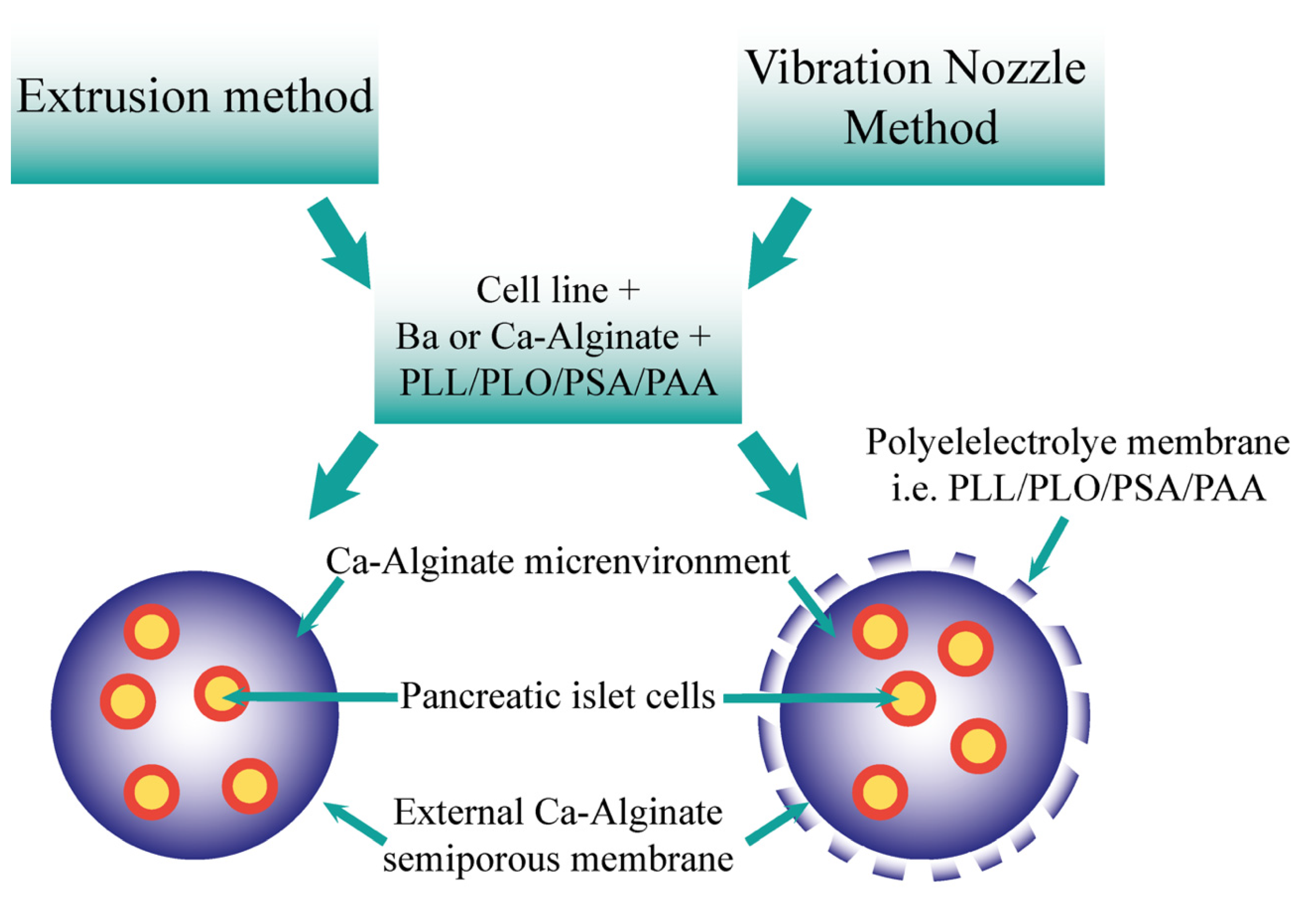
7. Cell Microencapsulation
8. Vibrational Nozzle Method
9. Islet Encapsulation and Current Progress
10. Limitations
10.1. Pharmaceutical Drawbacks of Microencapsulation
10.2. In Vivo Limitations
11. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Codario, R. Pathophysiology of Type-2 Diabetes. In Type 2 Diabetes, Pre-Diabetes, and the Metabolic Syndrome; Humana Press: Totowa, NJ, USA, 2011; pp. 1–14. [Google Scholar] [CrossRef]
- Nolan, C.J.; Damm, P.; Prentki, M. Type 2 diabetes across generations: From pathophysiology to prevention and management. Lancet 2011, 378, 169–181. [Google Scholar] [CrossRef]
- Barbeau, W.E.; Bassaganya-Riera, J.; Hontecillas, R. Putting the pieces of the puzzle together—A series of hypotheses on the etiology and pathogenesis of type 1 diabetes. Med. Hypotheses 2007, 68, 607–619. [Google Scholar] [CrossRef]
- Fakhoury, M.; Negrulj, R.; Mooranian, A.; Al-Salami, H. Inflammatory bowel disease: Clinical aspects and treatments. J. Inflamm. Res. 2014, 7, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.S.; Epstein, S.; Corkey, B.E.; Grant, S.F.A.; Gavin, R.J.R.; Aguilar, R.B. The Time Is Right for a New Classification System for Diabetes: Rationale and Implications of the β-Cell-Centric Classification Schema. Diabetes Care 2016, 39, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.Y.; Colagiuri, S. Diagnostic Criteria and Classification. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment; Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–17. [Google Scholar] [CrossRef]
- Ligtenberg, J.J.; Reitsma, W.D.; Van Haeften, T.W. Gliclazide mainly affects insulin secretion in second phase of type 2 diabetes mellitus. Horm. Metab. Res. 2001, 33, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Leitao, C.B.; Bernetti, K.; Tharavanij, T.; Cure, P.; Ricordi, C.; Alejandro, R. Type 2 diabetes mellitus phenotype and graft survival after islet transplantation. Transplantation 2009, 88, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Holden, S.E.; Jenkins-Jones, S.; Morgan, C.L.; Schernthaner, G.; Currie, C.J. Glucose-lowering with exogenous insulin monotherapy in type 2 diabetes: Dose association with all-cause mortality, cardiovascular events and cancer. Diabetes Obes. Metab. 2014, 17, 350–362. [Google Scholar] [CrossRef]
- Moore, P.A.; Zgibor, J.C.; Dasanayake, A.P. Diabetes: A growing epidemic of all ages. J. Am. Dent. Assoc. 2003, 134, 11S–15S. [Google Scholar] [CrossRef]
- Bas, V.N.; Cetinkaya, S.; Agladioglu, S.Y.; Kendirici, H.N.; Bilgili, H.; Yildirim, N.; Aycan, Z. Insulin oedema in newly diagnosed type 1 diabetes mellitus. J. Clin. Res. Pediatric Endocrinol. 2010, 2, 46–48. [Google Scholar] [CrossRef]
- Moser, A.; Hsu, H.T.; van Endert, P. Beta cell antigens in type 1 diabetes: Triggers in pathogenesis and therapeutic targets. F1000 Biol. Rep. 2010, 2, 75. [Google Scholar] [CrossRef]
- Schober, E.; Rami, B. Dermatological side effects and complications of continuous subcutaneous insulin infusion in preschool-age and school-age children. Pediatric Diabetes 2009, 10, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Fleury-Milfort, E. Insulin replacement therapy. Minimizing complications and side effects. Adv. Nurse Pract. 2008, 16, 39–40. [Google Scholar]
- Consoli, A. Insulin Treatment. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment; Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–24. [Google Scholar]
- Dominguez-Bendala, J.; Pileggi, A.; Ricordi, C. Islet cell therapy and pancreatic stem cells. In Handbook of Stem Cells, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 2, pp. 835–853. [Google Scholar] [CrossRef]
- Efrat, S. Beta-cell replacement for insulin-dependent diabetes mellitus. Adv. Drug Deliv. Rev. 2008, 60, 114–123. [Google Scholar] [CrossRef]
- Poggioli, R.; Faradji, R.N.; Ponte, G.; Betancourt, A.; Messinger, S.; Baidal, D.A.; Froud, T.; Ricordi, C.; Alejandro, R. Quality of Life After Islet Transplantation. Arab. Archaeol. Epigr. 2006, 6, 371–378. [Google Scholar] [CrossRef]
- Gangemi, A.; Salehi, P.; Hatipoglu, B.; Martellotto, J.; Barbaro, B.; Kuechle, J.B.; Qi, M.; Wang, Y.; Pallan, P.; Owens, C.; et al. Islet transplantation for brittle type 1 diabetes: The UIC protocol. Am. J. Transpl. 2008, 8, 1250–1261. [Google Scholar] [CrossRef]
- Clatworthy, M.; Watson, C.; Allison, M.; Dark, J. Transplantation at a Glance; John Wiley & Sons, Incorporated: Somerset, UK, 2012. [Google Scholar]
- Froud, T.; Ricordi, C.; Baidala, D.A.; Hafiz, M.M.; Ponte, G.; Cure, P.; Pileggi, A.; Poggioli, R.; Ichii, H.; Khan, A.; et al. Islet Transplantation in Type 1 Diabetes Mellitus Using Cultured Islets and Steroid-Free Immunosuppression: Miami Experience. Am. J. Transpl. 2005, 5, 2037–2046. [Google Scholar] [CrossRef]
