
Glutathione S-Transferases in Marine Copepods




1
Marine Biotechnology Department, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
2
Integrative Marine Ecology Department, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
*
Author to whom correspondence should be addressed.
†
These authors equally contributed to the work.
J. Mar. Sci. Eng. 2021, 9(9), 1025; https://doi.org/10.3390/jmse9091025
Submission received: 2 August 2021
/
Revised: 14 September 2021
/
Accepted: 15 September 2021
/
Published: 18 September 2021
(This article belongs to the Special Issue Marine Omics & Biotechnology)
Abstract
:The glutathione S-transferase (GST) is a complex family of phase II detoxification enzymes, known for their ability to catalyze the conjugation of the reduced form of glutathione (GSH) to a wide variety of endogenous and exogenous electrophilic compounds for detoxification purposes. In marine environments, copepods are constantly exposed to multiple exogenous stressors, thus their capability of detoxification is key for survival. Full identification of the GST family in copepods has been limited only to few species. As for insects, the GST family includes a wide range of genes that, based on their cellular localization, can be divided in three classes: cytosolic, microsomal, and mitochondrial. The role of GSTs might have class-specific features, thus understanding the nature of the GST family has become crucial. This paper covers information of the GST activity in marine copepods based on studies investigating gene expression, protein content, and enzymatic activity. Using published literature and mining new publicly available transcriptomes, we characterized the multiplicity of the GST family in copepods from different orders and families, highlighting the possible role of these genes as biomarker for ocean health status monitoring.
1. Introduction
Marine organisms are continuously exposed to stressful conditions, including both biotic and abiotic stressors, which may alter the homeostasis of an organism leading, in extreme conditions, to its death. In order to adapt and survive to stressful conditions, marine organisms have evolved defense strategies, which include the activation of detoxification enzymes [1]. Glutathione S-transferases (GSTs) are a family of phase II detoxification enzymes, known for their ability to catalyze the conjugation of the reduced form of glutathione (GSH) to a wide variety of endogenous and exogenous electrophilic compounds for detoxification purposes [2]. Glutathione, a tripeptide formed by glutamate, cysteine, and glycine, can be a scavenger molecule on its own, but also a substrate in enzymatic reactions [3]. In its reduced form, glutathione (GSH) has a free thiol group on the cysteine residue. Two thiol groups react with each other, losing two electrons, giving rise to the oxidized glutathione form (GSSG) (reaction catalyzed by glutathione peroxidase), which can be again reduced to GSH by the enzyme glutathione reductase.
GSTs transfer the GSH to the product obtained from phase I reactions (reactive/polar groups are added to a xenobiotic by enzymatic activities, mainly by the cytochrome P450 enzymes) in order to facilitate its elimination from the body, generally by the activity of the ATP-binding cassette (ABC) transporter proteins [4]. GSTs have been found in both prokaryotes and eukaryotes [3]. The family is highly complex with multiple genes, which, depending on their cellular localization, belong to three classes: cytosolic, microsomal, and mitochondrial (Figure 1). The cytosolic class consists of six subclasses (delta/epsilon, omega, sigma, theta, mu, zeta) that are mainly involved in detoxification activities [5]. In insects, members of the delta/epsilon subclass are implicated in resistance to pesticides, while the omega, theta, and zeta subclasses appear to be involved in other cellular processes, including protection against oxidative stress [5]. The microsomal GSTs (membrane-associated proteins in eicosanoid and glutathione metabolism, MAPEG), which also include several subclasses (mGST1-mGST8), are mainly involved in the eicosanoid and glutathione metabolism, and are localized to the mitochondrion and endoplasmic reticulum (ER). The mitochondrial class, also known as kappa GST, mostly represented by a single gene, shares high protein domain similarity with cytosolic GSTs, thus it is likely to have the same substrate specificity.
Among marine organisms, copepods play a central role in food webs and biogeochemical cycles [6]. These zooplankters are naturally exposed to a wide array of biological and anthropogenic stressors, with negative impact on secondary production and population recruitment [7,8]. Due to their key role in detoxification, GSTs are considered important biomarkers of xenobiotic-induced oxidative stress in ecotoxicological studies. Understanding their species-specific differences is key to obtaining insights into the defense mechanisms that copepods potentially employ to respond to various environmental stressors. To date, the diversity of the GST superfamily has been characterized in some marine copepods, including two calanoids, C. finmarchicus and Eurytemora affinis [5,9]; two harpacticoids, T. japonicus and T. kingsejongensis [9,10]; and a single cyclopoid, P. nana [10]. In recent years, new genomic and transcriptomic resources have become available also for many other copepods for which the GST activity has been previously investigated (see below, Table 1). The aim of this study was to outline the most recent studies on the glutathione S-transferase activity in copepods in response to abiotic and biotic factors. Furthermore, we mined, in silico, new publicly available copepod transcriptomes for which GSTs have been used as biomarkers of stress and compared the GST gene diversity with the one previously described in literature.
Glutathione S-Transferase Activity in Response to Environmental Stressors
In order to give a comprehensive overview of GSTs’ functional responses in copepods, studies showing gene expression changes by reverse transcription quantitative PCR (RT-qPCR) and RNA sequencing (RNA-seq), protein content variations, and enzymatic activities have been reported in Table 1. These studies investigated the GST responses after exposure to aquatic pollutants (e.g., heavy metals, polycyclic aromatic hydrocarbons, endocrine disrupting chemicals, nanoparticles, microplastics), algal toxins, or altered physical–chemical environmental parameters (temperature, pH).
The harpacticoid copepod Tigriopus japonicus has been often used for ecotoxicological studies. The GST gene expression and enzymatic activity in this copepod have been reported to increase after exposure to a water-accommodated fraction of crude oil [11,12]; endocrine disrupting chemicals [13], hydrogen peroxide [14], heavy metals (e.g., manganese [14], copper [14,15], nickel [16], cadmium [15,17], silver, and arsenic [15]), the brominated flame retardants (e.g., hexabromocyclododecanes) [18], low pH (pH 7.5 and 7.0; [19]), β-Naphtoflavone [20], the antimicrobial agent triclosan [21], and the widely used insecticide cypermethrin [22]. The cytosolic GST belonging to the sigma subclass, has been reported as the most up-regulated after exposure to both hydrogen peroxide and trace metals (i.e., silver, arsenic, cadmium, and copper). In contrast, a decrease in the GST expression was observed in T. japonicus exposed to triphenyltin [23], to the multi-walled carbon nano-tubes (MWCNTs), and to the nano-particles widely used in various industrial fields [24]. Similarly, the copepod Tigriopus californicus exposed to pro-oxidant conditions, i.e., hydrogen peroxide and paraquat, showed differential GST expression. In particular, both male and female individuals were exposed to hydrogen peroxide and paraquat for 3 and 6 h, and GSTs were found highly expressed in both sexes under both stress treatments at 6 h [25]. On the contrary, after 3 h of hydrogen peroxide exposure, two transcripts coding for GST were up-regulated and three were down-regulated in males, while four GSTs were up-regulated in males after paraquat exposure for 3 h. After 6 h, three GSTs were up-regulated in females exposed to both hydrogen peroxide and paraquat. In another transcriptomic study, T. californicus males and females were also exposed to hydrogen peroxide and a decreased antioxidant condition (reduced carotenoid due to a yeast diet) and results showed an increase in GST expression only in males [26].
Another copepod used in molecular ecotoxicology is the euryhaline estuarine species Eurytemora affinis. Cailleaud et al. [27] evaluated the GST enzymatic activity in E. affinis exposed to hydrophobic organic contaminants (HOC) of the Seine Estuary, the largest megatidal estuary along the English Channel. GST levels significantly increased where HOC were observed at the highest concentrations in the water column.
Significant increases in the expression of GSTs have also been reported in two dominant calanoid species from the Arctic and the boreal regions, Calanus glacialis and C. finmarchicus, exposed to mercury [28] and water-accommodated fractions (WAF) of oil [11]. In both studies, time-dependent and concentration-dependent responses were observed in the two species, however the time of response and its magnitude was different. When exposed to mercury, C. glacialis showed a quicker and higher GST expression (mRNA) compared with C. finmarchicus; conversely, exposure to WAF induced in C. finmarchicus a higher GST expression level compared to this congener [11]. In C. finmarchicus, significant expression of GST has also been reported in response to naphthalene [29] and dispersed oil [30].
Ensibi et al. [31] studied the effects of cadmium chloride (at 0, 0.2, and 0.4 µg/L for 24, 48, and 72 h exposure) on Centropages ponticus, one of the key species in the Mediterranean Sea. They found that GST was significantly influenced with a time- and concentration-dependent increase in GST activity, except for the 72 h exposure, for which the variation in activity was not significant. In a recent study, a subclass-specific expression was reported for two cytosolic GSTs (theta and mu) in the calanoid Acartia tonsa exposed to a polycyclic aromatic hydrocarbon (PAH) 1,2-dimethylnaphthalene [32]. The expression of the two GSTs varied depending on the concentrations (0.574, 5.736, and 57.358 μg/L) and the time of exposure (24 h, 48 h, 72 h, and 96 h). While GST-theta had its highest expression at 5.736 μg/L for 96 h, GST-mu was regulated already after 72 h at 0.574 μg/L and 5.736 μg/L. Similarly, the same stressor (PAH 1,2-dimethylnaphthalene) induced a significant increase in expression of two cytosolic GSTs (theta, delta/epsilon) in the copepod Pseudodiaptomus poplesia [33].
In recent years, plastic pollution has become a concern for the marine environment [34]; to date, few studies have focused on clarifying the effects that microplastics may have on copepods. Jeong et al. [35] exposed for 24 h the copepod Paracyclopina nana to polystyrene nano- and microbeads (20 µg/mL) with different sizes (0.05, 0.5, 6 µm); the authors investigated the effects on the copepod growth rate, fecundity, and GST activity. An increase in the GST activity was observed upon ingestion and egestion of both particles and specifically after exposure to 0.05 µm polystyrene nanobead, and to 0.5 and 6 µm microbeads. In the same copepod, the GST activity as an indicator of the antioxidant system was investigated when the copepod was exposed to methylmercury (MeHg) at various concentrations (1 ng/L, 10 ng/L, 100 ng/L, 500 ng/L, and 1,000 ng/L) [36]. While data showed an increase in GST activity after 24 h exposure to MeHg at 1–100 ng/L, no significant variations in the expression levels of the analyzed GST genes was found. GST activity was reported to increase in P. nana exposed to cypermethrin, a widely used insecticide in agriculture [22].
Various studies have also focused on the detoxification ability of the copepod sea lice, a salmon parasite responsible for severe economic losses in the aquaculture production worldwide. The effect of the delousing drug deltamethrin (AlphaMaxTM) was investigated in Caligus rogercressereyi by using a transcriptomic approach [37]. Between the differentially expressed genes, the authors found the down-regulation of the glutathione S-transferase DHAR1. Recently, Dalvin et al. [38] investigated a microsomal glutathione S-transferase 1 (LsMFST1L) in the salmon louse Lepeophtheirus salmonis by sampling various sea lice stages, from egg to adult lice in pentaplicate, and found a constitutive transcript level throughout the studied development stages, with the highest levels in ovaries or gut.