- Shapiro, A.M.; Gallant, H.L.; Hao, E.G.; Lakey, J.R.; McCready, T.; Rajotte, R.V.; Yatscoff, R.W.; Kneteman, N.M. The portal immunosuppressive storm: Relevance to islet transplantation? Ther. Drug Monit. 2005, 27, 35–37. [Google Scholar] [CrossRef]
- Shapiro, A.M.; Ricordi, C.; Hering, B.J.; Auchincloss, H.; Lindblad, R.; Robertson, R.P.; Secchi, A.; Brendel, M.D.; Berney, T.; Brennan, D.C.; et al. International trial of the Edmonton protocol for islet transplantation. N. Engl. J. Med. 2006, 355, 1318–1330. [Google Scholar] [CrossRef] [Green Version]
- Piemonti, L.; Socci, C.; Nano, R.; Maffi, P.; Secchi, A. Islet Cell or Pancreas Transplantation. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment, Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–40. [Google Scholar] [CrossRef]
- Pareta, R.A.; Farney, A.; Opara, E.C. Design of a Bioartificial Pancreas. Pathobiology 2013, 80, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Pareta, R.; McQuilling, J.P.; Farney, A.C.; Opara, E.C. Bioartificial Pancreas: Evaluation of Crucial Barriers to Clinical Application. In Organ Donation and Transplantation-Public Policy and Clinical Perspectives; IntechOpen Limited: London, UK, 2012; pp. 239–266. [Google Scholar]
- de Vos, P.; Faas, M.M.; Strand, B.; Calafiore, R. Alginate-based microcapsules for immunoisolation of pancreatic islets. Biomaterials 2006, 27, 5603–5617. [Google Scholar] [CrossRef] [PubMed]
- Eramura, Y.; Iwata, H. Bioartificial pancreas: Microencapsulation and conformal coating of islet of Langerhans. Adv. Drug Deliv. Rev. 2010, 62, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Rokstad, A.M.A.; Lacík, I.; de Vos, P.; Strand, B.L. Advances in biocompatibility and physico-chemical characterization of microspheres for cell encapsulation. Adv. Drug Deliv. Rev. 2014, 67–68, 111–130. [Google Scholar] [CrossRef]
- Desai, T.A.; Chu, W.H.; Rasi, G.; Vallebona, P.S.; Guarino, E.; Ferrari, M. Microfabricated Biocapsules Provide Short-Term Immunoisolation of Insulinoma Xenografts. Biomed. Microdevices 1999, 1, 131–138. [Google Scholar] [CrossRef]
- Desai, T.A.; Chu, W.H.; Ferrari, M.; Rasi, G.; Sinibaldi-Vallebona, P.; Borboni, P.; Beattie, G.M.; Hayek, A. Implantation of microfabricated immunoisolating biocapsules. In Micro-and Nanofabricated Structures and Devices for Biomedical Environmental Applications; SPIE: Bellingham, WA, USA, 1998; Volume 3258, pp. 40–47. [Google Scholar]
- Lopez-Mendez, T.B.; Santos-Vizcaino, E.; Pedraz, J.L.; Orive, G.; Hernandez, R.M. Cell microencapsulation technologies for sustained drug delivery: Latest advances in efficacy and biosafety. J. Control. Release 2021, 335, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Kumar, R.; Laloo, D.; Hemalatha, S. Diabetes mellitus: An overview on its pharmacological aspects and reported medicinal plants having antidiabetic activity. Asian Pac. J. Trop. Biomed. 2012, 2, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Ley, S.H.; Meigs, J.B. Epidemiology and Risk Factors of Type 2 Diabetes. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment; Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–26. [Google Scholar] [CrossRef]
- Guglielmi, C.; Leslie, R.D.; Pozzilli, P. Epidemiology and Risk Factors of Type 1 Diabetes. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment; Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–15. [Google Scholar] [CrossRef]
- Federation, I.D. IDF Diabetes Atlas. IDF Diabetes Atlas. Brussels, Belgium: International Diabetes Federation; International Diabetes Federation (IDF): Brussels, Belgium, 2019; Volume 9. [Google Scholar]
- Coppieters, K.T.; Roep, B.O.; von Herrath, M.G. Beta cells under attack: Toward a better understanding of type 1 diabetes immunopathology. Semin. Immunopathol. 2011, 33, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pickup, J.C. Insulin Pumps. In Diabetes. Epidemiology, Genetics, Pathogenesis, Diagnosis, Prevention, and Treatment, Bonora, E., DeFronzo, R., Eds.; Springer International Publishing: Cham, Denmark, 2018; pp. 1–13. [Google Scholar] [CrossRef]
- Rickels, M.R.; Robertson, R.P. Pancreatic Islet Transplantation in Humans: Recent Progress and Future Directions. Endocr. Rev. 2019, 40, 631–668. [Google Scholar] [CrossRef] [Green Version]
- Leitao, C.; Peixoto, E.M.L.; Westphalen, A.C.; Mireles-Zavala, L.G.; Lauriola, V.; Bernetti, K.; Corrales, A.; Ricordi, C.; Alejandro, R. Liver Fat Accumulation after Islet Transplantation and Graft Survival. Cell Transplant. 2014, 23, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaithilingam, V.; Tuch, B.E. Islet Transplantation and Encapsulation: An Update on Recent Developments. Rev. Diabet. Stud. 2011, 8, 51–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrinpar, A.; Loomba, R. Review article: The emerging interplay among the gastrointestinal tract, bile acids and incretins in the pathogenesis of diabetes and non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2012, 36, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Chiang, J.Y. Bile Acid signaling in liver metabolism and diseases. J. Lipids 2012, 2012, 754067. [Google Scholar] [CrossRef] [Green Version]