GST were also found to be significantly regulated when copepods were fed on toxic microalgae. In C. finmarchicus feeding on the saxitoxin-producing dinoflagellate Alexandrium fundyense, differential expression was reported for four GSTs belonging to different subclasses [39]. Increases in the GST expression levels were also reported in T. japonicus feeding on the dinoflagellate Gymnodinium catenatum [40] and in C. helgolandicus feeding on the toxic diatom Skeletonema marinoi in either laboratory or field conditions [41,42,43]. Down-regulation of GST was also found when C. helgolandicus was fed with the non-brevetoxin-producing dinoflagellate Karenia brevis [44], while GST did not show significant expression variation when C. sinicus was fed on S. marinoi [45]. Chen et al. [46] studied the effects of the ingestion of two diatoms, Chaetoceros muelleri and Nitzschia closterium f. minutissima, by three different copepod species, i.e., T. japonicus, and the pelagic calanoids Acartia pacifica and Pseudodiaptomus annandalei. In particular, they used the algae for rearing nauplii to adults and evaluated survival rates to adults and activities of the defensive enzymes, including GST, of the adult copepods. For T. japonicus, neither algal species induced significant GST activity. GST data for A. pacifica fed N. closterium f. minutissima were not available due to copepod mortality. GST activity of the copepod A. pacifica fed on C. muelleri induced a significant concentration-dependent increase in GST activity compared to copepods fed on Isochrysis galbana, used as control algae. Finally, GST activities increased in the copepod P. annandalei fed on both C. muelleri and N. closterium f. minutissima compared to the copepod fed on I. galbana, but this increase was not concentration-dependent.
Rises in pH and temperature are predicted by global climate change scenarios. In particular, current levels of atmospheric CO2 are expected to more than double by 2100 (Intergovernmental Panel on Climate Change IPCC 2007). Zhang et al. [47] tested and analyzed the effects of acidified seawater with elevated concentration of CO2 (i.e., 0.08, 0.20, 0.50, and 1.00%) on the planktonic copepod Centropages tenuiremis, but they did not find significant differences between the control and the treatment groups. Glippa et al. [48] studied temporal variations in water column physical–chemical characteristics, oxidative stress levels, and antioxidant responses of Acartia sp. (the samples were A. bifilosa and A. tonsa) copepods. Samples were collected during a two-week field survey (August 2015) in the western Gulf of Finland, Baltic Sea, where the expected pH and temperature changes are larger than values predicted by climate change scenarios predicted for the next century (up to 0.5 units in pH and 5 °C in temperature). GST activity was higher in week II compared to week I, and, in particular, it was higher at the surface compared to the other depths for both weeks. Altogether, the results showed that copepods were fairly tolerant to the weekly variations and suggest that this can be related to an efficient antioxidant defense system. Another study [49], performed during the 2016 West Coast Ocean Acidification cruise in the California Current System, also focused on understanding the effects of ocean temperature and pH variations on the oxidative stress defense system of the copepod Calanus pacificus, and compared its response to the one of the pelagic pteropod mollusk Limacina helicina. Copepods showed higher GST activities and total glutathione content compared to pteropods. Data suggested a greater copepod defense capacity and higher adaptive potential, but a risk for pteropods in a high CO2 global scenario. Souza et al. [43] also tested the effects of short-duration exposure of ultraviolet radiation on the freshwater calanoid copepod Eudiaptomus gracilis and found a rapid increase in GST activity.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of GST studies in marine copepods after exposure to stressful conditions. Copepod species, abiotic/biotic stress, copepod response, and approach used are listed for each study. GST families are specified when available. For approaches: (a) RT-qPCR, (b) RNA-Seq, (c) protein content, and (d) enzymatic activity.
Table 1.
Summary of GST studies in marine copepods after exposure to stressful conditions. Copepod species, abiotic/biotic stress, copepod response, and approach used are listed for each study. GST families are specified when available. For approaches: (a) RT-qPCR, (b) RNA-Seq, (c) protein content, and (d) enzymatic activity.
| Species | GST | Stress | Response | Approach | Ref |
|---|---|---|---|---|---|
| Tigriopus japonicus | GST | Water-accommodated fraction (WAF) of crude oil | Increase | a | [11] |
| GST | WAF of crude oil | Increase | d | [12] | |
| GST | Endocrine disruptor chemicals (i.e., 4,4′-octylphenol and polychlorinated biphenyl) | Increase for 4,4′-octylphenol and decrease for polychlorinated biphenyl | a, c | [13] | |
| GST | Heavy metals (e.g., manganese, copper, nickel, cadmium, silver, and arsenic) | Increase | a, c, d | [10,14,15,16,17,50] | |
| GST | Brominated flame retardants (e.g., hexabromocyclododecanes) | Increase | a | [18] | |
| GST sigma, GST omega, GST mu, GST delta/epsilon | β-Naphtoflavone | Increase | a | [20] | |
| GST theta | Triclosan | Increase | a, d | [21] | |
| GST | Hydrogen peroxide | Increase | a, d | [14] | |
| Ten GSTs | Hydrogen peroxide and trace metals (i.e., silver, arsenic, cadmium, and copper) | GST sigma was the most up-regulated | a | [51] | |
| GST | Triphenyltin | Down | a | [23] | |
| GST | Cypermethrin | Increase | a, d | [22] | |
| GST | Multi-walled carbon nanotubes (MWCNTs) are nanoparticles widely applicable in various industrial fields. | Down | c, d | [24] | |
| GST | Low pH | Increase | a, d | [19] | |
| GST-kappa, GST-mu5, and GST-omega | Toxin-producing dinoflagellate Gymnodinium catenatum | Up | a | [40] | |
| GST | Two diatoms, Chaetoceros muelleri and Nitzschia closterium f. minutissima | No variation | d | [46] | |
| Tigriopus californicus | GSTs | Hydrogen peroxide and paraquat | Up/Down | b | [25] |
| GSTs | Hydrogen peroxide and yeast diet | Up only in males | b | [26] | |
| Paracyclopina nana | GST | Cypermethrin | Increase | a, d | [22] |
| GST | Copper | Increase | d | [10] | |
| GST | Methylmercury | Increase in activity, no variation in gene expression | a, c, d, | [36] | |
| GST | Polystyrene microbeads | Increase | d | [35] | |
| Eurytemora affinis | GST activity | Hydrophobic organic contaminant (HOC) | Increase | d | [27] |
| Calanus finmarchicus | GST | Naphtalene | Up | a | [29] |
| GST | Dispersed oil | Up | a | [30] | |
| GST | Water-accommodated fraction (WAF) of oil | Up | a | [11] | |