- Mikov, M.; Al-Salami, H.; Golocorbin-Kon, G. Potentials and Limitations of Bile Acids and Probiotics in Diabetes Mellitus; IntechOpen Limited: London, UK, 2012; pp. 365–402. [Google Scholar]
- Al-Kassas, R.S.; Al-Gohary, O.M.; Al-Faadhel, M.M. Controlling of systemic absorption of gliclazide through incorporation into alginate beads. Int. J. Pharm. 2007, 341, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Hamaguchi, M.; Kaji, A.; Sakai, R.; Osaka, T.; Inoue, R.; Kashiwagi, S.; Mizushima, K.; Uchiyama, K.; Takagi, T.; et al. Intake of sucrose affects gut dysbiosis in patients with type 2 diabetes. J. Diabetes Investig. 2020, 11, 1623–1634. [Google Scholar] [CrossRef] [PubMed]
- Prawitt, J.; Caron, S.; Staels, B. Bile Acid Metabolism and the Pathogenesis of Type 2 Diabetes. Curr. Diabetes Rep. 2011, 11, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R. Type 2 diabetes: Etiology and reversibility. Diabetes Care 2013, 36, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Negrulj, R.; Mooranian, A.; Al-Salami, H. Potentials and Limitations of Bile Acids in Type 2 Diabetes Mellitus: Applications of Microencapsulation as a Novel Oral Delivery System. J. Endocrinol. Diabetes Mellit. 2013, 1, 49–59. [Google Scholar]
- Chang, T.M.S. Semipermeable Microcapsules. Science 1964, 146, 524–525. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.M.S. Artificial Cells: Biotechnology, Nanomedicine, Regenerative Medicine, Blood Substitutes, Bioencapsulation, and Cell/Stem Cell Therapy; World Scientific: Singapore, 2007. [Google Scholar] [CrossRef]
- Mooranian, A.; Negrulj, R.; Al-Sallami, H.S.; Fang, Z.; Mikov, M.; Golocorbin-Kon, S.; Fakhoury, M.; Aruso, F.; Al-Salami, H. Release and swelling studies of an innovative antidiabetic-bile acid microencapsulated formulation, as a novel targeted therapy for diabetes treatment. J. Microencapsul. 2015, 32, 151–156. [Google Scholar] [CrossRef]
- Mooranian, A.; Negrulj, R.; Al-Sallami, H.S.; Fang, Z.; Mikov, M.; Golocorbin-Kon, S.; Fakhoury, M.; Watts, G.F.; Matthews, V.; Arfuso, F.; et al. Probucol Release from Novel Multicompartmental Microcapsules for the Oral Targeted Delivery in Type 2 Diabetes. AAPS PharmSciTech 2014, 16, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Salami, H.; Mooranian, A.; Negrulj, R.; Chen-Tan, N.; Al-Sallami, H.S.; Fang, Z.; Mukkur, T.; Mikov, M.; Goločorbin-Kon, S.; Watts, G.F.; et al. Microencapsulation as a novel delivery method for the potential antidiabetic drug, Probucol. Drug Des. Dev. Ther. 2014, 8, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Mooranian, A.; Negrulj, R.; Mathavan, S.; Martinez, J.; Sciarretta, J.; Chen-Tan, N.; Mukkur, T.; Mikov, M.; Lalic-Popovic, M.; Stojančević, M.; et al. Stability and Release Kinetics of an Advanced Gliclazide-Cholic Acid Formulation: The Use of Artificial-Cell Microencapsulation in Slow Release Targeted Oral Delivery of Antidiabetics. J. Pharm. Innov. 2014, 9, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alagpulinsa, D.A.; Cao, J.J.L.; Driscoll, R.K.; Sîrbulescu, R.F.; Penson, M.F.E.; Sremac, M.; Engquist, E.; Brauns, T.A.; Markmann, J.F.; Melton, D.A.; et al. Alginate-microencapsulation of human stem cell–derived β cells with CXCL 12 prolongs their survival and function in immunocompetent mice without systemic immunosuppression. Am. J. Transplant. 2019, 19, 1930–1940. [Google Scholar] [CrossRef] [PubMed]
- Mooranian, A.; Negrulj, R.; Chen-Tan, N.; Fakhoury, M.; Arfuso, F.; Jones, F.; Al-Salami, H. Advanced bile acid-based multi-compartmental microencapsulated pancreatic beta-cells integrating a polyelectrolyte-bile acid formulation, for diabetes treatment. Artif. Cells Nanomed. Biotechnol. 2016, 44, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Mooranian, A.; Negrulj, R.; Arfuso, F.; Al-Salami, H. Characterization of a novel bile acid-based delivery platform for microencapsulated pancreatic β-cells. Artif. Cells Nanomed. Biotechnol. 2014, 44, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, A.J.; O’Neal, D.N.; Nolan, C.J.; Januszewski, A.S. The Pathobiology of Diabetes Mellitus. In Pancreatic Islet Biology; Hardikar, A.A., Ed.; Springer International Publishing: Cham, Denmark, 2016; pp. 1–48. [Google Scholar] [CrossRef]
- Da Silva Xavier, G. The Cells of the Islets of Langerhans. J. Clin. Med. 2018, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.B.; De Leo, D.; Joseph, J.W.; McQuaid, T.S.; Ha, X.F.; Xu, F.; Tsushima, R.G.; Pennefather, P.S.; Salapatek, A.M.; Wheeler, M.B. Increased uncoupling protein-2 levels in beta-cells are associated with impaired glucose-stimulated insulin secretion: Mechanism of action. Diabetes 2001, 50, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Rorsman, P.; Ashcroft, F.M. Pancreatic β-Cell Electrical Activity and Insulin Secretion: Of Mice and Men. Physiol. Rev. 2018, 98, 117–214. [Google Scholar] [CrossRef]
- Hohmeier, H.E.; Newgard, C.B. Cell lines derived from pancreatic islets. Mol. Cell. Endocrinol. 2004, 228, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Skelin, M.; Rupnik, M.; Cencic, A. Pancreatic beta cell lines and their applications in diabetes mellitus research. ALTEX 2010, 27, 105–113. [Google Scholar] [CrossRef] [Green Version]
- McClenaghan, N.H.; Flatt, P.R. Engineering cultured insulin-secreting pancreatic B-cell lines. J. Mol. Med. 1999, 77, 235–243. [Google Scholar] [CrossRef]
- Ulloa-Aguirre, A.; Conn, P.M. Cellular Endocrinology in Health and Disease; Elsevier Science & Technology: San Diego, CA, USA, 2014. [Google Scholar]
- Green, A.D.; Vasu, S.; Flatt, P.R. Cellular models for beta-cell function and diabetes gene therapy. Acta Physiol. 2018, 222, e13012. [Google Scholar] [CrossRef]
- Miyazaki, S.; Tashiro, F.; Tsuchiya, T.; Sasaki, K.; Miyazaki, J.-i. Establishment of a long-term stable β-cell line and its application to analyze the effect of Gcg expression on insulin secretion. Sci. Rep. 2021, 11, 477. [Google Scholar] [CrossRef]
- Efrat, S. Regulation of insulin secretion: Insights from engineered beta-cell lines. Ann. N. Y. Acad. Sci. 2004, 1014, 88–96. [Google Scholar] [CrossRef]
- Cui, Y.X.; Shakesheff, K.M.; Adams, G. Encapsulation of RIN-m5F cells within Ba2+ cross-linked alginate beads affects proliferation and insulin secretion. J. Microencapsul. 2006, 23, 663–676. [Google Scholar] [CrossRef] [PubMed]
- McClenaghan, N.H. Physiological regulation of the pancreatic β-cell: Functional insights for understanding and therapy of diabetes. Exp. Physiol. 2007, 92, 481. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M.; McCluskey, J.T.; McClenaghan, N.H.; Flatt, P.R. Functional examination of microencapsulated bioengineered insulin-secreting beta-cells. Cell Biol. Int. 2001, 25, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, J.; Araki, K.; Yamato, E.; Ikegami, H.; Asano, T.; Shibasaki, Y.; Oka, Y.; Yamamura, K. Establishment of a pancreatic beta cell line that retains glucose-inducible insulin secretion: Special reference to expression of glucose transporter isoforms. Endocrinology 1990, 127, 126–132. [Google Scholar] [CrossRef]
- Ulrich, A.B.; Schmied, B.M.; Standop, J.; Schneider, M.B.; Pour, P.M. Pancreatic cell lines: A review. Pancreas. 2002, 24, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Han, J.; Wang, Q.; Guo, S.; Sun, H.; Zhang, X. Effects of sesamin on streptozotocin (STZ)-induced NIT-1 pancreatic beta-cell damage. Int. J. Mol. Sci. 2012, 13, 16961–16970. [Google Scholar] [CrossRef] [Green Version]
- Poitout, V.; Stout, L.E.; Armstrong, M.B.; Walseth, T.F.; Sorenson, R.L.; Robertson, R.P. Morphological and functional characterization of beta TC-6 cells-an insulin-secreting cell line derived from transgenic mice. Diabetes 1995, 44, 306–313. [Google Scholar] [CrossRef]
- Efrat, S.; Leiser, M.; Surana, M.; Tal, M.; Fusco-Demane, D.; Fleischer, N. Murine insulinoma cell line with normal glucose-regulated insulin secretion. Diabetes 1993, 42, 901–907. [Google Scholar] [CrossRef]
- Lin, Y.-J.; Wang, J.-P.; Chung, Y.-T.; Sun, Y.-L.; Chou, Y.-C. Viability and Functions of Alginate-microencapsulated Islets Isolated from Neonatal Pigs. Asian-Australas. J. Anim. Sci. 2007, 20, 795–801. [Google Scholar] [CrossRef]
- Suzuki, R.; Okada, N.; Miyamoto, H.; Yoshioka, T.; Sakamoto, K.; Oka, H.; Tsutsumi, Y.; Nakagawa, S.; Miyazaki, J.; Mayumi, T. Cyotomedical therapy for insulinopenic diabetes using microencapsulated pancreatic beta cell lines. Life Sci. 2002, 71, 1717–1729. [Google Scholar] [CrossRef]
- Bertolotti, A.; Borgogna, M.; Facoetti, A.; Marsich, E.; Nano, R. The effects of alginate encapsulation on NIT-1 insulinoma cells: Viability, growth and insulin secretion. In Vivo 2009, 23, 929–935. [Google Scholar]
- Carter, J.D.; Dula, S.B.; Corbin, K.L.; Wu, R.; Nunemaker, C.S. A practical guide to rodent islet isolation and assessment. Biol. Proced. Online 2009, 11, 3–31. [Google Scholar] [CrossRef] [Green Version]
- Marzorati, S.; Ramirez-Dominguez, M. Mouse Islet Isolation. Islets Langerhans 2015, 83–107. [Google Scholar] [CrossRef]
- Scharp, D.W.; Marchetti, P. Encapsulated islets for diabetes therapy: History, current progress, and critical issues requiring solution. Adv. Drug Deliv. Rev. 2014, 67–68, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Smink, A.M.; Faas, M.M.; de Vos, P. Toward Engineering a Novel Transplantation Site for Human Pancreatic Islets. Diabetes 2013, 62, 1357–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, J.L.; Williams, G.; Williams, L.J.; Tuch, B.E. Differentiation of transplanted microencapsulated fetal pancreatic cells. Transplantation 2007, 83, 1440–1448. [Google Scholar] [CrossRef]