| GST | Mercury | Increase | a | [28] | |
| Three cytosolic GSTs and an omega GST | Alexandrium fundyense | Up | b | [39] | |
| Calanus glacialis | GST | Water-accommodated fraction (WAF) of oil | Up | a | [11] |
| GST | Mercury | Increase | a | [28] | |
| Calanus helgolandicus | GST | Skeletonema marinoi | Up-regulation for the Swedish copepod population | a | [42] |
| Microsomal GST3 | Skeletonema marinoi | Up | a, b | [41] | |
| GST | Sampling cruise in the Adriatic Sea (Mediterranean Sea) when a natural diatom bloom was observed | Up | a | [43] | |
| GST | Karenia brevis | Down | a | [44] | |
| Calanus sinicus | GST | Skeletonema marinoi | No variation | a | [45] |
| Calanus pacificus | GST | California West Coast Ocean Acidification Cruise Current System (effect of pH and temperature) | High GST activity | c, d | [49] |
| Centropages ponticus | GST | Cadmium chloride | Increase | d | [31] |
| Centropages tenuiremis | GST | Acidified seawater | No significant differences | c, d | [47] |
| Acartia tonsa | GST-theta and GST-mu | Polycyclic aromatic hydrocarbon (PAH) 1,2-dimethylnaphthalene | Increase | a | [32] |
| GST | Two-week field survey (effect of pH and temperature) | GST activity was higher in week II compared to week I, and, in particular, it was higher in the surface compared to the other depths for both weeks. | c, d | [48] | |
| Acartia bifilosa | GST | Two-week field survey (effect of pH and temperature) | GST activity was higher in week II compared to week I, and, in particular, it was higher in the surface compared to the other depths for both weeks. | c, d | [48] |
| Acartia pacifica | GST | Two diatoms, Chaetoceros muelleri and Nitzschia closterium f. minutissima | Increase with C. muelleri | d | [46] |
| Pseudodiaptomus poplesia | GST delta/epsilon and GST theta | PAH 1,2-dimethylnaphthalene, pyrene | GST delta/epsilon up-regulation for both stressors, GST theta only for 1,2-dimethylnaphthalene | a | [33] |
| Pseudodiaptomus annandalei | GST | Two diatoms, Chaetoceros muelleri and Nitzschia closterium f. minutissima | Increase with both algae | d | [46] |
| Eudiaptomus gracilis | GST | Ultraviolet radiation | Increase | d | [52] |
| Caligus rogercressereyi | GST | Delousing drug deltamethrin (AlphaMaxTM) | Down | b | [37] |
| Lepeophtheirus salmonis | Microsomal glutathione S-transferase 1 | Various lice stages from egg to adult lice | Constitutive transcript level throughout the studied development stages, with the highest levels in the ovaries or gut | a, b | [38] |
2. Materials and Methods
For the copepods whose glutathione S-transferase activity has been investigated (see Table 1), transcriptomic resources were mined in silico to search for putative GST-encoding transcripts. Using an in silico workflow, GSTs from the copepod E. affinis [9] were used as queries to search on NCBI transcriptome shotgun assembly (TSA) database for the following copepods: Calanus helgolandicus (PRJNA640515), C. glacialis (PRJNA237014), Pseudodiaptomus annandalei (PRJNA558682), Acartia tonsa (PRJEB20069), Caligus rogercresseyi (PRJNA234316), and Lepeophtheirus salmonis (PRJNA73431). Searches were performed using tblastn algorithm. For the copepod T. califonicus (PRJNA237968), we mined the collection of proteins (n = 15,557) deposited on the non-redundant protein database (nr) using the blastp algorithm. In spite of species-specific differences, the mined transcriptomes were highly comparable. RNA-Seq was performed from paired-end (PE) reads sequenced on Illumina platforms, with the exception of C. glacialis, which was sequenced on the Ion Torrent plataform (Table S1). Furhermore, the transcriptomes were assembled de novo using Trinity software, with the exception of L. salmonis; the number of transcripts ranged from 36 k to 183 k (Table S1).
To confirm that the identified putative GSTs for the selected species are true members of the GST family, we used a well-established vetting protocol that involved both reciprocal BLAST and structural motif analyses [5]. Briefly, each of the new identified transcripts encoding a putative GST was fully translated using the ‘‘Translate” tool of ExPASy [53] and then the deduced protein was used as the input query for a blastp search of the non-redundant arthropod protein sequences curated at NCBI. Protein structural motifs were analyzed using the online program Pfam [54].
The collection of the identified predicted GSTs from the seven mined transcriptomes was compared to those previously known in literature for Calanus finmarchicus [5], Eurytemora affinis [9], T. japonicus, and Paracyclopina nana [10]. To support the assignment of predicted GST proteins into classes and to establish their relationship to each other and to those from other copepods, an unrooted phylogenetic tree of the cytosolic, microsomal, and mitochindrial GSTs was generated. Amino acid sequences for the identified GST transcripts were aligned using ClustalW software (Galaxy version 2.1; [55]) and FASTTREE was used to build a maximum-likelihood phylogenetic tree (Galaxy Version 2.1.10+galaxy1) using the protein evolution model JTT+ CAT [56].
3. Results
In silico mining of the transcriptome assemblies for the copepods C. helgolandicus, C. glacialis, P. annandalei, A. tonsa, T. californicus, C. rogercresseyi, and L. salmonis resulted in the identification of 14 to 33 transcripts encoding putative GSTs. The highest gene duplication was found for the calanoid C. helgolandicus, followed by the harpacticoid T. californicus (n = 22), while the lowest number of GSTs was found in C. glacialis. In all species, GSTs were identified as members of the cytosolic, microsomal (MAPEG), and mitochondrial classes. The highest number of transcripts encoding GSTs was found in the cytosolic class, which included the delta, sigma, zeta, mu, omega, and theta subclasses. For most of the copepods, the highest diversification was found for the delta and sigma subclasses, with the highest number of transcripts reported in C. helgolandicus with, respectively, 10 sigma and 8 delta GSTs. A single GST was identified in the mitochondrial class, with the exception of L. salmonis, for which two transcripts encoding for kappa GSTs were found. The MAPEG class also included several subclassess, with the high duplication in C. helgolandicus, P. annandalei, and T. californicus. The total number of GSTs and their distributions found for all the seven copepods were highly comparable with the ones reported for the calanoids C. finmarchicus and E. affinis, the harpacticoid T. japonicus, and the cyclopoid P. nana (Figure 2). In spite of some differences, which can also be related to differences among transcriptomes (e.g., depth of sequencing, coverage), the distribution of GSTs was not significantly different between orders (Figure 2).