- Si, Z.; Tuch, B.E.; Walsh, D.A. Development of human fetal pancreas after transplantation into SCID mice. Cells Tissues Organs 2001, 168, 147–157. [Google Scholar] [CrossRef]
- Scharfmann, R.; Pechberty, S.; Hazhouz, Y.; von Bulow, M.; Bricout-Neveu, E.; Grenier-Godard, M.; Guez, F.; Rachdi, L.; Lohmann, M.; Czernichow, P.; et al. Development of a conditionally immortalized human pancreatic beta cell line. J. Clin. Investig. 2014, 124, 2087–2098. [Google Scholar] [CrossRef] [Green Version]
- Ravassard, P.; Hazhouz, Y.; Pechberty, S.; Bricout-Neveu, E.; Armanet, M.; Czernichow, P.; Scharfmann, R. A genetically engineered human pancreatic β cell line exhibiting glucose-inducible insulin secretion. J. Clin. Investig. 2011, 121, 3589–3597. [Google Scholar] [CrossRef] [PubMed]
- Tsonkova, V.G.; Sand, F.W.; Wolf, X.A.; Grunnet, L.G.; Kirstine Ringgaard, A.; Ingvorsen, C.; Winkel, L.; Kalisz, M.; Dalgaard, K.; Bruun, C.; et al. The EndoC-βH1 cell line is a valid model of human beta cells and applicable for screenings to identify novel drug target candidates. Mol. Metab. 2018, 8, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Weir, G.C. Islet encapsulation: Advances and obstacles. Diabetologia 2013, 56, 1458–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.; Hallé, J.-P.; Poncelet, D.; Neufeld, R. Therapeutic cell encapsulation techniques and applications in diabetes. Adv. Drug Deliv. Rev. 2014, 67, 74–83. [Google Scholar] [CrossRef]
- Lin, C.C.; Anseth, K.S. Cell-cell communication mimicry with poly(ethylene glycol) hydrogels for enhancing beta-cell function. Proc. Natl. Acad. Sci. USA 2011, 108, 6380–6385. [Google Scholar] [CrossRef] [Green Version]
- Beck, J.; Angus, R.; Madsen, B.; Britt, D.; Vernon, B.; Nguyen, K.T. Islet encapsulation: Strategies to enhance islet cell functions. Tissue Eng. 2007, 13, 589–599. [Google Scholar] [CrossRef]
- Dorati, R.; Genta, I.; Modena, T.; Conti, B. Microencapsulation of a hydrophilic model molecule through vibration nozzle and emulsion phase inversion technologies. J. Microencapsul. 2013, 30, 559–570. [Google Scholar] [CrossRef]
- Patil, P.; Chavanke, D.; Wagh, M. A review on ionotropic gelation method: Novel approach for controlled gastroretentive gelispheres. Int. J. Pharm 2012, 4, 27–32. [Google Scholar]
- Jones, M.; Walker, D.; Ionescu, C.M.; Kovacevic, B.; Wagle, S.R.; Mooranian, A.; Brown, D.; Al-Salami, H. Microencapsulation of Coenzyme Q10 and bile acids using ionic gelation vibrational jet flow technology for oral delivery. Ther. Deliv. 2020, 11, 791–805. [Google Scholar] [CrossRef]
- Wagle, S.R.; Walker, D.; Kovacevic, B.; Gedawy, A.; Mikov, M.; Golocorbin-Kon, S.; Mooranian, A.; Al-Salami, H. Micro-Nano formulation of bile-gut delivery: Rheological, stability and cell survival, basal and maximum respiration studies. Sci. Rep. 2020, 10, 7715. [Google Scholar] [CrossRef] [PubMed]
- Wagle, S.R.; Kovacevic, B.; Walker, D.; Ionescu, C.M.; Shah, U.; Stojanovic, G.; Kojic, S.; Mooranian, A.; Al-Salami, H. Alginate-based drug oral targeting using bio-micro/nano encapsulation technologies. Expert Opin. Drug Deliv. 2020, 17, 1361–1376. [Google Scholar] [CrossRef]
- Mooranian, A.; Negrulj, R.; Mathavan, S.; Martinez, J.; Sciarretta, J.; Chen-Tan, N.; Mukkur, T.K.; Mikov, M.; Lalic-Popovic, M.; Stojancevic, M.; et al. An advanced microencapsulated system: A platform for optimized oral delivery of antidiabetic drug-bile acid formulations. Pharm. Dev. Technol. 2014, 20, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.; Meghwal, M.; Das, K. Microencapsulation: An overview on concepts, methods, properties and applications in foods. Food Front. 2021. [Google Scholar] [CrossRef]
- Tomei, A.A.; Manzoli, V.; Fraker, C.; Giraldo, J.; Velluto, D.; Najjar, M.; Pileggi, A.; Molano, R.; Ricordi, C.; Stabler, C.L.; et al. Device design and materials optimization of conformal coating for islets of Langerhans. Proc. Natl. Acad. Sci. USA 2014, 111, 10514–10519. [Google Scholar] [CrossRef] [Green Version]
- Villa, C.; Manzoli, V.; Abreu, M.M.; Verheyen, C.A.; Seskin, M.; Najjar, M.; Molano, R.D.; Torrente, Y.; Ricordi, C.; Tomei, A.A. Effects of Composition of Alginate-Polyethylene Glycol Microcapsules and Transplant Site on Encapsulated Islet Graft Outcomes in Mice. Transplantation 2017, 101, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Stock, A.A.; Manzoli, V.; De Toni, T.; Abreu, M.M.; Poh, Y.C.; Ye, L.; Roose, A.; Pagliuca, F.W.; Thanos, C.; Ricordi, C.; et al. Conformal Coating of Stem Cell-Derived Islets for β Cell Replacement in Type 1 Diabetes. Stem Cell Rep. 2020, 14, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Ren, Y.; Wang, W.; Hao, H.; Tang, M.; Zhang, Z.; Yang, J.; Zheng, Y.; Shi, X. Transglutaminase-Catalyzed Encapsulation of Individual Mammalian Cells with Biocompatible and Cytoprotective Gelatin Nanoshells. ACS Biomater. Sci. Eng. 2020, 6, 2336–2345. [Google Scholar] [CrossRef]
- Lim, G.J.; Zare, S.; Van Dyke, M.; Atala, A. Cell Microencapsulation. In Therapeutic Applications of Cell Microencapsulation; Pedraz, J.L., Orive, G., Eds.; Springer: New York, NY, USA, 2010; pp. 126–136. [Google Scholar] [CrossRef]
- Farina, M.; Alexander, J.F.; Thekkedath, U.; Ferrari, M.; Grattoni, A. Cell encapsulation: Overcoming barriers in cell transplantation in diabetes and beyond. Adv. Drug Deliv. Rev. 2019, 139, 92–115. [Google Scholar] [CrossRef]
- Desai, T.; Shea, L.D. Advances in islet encapsulation technologies. Nat. Rev. Drug Discov. 2017, 16, 338–350. [Google Scholar] [CrossRef]
- Gasperini, L.; Mano, J.F.; Reis, R.L. Natural polymers for the microencapsulation of cells. J. R. Soc. Interface 2014, 11, 20140817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.C.; Chang, T.M.S. Artificial cell microencapsulated stem cells in regenerative medicine, tissue engineering and cell therapy. Adv. Exp. Med. Biol. 2010, 670, 68–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.M.S. Therapeutic applications of polymeric artificial cells. Nat. Rev. Drug Discov. 2005, 4, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Ashimova, A.; Yegorov, S.; Negmetzhanov, B.; Hortelano, G. Cell Encapsulation Within Alginate Microcapsules: Immunological Challenges and Outlook. Front. Bioeng. Biotechnol. 2019, 7, 380. [Google Scholar] [CrossRef] [Green Version]
- Olabisi, R.M. Cell microencapsulation with synthetic polymers. J. Biomed. Mater. Res. Part A 2015, 103, 846–859. [Google Scholar] [CrossRef] [Green Version]
- Ryan, A.J.; O’Neill, H.S.; Duffy, G.P.; O’Brien, F.J. Advances in polymeric islet cell encapsulation technologies to limit the foreign body response and provide immunoisolation. Curr. Opin. Pharmacol. 2017, 36, 66–71. [Google Scholar] [CrossRef]
- O’Shea, G.M.; Sun, A.M. Encapsulation of rat islets of Langerhans prolongs xenograft survival in diabetic mice. Diabetes 1986, 35, 943–946. [Google Scholar] [CrossRef]
- Soon-Shiong, P. Encapsulated islet cell therapy for the treatment of diabetes: Intraperitoneal injection of islets. J. Control. Release 1996, 39, 399–409. [Google Scholar] [CrossRef]
- Soon-Shiong, P.; Feldman, E.; Nelson, R.; Komtebedde, J.; Smidsrod, O.; Skjak-Braek, G.; Espevik, T.; Heintz, R.; Lee, M. Successful Reversal of spontaneous diabetes in dogs by intraperitoneal Microencapsulated islets. Transplantation 1992, 54, 769–774. [Google Scholar] [CrossRef]
- Bochenek, M.A.; Veiseh, O.; Vegas, A.J.; McGarrigle, J.J.; Qi, M.; Marchese, E.; Omami, M.; Doloff, J.C.; Mendoza-Elias, J.; Nourmohammadzadeh, M.; et al. Alginate encapsulation as long-term immune protection of allogeneic pancreatic islet cells transplanted into the omental bursa of macaques. Nat. Biomed. Eng. 2018, 2, 810–821. [Google Scholar] [CrossRef]
- Whelehan, M.; Marison, I.W. Microencapsulation using vibrating technology. J. Microencapsul. 2011, 28, 669–688. [Google Scholar] [CrossRef]
- Mooranian, A.; Zamani, N.; Kovacevic, B.; Ionescu, C.M.; Luna, G.; Mikov, M.; Goločorbin-Kon, S.; Stojanovic, G.; Kojic, S.; Al-Salami, H. Pharmacological effects of secondary bile acid microparticles in diabetic murine model. Curr. Diabetes Rev. 2022, 18. [Google Scholar] [CrossRef]
- Mathavan, S.; Ionescu, C.M.; Kovacevic, B.; Mikov, M.; Golocorbin-Kon, S.; Mooranian, A.; Dass, C.R.; Al-Salami, H. Histological effects of pharmacologically active human bile acid nano/micro-particles in Type-1 diabetes. Ther. Deliv. 2020, 11, 157–171. [Google Scholar] [CrossRef]
- Mooranian, A.; Zamani, N.; Mikov, M.; Goločorbin-Kon, S.; Stojanovic, G.; Arfuso, F.; Al-Salami, H. Stability and biological testing of taurine-conjugated bile acid antioxidant microcapsules for diabetes treatment. Ther. Deliv. 2019, 10, 99–106. [Google Scholar] [CrossRef]
- Mamo, J.C.L.; Lam, V.; Brook, E.; Mooranian, A.; Al-Salami, H.; Fimognari, N.; Nesbit, M.; Takechi, R. Probucol prevents blood–brain barrier dysfunction and cognitive decline in mice maintained on pro-diabetic diet. Diabetes Vasc. Dis. Res. 2019, 16, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooranian, A.; Negrulj, R.; Chen-Tan, N.; Al-Sallami, H.S.; Fang, Z.; Mukkur, T.; Mikov, M.; Golocorbin-Kon, S.; Fakhoury, M.; Arfuso, F.; et al. Novel artificial cell microencapsulation of a complex gliclazide-deoxycholic bile acid formulation: A characterization study. Drug Des. Dev. Ther. 2014, 8, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Loh, Q.L.; Wong, Y.Y.; Choong, C. Combinatorial effect of different alginate compositions, polycations, and gelling ions on microcapsule properties. Colloid Polym. Sci. 2012, 290, 619–629. [Google Scholar] [CrossRef]