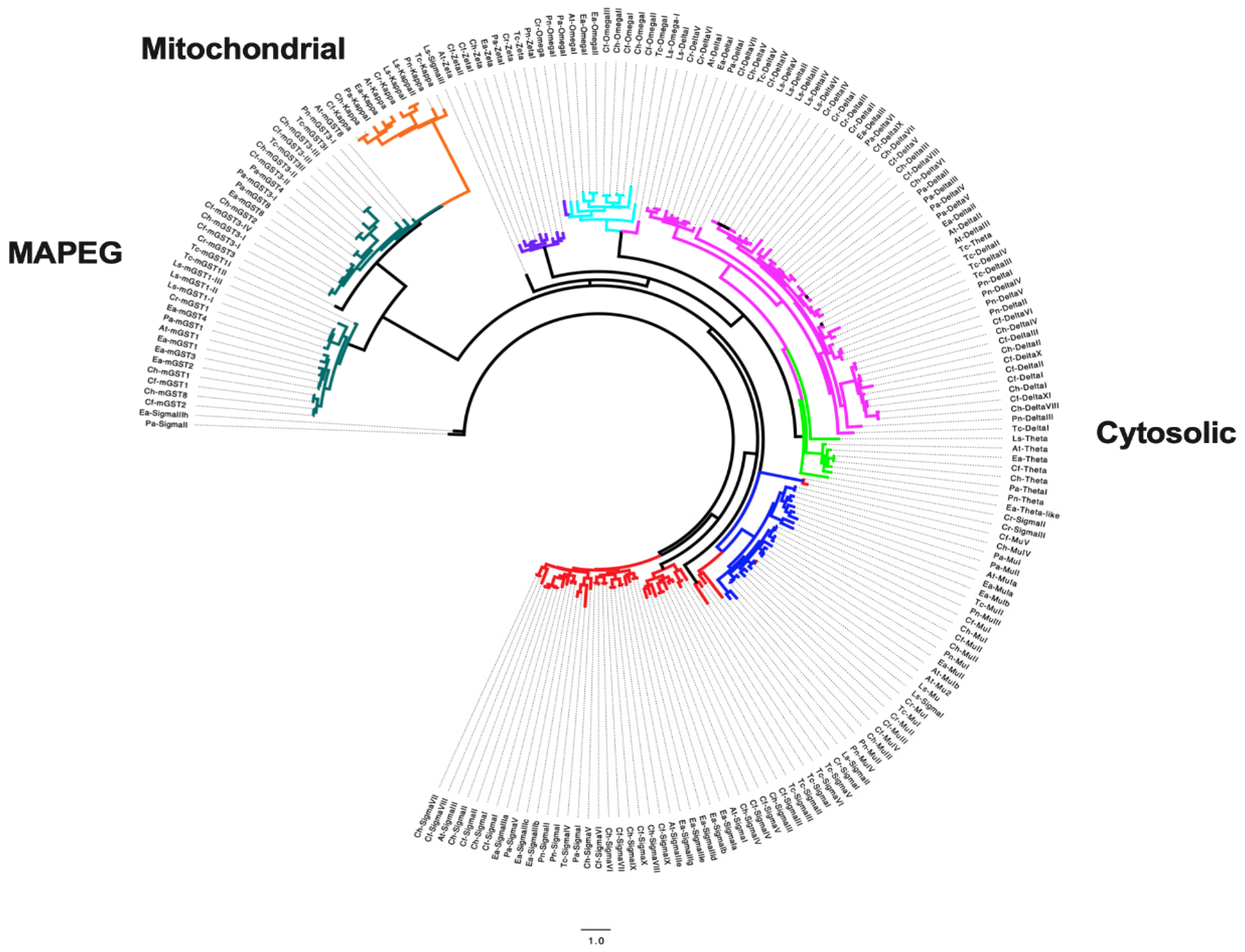
Phylogenetic analysis of the GSTs from all copepods placed the newly identified GSTs into distinct branches (Figure 3), which were consistent with their classification into the cytosolic, microsomal, and mitochondrial subclasses. The microsomal and mitochondrial subclasses clustered into the same branch, suggesting a common functionality. The cytosolic branch separated into different clades, with delta/theta and omega being closely related (Figure 3). The sigma subclass, which is highly diversified, separated from the other cytosolic subclasses (Figure 3). In spite of some “outliers”, the clustering pattern observed for the newly identified GSTs agreed with their annotation. It has to be noted that, as opposed to what has been reported in Roncalli et al. [5], the clustering pattern within individual subclasses varied. While for C. finmarchicus, GSTs from the same subclass were located on a single branch [5], here, for all newly reported GSTs, individual GSTs from the same subclass clustered with the ones from other copepods. Overall, this phylogenetic analysis confirmed the high diversity of the GST family among different orders.
4. Discussion
The glutathione S-transferase is a complex and large gene family present in both prokaryotes and eukaryotes. Based on their cellular localization, GSTs belong to three major classes: cytosolic, microsomal, and mitochondrial. With the exception of the mitochondrial class, the cytosolic and the microsomal classes also include several subclasses. GST members of different classes and/or subclasses have a different function. In insects, the cytosolic delta/epsilon subclass is mostly associated with detoxification, with its genes involved in resistance to pesticides [5]. The sigma class is likely associated with development, while the omega, theta, and zeta sub-classes appear to be involved in other cellular processes, including protection against oxidative stress. The microsomal GSTs are involved in eicosanoid and glutathione metabolism, while still little is known about the function of the mitochondrial GSTs, which seems not exclusively associated with the detoxification response [5]. The complexity of the GST family has been characterized in many insects, with discoveries of high gene duplication, with a total number of GSTs ranging from 13 genes (Apis mellifera) to 41 in Triboleum castaneum [5].