- Chan, G.; Mooney, D.J. Ca2+ released from calcium alginate gels can promote inflammatory responses in vitro and in vivo. Acta Biomater. 2013, 9, 9281–9291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morch, Y.A.; Qi, M.; Gundersen, P.O.; Formo, K.; Lacik, I.; Skjak-Braek, G.; Oberholzer, J.; Strand, B.L. Binding and leakage of barium in alginate microbeads. J. Biomed. Mater. Res. Part A 2012, 100, 2939–2947. [Google Scholar] [CrossRef] [Green Version]
- Vaithilingam, V.; Kollarikova, G.; Qi, M.; Lacik, I.; Oberholzer, J.; Guillemin, G.J.; Tuch, B.E. Effect of prolonged gelling time on the intrinsic properties of barium alginate microcapsules and its biocompatibility. J. Microencapsul. 2011, 28, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Hecht, H.; Srebnik, S. Structural Characterization of Sodium Alginate and Calcium Alginate. Biomacromolecules 2016, 17, 2160–2167. [Google Scholar] [CrossRef] [PubMed]
- Niaz, T.; Nasir, H.; Shabbir, S.; Rehman, A.; Imran, M. Polyionic hybrid nano-engineered systems comprising alginate and chitosan for antihypertensive therapeutics. Int. J. Biol. Macromol. 2016, 91, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.; Zarate, J.; Orive, G.; Hernández, R.M.; Pedraz, J.L. Biomaterials in Cell Microencapsulation. In Therapeutic Applications of Cell Microencapsulation; Pedraz, J.L., Orive, G., Eds.; Springer: New York, NY, USA, 2010; pp. 5–21. [Google Scholar] [CrossRef]
- Chávarri, M.; Marañón, I.; Villarán, M.C. Encapsulation Technology to Protect Probiotic Bacteria; IntechOpen: London, UK, 2012. [Google Scholar]
- Serp, D.; Cantana, E.; Heinzen, C.; Von Stockar, U.; Marison, I.W. Characterization of an encapsulation device for the production of monodisperse alginate beads for cell immobilization. Biotechnol. Bioeng. 2000, 70, 41–53. [Google Scholar] [CrossRef]
- Stark, D.; Kornmann, H.; Munch, T.; Sonnleitner, B.; Marison, I.W.; von Stockar, U. Novel type of in situ extraction: Use of solvent containing microcapsules for the bioconversion of 2-phenylethanol from L-phenylalanine by Saccharomyces cerevisiae. Biotechnol. Bioeng. 2003, 83, 376–385. [Google Scholar] [CrossRef]
- Chan, E.S.; Lee, B.B.; Ravindra, P.; Poncelet, D. Prediction models for shape and size of ca-alginate macrobeads produced through extrusion-dripping method. J. Colloid Interface Sci. 2009, 338, 63–72. [Google Scholar] [CrossRef]
- de Vos, P.; Lazarjani, H.A.; Poncelet, D.; Faas, M.M. Polymers in cell encapsulation from an enveloped cell perspective. Adv. Drug Deliv. Rev. 2014, 67–68, 15–34. [Google Scholar] [CrossRef]
- Maxwell, K.G.; Millman, J.R. Applications of iPSC-derived beta cells from patients with diabetes. Cell Rep. Med. 2021, 2, 100238. [Google Scholar] [CrossRef]
- Shahjalal, H.M.; Abdal Dayem, A.; Lim, K.M.; Jeon, T.-I.; Cho, S.-G. Generation of pancreatic β cells for treatment of diabetes: Advances and challenges. Stem Cell Res. Ther. 2018, 9, 355. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Du, K.; Zou, C. Current progress in stem cell therapy for type 1 diabetes mellitus. Stem Cell Res. Ther. 2020, 11, 275. [Google Scholar] [CrossRef]
- Legøy, T.A.; Vethe, H.; Abadpour, S.; Strand, B.L.; Scholz, H.; Paulo, J.A.; Ræder, H.; Ghila, L.; Chera, S. Encapsulation boosts islet-cell signature in differentiating human induced pluripotent stem cells via integrin signalling. Sci. Rep. 2020, 10, 414. [Google Scholar] [CrossRef] [Green Version]
- Legøy, T.A.; Mathisen, A.F.; Salim, Z.; Vethe, H.; Bjørlykke, Y.; Abadpour, S.; Paulo, J.A.; Scholz, H.; Ræder, H.; Ghila, L.; et al. In vivo Environment Swiftly Restricts Human Pancreatic Progenitors Toward Mono-Hormonal Identity via a HNF1A/HNF4A Mechanism. Front. Cell Dev. Biol. 2020, 8, 109. [Google Scholar] [CrossRef]
- Carlsson, P.-O.; Espes, D.; Sedigh, A.; Rotem, A.; Zimerman, B.; Grinberg, H.; Goldman, T.; Barkai, U.; Avni, Y.; Westermark, G.T.; et al. Transplantation of macroencapsulated human islets within the bioartificial pancreas βAir to patients with type 1 diabetes mellitus. Am. J. Transplant. 2018, 18, 1735–1744. [Google Scholar] [CrossRef]
- Ilic, D.; Liovic, M. Industry updates from the field of stem cell research and regenerative medicine in January 2021. Regen. Med. 2021, 16, 423–429. [Google Scholar] [CrossRef]
- Ilic, D.; Liovic, M. Industry updates from the field of stem cell research and regenerative medicine in February 2021. Regen. Med. 2021, 16, 517–523. [Google Scholar] [CrossRef]
- Shapiro, A.; Thompson, D.; Donner, T.W.; Bellin, M.D.; Hsueh, W.; Pettus, J.H.; Wilensky, J.S.; Daniels, M.; Wang, R.M.; Kroon, E.J. Insulin Expression and Glucose-Responsive Circulating C-peptide in Type 1 Diabetes Patients Implanted Subcutaneously with Pluripotent Stem Cell-Derived Pancreatic Endoderm Cells in a Macro-Device. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3501034 (accessed on 15 February 2021).