In marine copepods, GSTs have been widely used as indicators of detoxification, however, most of the studies have depended on a single GST transcript used as a biomarker. This was due to the general lack of information about the specific functional roles of different GST protein classes in copepods. As speculated by Choi et al. [9], for example, the higher number of GST kappa genes in the genomes of E. affinis, Tigriopus spp., and P. nana suggests involvement of this class in metabolic pathways, and not only in detoxification. In their study, it was also discussed that microsomal GSTs have been associated to aging, lipid peroxidation, and development in insects, however, more effort is still needed to elucidate the role of these GST classes in invertebrates, and, in particular, in copepods. As for the cytosolic class, up-regulation of sigma and omega GST genes in copepods has been associated with detoxification of hydrophobic xenobiotics (polycyclic aromatic hydrocarbons (PAHs) and polychlorinated biphenyls (PCBs)) and metals [15,57]. This class also plays a role as antioxidants [14], but it is not clear whether this is a common functional feature in all copepod species due to the lack of knowledge on the relationship between GST class-specific gene expression and xenobiotics. Park et al. [10], for example, showed species-specific differences in GSTs’ gene inducibility when comparing T. japonicus and P. nana exposed to copper, with different gene expression modulation in the two species. In the future, to better understand and characterize the biophysical properties, and hence functional diversity, of each GST class, an in-depth analysis of the structural and catalytic properties of GST classes would be necessary in ecotoxicology studies on copepods.
Given the high gene multiplicity in the GST superfamily, these studies may not fully represent the copepods’ detoxification response. In recent years, the development of new genomic ad transcriptomic resources for marine copepods has opened the opportunity to also investigate the GST family in those organisms. Characterization of the GST family would allow a better understanding of the role of the GSTs in detoxification.
The GST gene diversity has been described in the calanoids C. finmarchicus and E. affinis, in the harpacticoids T. japonicus and T. kingsejongensis, and in the cyclopoid P. nana. All these studies confirmed the high diversification of GSTs (19–41 GSTs), with the highest duplication within the cytosolic class. Results of our mining analysis confirmed the high diversity also for the calanoids, P. annandalei, C. helgolandicus, A. tonsa, and C. glacialis, and the siphonostomatoids fish parasites, C. rogercresseyi and L. salmonis. In all copepods, the highest duplication was found in the delta and sigma subclasses with more than 8 transcripts in C. helgolandicus and P. annandalei. This finding is consistent with what has been reported for C. finmarchicus with more than 10 different delta GSTs, suggesting a possible genus-specific complexity of the delta subclass. To our surprise, we did find in P. annandalei and L. salmonis, two different transcripts encoding the mitochondrial kappa gene. The mitochondrial class consists of a single gene in all other copepods, as well as in insects.
GST genomic structural differences and different GST subclass distributions have been found between different copepod orders. In the calanoid E. affinis, a higher number of exons and a longer open reading frame (ORF) was reported compared with the harpacticoids Tigriopus spp. and the cyclopoid P. nana [9]. Furthermore, the authors suggested that the differences found in the distribution of GSTs were associated with the differences in the habitats. A GST species-dependent diversity was reported in Jeong et al. [58] between T. californicus and T. kingsejongensis. The authors suggested that the species-specific expansion was likely due to the different marine habitats and environmental pressures to which the copepods were exposed [58]. Although our investigation has been purely based on transcriptomic resources, we did not find any differences in the number of GSTs and in their class/subclass distribution between different copepod orders and or between different species. Although the overall number of GST did change between species, the class and subclass distributions were similar and, thus, highly comparable.
Differences in GSTs have also been investigated across populations in two Calanus species. In the first study, the complexity of GSTs was compared between C. finmarchicus individuals from the Gulf of Maine and the Norwegian Sea population. In spite of the difference in environment between the two populations, the study reported that the GST diversity was highly conserved, with more than 95% of the genes showing good to excellent amino acid identity (88–100%) between the two transcriptomes [5]. In another study, the expression of GSTs was compared between C. helgolandicus individuals from three different populations, i.e., Swedish western coast, English Channel, and the Adriatic Sea (Mediterranean Sea) [42]. Copepod populations responded differently to the ingestion of the same clone of the diatom Skeletonema marinoi producing oxylipins, secondary metabolites known to induce reduction in egg production and hatching success, teratogenesis, apoptosis, and gene expression alteration in copepods [59,60,61,62,63]. All three Calanus populations co-exist with S. marinoi, but are known to experience different exposure timing along the year. Results demonstrated that the Swedish western coast population of C. helgolandicus was the only one able to activate the major part of the detoxification enzymes, including GST, being the less susceptible to the toxic diet compared to the others. The differences suggested an evolved predator resistance to toxic metabolites in populations with a longer history of exposure to pronounced and long-lasting diatom blooms [42].
Existence of multiple GST transcripts and proteins has been associated with the need to detoxify numerous and diverse xenobiotics or to increase protection against oxidative stress [64]. Different protein isoforms have, in fact, specific xenobiotic affinity and catalytic activity [57]. Moreover, the promoter region of the gamma GST contains a functional xenobiotic response element (XRE), suggesting xenobiotic-specific inducibility of GST genes [65]. High gene diversity may, thus, lead to a more efficient protection of animals against suboptimal environmental conditions due to xenobiotics and xenobiotic-induced oxidative stress. Multidisciplinary approaches combining transcriptomic, proteomic, and enzymatic analyses may help to elucidate this intriguing question in copepods.
In conclusion, characterization of the GST family in marine copepods opens opportunities for functional studies of detoxification and provides a diverse set of biomarkers for these species. The observed heterogeneous response among copepod species, even when exposed to the same stressor, highlights the need to better understand the isoforms investigated, as well as the environment where they have been sampled (e.g., co-evolution with toxic algal species). To date, few studies have reported the specific subclass investigated, thus, future studies should make more effort in selecting the more relevant GST gene in quantitative expression analysis. The development of new biomarkers, which might be achieved by whole transcriptome and genome sequencing combined with functional and enzymatic analyses, will likely be greatly useful for future investigations evaluating ecosystem health status and organism–environment interactions.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jmse9091025/s1, Table S1: Summary of transcriptomic resources mined in silico in the study. Each dataset has a description of the order, family, and species of interest, as well as the NCBI Bioproject accession number. Sequencing platform (all paired-end—PE), assembly software, and number of assembled transcripts are listed.
Author Contributions
Conceptualization, C.L. and V.R.; methodology, C.L. and V.R.; writing—original draft preparation, C.L. and V.R.; writing—review and editing, C.L., V.R. and Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All datasets mined in this study were already publicly available on the National Center for Biotechnology Information (NCBI) in the transcriptome shotgun assembly (TSA) database. For each dataset, Bioproject numbers are provided in Table S1.
Acknowledgments
Authors thank Servier Medical Art (SMART) website (https://smart.servier.com/; accessed on 1 May 2021) by Servier for elements in Figure 1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lauritano, C.; Ianora, A. Chemical Defense in Marine Organisms. Mar. Drugs 2020, 18, 518. [Google Scholar] [CrossRef]
- Townsend, D.M.; Tew, K.D. The Role of Glutathione- S -Transferase in Anti-Cancer Drug Resistance. Oncogene 2003, 22, 7369–7375. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Procaccini, G.; Ianora, A. Gene Expression Patterns and Stress Response in Marine Copepods. Mar. Environ. Res. 2012, 76, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Van Straalen, N.M.; Roelofs, D. An Introduction to Ecological Genomics, 2nd ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2012; ISBN 978-0-19-959468-9. [Google Scholar]
- Roncalli, V.; Cieslak, M.C.; Passamaneck, Y.; Christie, A.E.; Lenz, P.H. Glutathione S-Transferase (GST) Gene Diversity in the Crustacean Calanus finmarchicus—Contributors to Cellular Detoxification. PLoS ONE 2015, 10, e0123322. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.T. Zooplankton Fecal Pellets, Marine Snow, Phytodetritus and the Ocean’s Biological Pump. Prog. Oceanogr. 2015, 130, 205–248. [Google Scholar] [CrossRef]
- Carotenuto, Y.; Vitiello, V.; Gallo, A.; Libralato, G.; Trifuoggi, M.; Toscanesi, M.; Lofrano, G.; Esposito, F.; Buttino, I. Assessment of the Relative Sensitivity of the Copepods Acartia tonsa and Acartia clausi Exposed to Sediment-Derived Elutriates from the Bagnoli-Coroglio Industrial Area. Mar. Environ. Res. 2020, 155, 104878. [Google Scholar] [CrossRef]
- Russo, E.; Ianora, A.; Carotenuto, Y. Re-Shaping Marine Plankton Communities: Effects of Diatom Oxylipins on Copepods and Beyond. Mar. Biol. 2018, 166, 9. [Google Scholar] [CrossRef]
- Choi, B.-S.; Kim, D.-H.; Kim, M.-S.; Park, J.C.; Lee, Y.H.; Kim, H.-J.; Jeong, C.-B.; Hagiwara, A.; Souissi, S.; Lee, J.-S. The Genome of the European Estuarine Calanoid Copepod Eurytemora affinis: Potential Use in Molecular Ecotoxicology. Mar. Pollut. Bull. 2021, 166, 112190. [Google Scholar] [CrossRef]
- Park, J.C.; Lee, M.-C.; Yoon, D.-S.; Han, J.; Park, H.G.; Hwang, U.-K.; Lee, J.-S. Genome-Wide Identification and Expression of the Entire 52 Glutathione S-Transferase (GST) Subfamily Genes in the Cu2+-Exposed Marine Copepods Tigriopus japonicus and Paracyclopina nana. Aquat. Toxicol. 2019, 209, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.H.; Altin, D.; Rørvik, S.F.; Øverjordet, I.B.; Olsen, A.J.; Nordtug, T. Comparative Study on Acute Effects of Water Accommodated Fractions of an Artificially Weathered Crude Oil on Calanus finmarchicus and Calanus glacialis (Crustacea: Copepoda). Sci. Total Environ. 2011, 409, 704–709. [Google Scholar] [CrossRef]
- Han, J.; Won, E.-J.; Hwang, D.-S.; Shin, K.-H.; Lee, Y.S.; Leung, K.M.-Y.; Lee, S.-J.; Lee, J.-S. Crude Oil Exposure Results in Oxidative Stress-Mediated Dysfunctional Development and Reproduction in the Copepod Tigriopus japonicus and Modulates Expression of Cytochrome P450 (CYP) Genes. Aquat. Toxicol. 2014, 152, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-M.; Park, T.-J.; Jung, S.-O.; Seo, J.S.; Park, H.G.; Hagiwara, A.; Yoon, Y.-D.; Lee, J.-S. Cloning and Characterization of Glutathione S-Transferase Gene in the Intertidal Copepod Tigriopus japonicus and Its Expression after Exposure to Endocrine-Disrupting Chemicals. Mar. Environ. Res. 2006, 62, S219–S223. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Lee, K.-W.; Park, H.; Park, H.G.; Raisuddin, S.; Ahn, I.-Y.; Lee, J.-S. Sequence, Biochemical Characteristics and Expression of a Novel Sigma-Class of Glutathione S-Transferase from the Intertidal Copepod, Tigriopus japonicus with a Possible Role in Antioxidant Defense. Chemosphere 2007, 69, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.-S.; Kim, B.-M.; Jeong, C.-B.; Leung, K.M.Y.; Park, G.S.; Lee, J.-S. Development of Enzyme-Linked Immunosorbent Assay (ELISA) for Glutathione S-Transferase (GST-S) Protein in the Intertidal Copepod Tigriopus japonicus and Its Application for Environmental Monitoring. Chemosphere 2013, 93, 2458–2466. [Google Scholar] [CrossRef]