- Lacík, I. Polymer chemistry in diabetes treatment by encapsulated islets of Langerhans: Review to 2006. Aust. J. Chem. 2006, 59, 508–524. [Google Scholar] [CrossRef]
- Calafiore, R.; Basta, G. Clinical application of microencapsulated islets: Actual prospectives on progress and challenges. Adv. Drug Deliv. Rev. 2013, 67–68, 84–92. [Google Scholar] [CrossRef]
- Orive, G.; Maria Hernández, R.; Rodríguez Gascón, A.; Calafiore, R.; Swi Chang, T.M.; Vos, P.d.; Hortelano, G.; Hunkeler, D.; Lacík, I.; Luis Pedraz, J. History, challenges and perspectives of cell microencapsulation. Trends Biotechnol. 2004, 22, 87–92. [Google Scholar] [CrossRef] [Green Version]
- De Vos, P.; Hamel, A.; Tatarkiewicz, K. Considerations for successful transplantation of encapsulated pancreatic islets. Diabetologia 2002, 45, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.R.; Khattak, S.F.; Roberts, S.C. Polyelectrolytes for cell encapsulation. Curr. Opin. Colloid Interface Sci. 2005, 10, 45–51. [Google Scholar] [CrossRef]
- Wagle, S.R.; Kovacevic, B.; Ionescu, C.M.; Walker, D.; Jones, M.; Carey, L.; Takechi, R.; Mikov, M.; Mooranian, A.; Al-Salami, H. Pharmacological and biological study of microencapsulated probucol-secondary bile acid in a diseased mouse model. Pharmaceutics 2021, 13, 1223. [Google Scholar] [CrossRef]
- Mooranian, A.; Jones, M.; Ionescu, C.; Walker, D.; Wagle, S.; Kovacevic, B.; Chester, J.; Foster, T.; John-ston, E.; Mikov, M.; et al. Advancements in Assessments of Bio-Tissue Engineering and Viable Cell De-livery Matrices Using Bile Acid-Based Pharmacological Biotechnologies. Nanomaterials 2021, 11, 1861. [Google Scholar] [CrossRef]
- Mooranian, A.; Ionescu, C.M.; Wagle, S.R.; Kovacevic, B.; Walker, D.; Jones, M.; Chester, J.; Johnston, E.; Danic, M.; Mikov, M.; et al. Chenodeoxycholic Acid Pharmacology in Biotechnology and Transplantable Pharmaceutical Applications for Tissue Delivery: An Acute Preclinical Study. Cells 2021, 10, 2437. [Google Scholar] [CrossRef]
- Mooranian, A.; Ionescu, C.M.; Wagle, S.R.; Kovacevic, B.; Walker, D.; Jones, M.; Chester, J.; Foster, T.; Johnston, E.; Mikov, M.; et al. Probucol pharmacological and bio-nanotechnological effects on surgically transplanted graft due to powerful anti-inflammatory, anti-fibrotic and potential bile acid modulatory actions. Pharmaceutics 2021, 13, 1304. [Google Scholar] [CrossRef] [PubMed]
- Mooranian, A.; Ionescu, C.M.; Wagle, S.R.; Kovacevic, B.; Walker, D.; Jones, M.; Chester, J.; Foster, T.; Johnston, E.; Kojic, S.; et al. Polyelectrolytes formulated with primary unconjugated bile acid optimised pharmacology of bio-engineered implant. Pharmaceutics 2021, 13, 1713. [Google Scholar] [CrossRef]
- Mooranian, A.; Foster, T.; Ionescu, C.; Walker, D.; Jones, M.; Wagle, S.; Kovacevic, B.; Chester, J.; John-ston, E.; Wong, E.; et al. Enhanced Bilosomal Properties Resulted in Optimum Pharmacological Effects by Increased Acidification Pathways. Pharmaceutics 2021, 13, 1184. [Google Scholar] [CrossRef] [PubMed]
- Mooranian, A.; Foster, T.; Ionescu, C.M.; Carey, L.; Walker, D.; Jones, M.; Wagle, S.R.; Kovacevic, B.; Chester, J.; Johnstone, E.; et al. The Effects of Primary Unconjugated Bile Acids on Nanoencapsulated Pharmaceutical Formulation of Hydrophilic Drugs: Pharmacological Implications. Drug Des. Dev. Ther. 2021, 15, 4423–4434. [Google Scholar] [CrossRef]
- Mooranian, A.; Carey, L.; Ionescu, C.M.; Walker, D.; Jones, M.; Wagle, S.R.; Kovacevic, B.; Foster, T.; Chester, J.; Johnston, E.; et al. The Effects of Accelerated Temperature-Controlled Stability Systems on the Release Profile of Primary Bile Acid-Based Delivery Microcapsules. Pharmaceutics 2021, 13, 1667. [Google Scholar] [CrossRef]
- Foster, T.; Ionescu, C.; Walker, D.; Jones, M.; Wagle, S.; Kovacevic, B.; Brown, D.; Mikov, M.; Mooranian, A.; Al-Salami, H. Chemotherapy-induced hearing loss: The applications of bio-nanotechnologies and bile acid-based delivery matrices. Ther. Deliv. 2021, 12, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Chester, J.; Johnston, E.; Walker, D.; Jones, M.; Ionescu, C.; Wagle, S.; Kovacevic, B.; Brown, D.; Mikov, M.; Mooranian, A.; et al. A Review on Recent Advancement on Age-Related Hearing Loss: The Applica-tions of Nanotechnology, Drug Pharmacology, and Biotechnology. Pharmaceutics 2021, 13, 1041. [Google Scholar] [CrossRef]
- Carey, L.; Walker, D.; Jones, M.; Ionescu, C.; Wagle, S.; Kovacevic, B.; Brown, D.; Mikov, M.; Mooranian, A.; Al-Salami, H. Bile acid-permeation enhancement for inner ear cochlear drug–pharmacological uptake: Bio-nanotechnologies in chemotherapy-induced hearing loss. Ther. Deliv. 2021. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mooranian, A.; Jones, M.; Ionescu, C.M.; Walker, D.; Wagle, S.R.; Kovacevic, B.; Chester, J.; Foster, T.; Johnston, E.; Kuthubutheen, J.; et al. Artificial Cell Encapsulation for Biomaterials and Tissue Bio-Nanoengineering: History, Achievements, Limitations, and Future Work for Potential Clinical Applications and Transplantation. J. Funct. Biomater. 2021, 12, 68. https://doi.org/10.3390/jfb12040068
Mooranian A, Jones M, Ionescu CM, Walker D, Wagle SR, Kovacevic B, Chester J, Foster T, Johnston E, Kuthubutheen J, et al. Artificial Cell Encapsulation for Biomaterials and Tissue Bio-Nanoengineering: History, Achievements, Limitations, and Future Work for Potential Clinical Applications and Transplantation. Journal of Functional Biomaterials. 2021; 12(4):68. https://doi.org/10.3390/jfb12040068
Chicago/Turabian StyleMooranian, Armin, Melissa Jones, Corina Mihaela Ionescu, Daniel Walker, Susbin Raj Wagle, Bozica Kovacevic, Jacqueline Chester, Thomas Foster, Edan Johnston, Jafri Kuthubutheen, and et al. 2021. "Artificial Cell Encapsulation for Biomaterials and Tissue Bio-Nanoengineering: History, Achievements, Limitations, and Future Work for Potential Clinical Applications and Transplantation" Journal of Functional Biomaterials 12, no. 4: 68. https://doi.org/10.3390/jfb12040068