- Wang, M.; Wang, G. Oxidative Damage Effects in the Copepod Tigriopus japonicus Mori Experimentally Exposed to Nickel. Ecotoxicology 2010, 19, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-H.; Wang, G.-Z. Biochemical Response of the Copepod Tigriopus japonicus Mori Experimentally Exposed to Cadmium. Arch. Environ. Contam. Toxicol. 2009, 57, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Lv, D.; Liu, W.; Shen, R.; Li, D.; Hong, H. Accumulation and Developmental Toxicity of Hexabromocyclododecanes (HBCDs) on the Marine Copepod Tigriopus japonicus. Chemosphere 2017, 167, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kang, H.-M.; Kim, M.-S.; Wang, M.; Kim, J.H.; Jeong, C.-B.; Lee, J.-S. Effects of Ocean Acidification on Life Parameters and Antioxidant System in the Marine Copepod Tigriopus japonicus. Aquat. Toxicol. 2019, 212, 186–193. [Google Scholar] [CrossRef]
- Rhee, J.-S.; Lee, Y.-M.; Kim, B.-M.; Leung, K.M.Y.; Kim, I.-C.; Yim, J.H.; Lee, J.-S. β -Naphthoflavone Induces Oxidative Stress in the Intertidal Copepod, Tigriopus japonicus: β-NF-Induced Oxidative Stress in Copepod. Environ. Toxicol. 2015, 30, 332–342. [Google Scholar] [CrossRef]
- Park, J.C.; Han, J.; Lee, M.-C.; Seo, J.S.; Lee, J.-S. Effects of Triclosan (TCS) on Fecundity, the Antioxidant System, and Oxidative Stress-Mediated Gene Expression in the Copepod Tigriopus japonicus. Aquat. Toxicol. 2017, 189, 16–24. [Google Scholar] [CrossRef]
- Zhou, J.; Kang, H.-M.; Lee, Y.H.; Jeong, C.-B.; Park, J.C.; Lee, J.-S. Adverse Effects of a Synthetic Pyrethroid Insecticide Cypermethrin on Life Parameters and Antioxidant Responses in the Marine Copepods Paracyclopina nana and Tigriopus japonicus. Chemosphere 2019, 217, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Yi, A.X.; Han, J.; Lee, J.-S.; Leung, K.M.Y. Ecotoxicity of Triphenyltin on the Marine Copepod Tigriopus japonicus at Various Biological Organisations: From Molecular to Population-Level Effects. Ecotoxicology 2014, 23, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Won, E.-J.; Kang, H.-M.; Hwang, D.-S.; Kim, D.-H.; Kim, R.-K.; Lee, S.-J.; Lee, J.-S. Effects of Multi-Walled Carbon Nanotube (MWCNT) on Antioxidant Depletion, the ERK Signaling Pathway, and Copper Bioavailability in the Copepod (Tigriopus japonicus). Aquat. Toxicol. 2016, 171, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Flanagan, B.A.; Partridge, M.; Huang, E.J.; Edmands, S. Sex Differences in Early Transcriptomic Responses to Oxidative Stress in the Copepod Tigriopus californicus. BMC Genom. 2020, 21, 759. [Google Scholar] [CrossRef]
- Li, N.; Arief, N.; Edmands, S. Effects of Oxidative Stress on Sex-Specific Gene Expression in the Copepod Tigriopus californicus Revealed by Single Individual RNA-Seq. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100608. [Google Scholar] [CrossRef] [PubMed]
- Cailleaud, K.; Forget-Leray, J.; Peluhet, L.; LeMenach, K.; Souissi, S.; Budzinski, H. Tidal Influence on the Distribution of Hydrophobic Organic Contaminants in the Seine Estuary and Biomarker Responses on the Copepod Eurytemora affinis. Environ. Pollut. 2009, 157, 64–71. [Google Scholar] [CrossRef]
- Øverjordet, I.B.; Altin, D.; Berg, T.; Jenssen, B.M.; Gabrielsen, G.W.; Hansen, B.H. Acute and Sub-Lethal Response to Mercury in Arctic and Boreal Calanoid Copepods. Aquat. Toxicol. 2014, 155, 160–165. [Google Scholar] [CrossRef]
- Hansen, B.H.; Altin, D.; Vang, S.-H.; Nordtug, T.; Olsen, A.J. Effects of Naphthalene on Gene Transcription in Calanus finmarchicus (Crustacea: Copepoda). Aquat. Toxicol. 2008, 86, 157–165. [Google Scholar] [CrossRef]
- Hansen, B.H.; Nordtug, T.; Altin, D.; Booth, A.; Hessen, K.M.; Olsen, A.J. Gene Expression of GST and CYP330A1 in Lipid-Rich and Lipid-Poor Female Calanus finmarchicus (Copepoda: Crustacea) Exposed to Dispersed Oil. J. Toxicol. Environ. Health A 2009, 72, 131–139. [Google Scholar] [CrossRef]
- Ensibi, C.; Yahia, M.N.D. Toxicity Assessment of Cadmium Chloride on Planktonic Copepods Centropages ponticus Using Biochemical Markers. Toxicol. Rep. 2017, 4, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, B.; Zeng, S.; Gong, Z.; Jing, F.; Zhang, J. Glutathione S-Transferase (GST) Genes from Marine Copepods Acartia tonsa: CDNA Cloning and MRNA Expression in Response to 1,2-Dimethylnaphthalene. Aquat. Toxicol. 2020, 224, 105480. [Google Scholar] [CrossRef]
- Zhuang, Y.; Yang, F.; Xu, D.; Chen, H.; Zhang, H.; Liu, G. Spliced Leader-Based Analyses Reveal the Effects of Polycyclic Aromatic Hydrocarbons on Gene Expression in the Copepod Pseudodiaptomus poplesia. Aquat. Toxicol. 2017, 183, 114–126. [Google Scholar] [CrossRef]
- Beiras, R.; Tato, T.; López-Ibáñez, S. A 2-Tier Standard Method to Test the Toxicity of Microplastics in Marine Water Using Paracentrotus lividus and Acartia clausi Larvae. Environ. Toxicol. Chem. 2019, 38, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.-B.; Kang, H.-M.; Lee, M.-C.; Kim, D.-H.; Han, J.; Hwang, D.-S.; Souissi, S.; Lee, S.-J.; Shin, K.-H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina nana. Sci. Rep. 2017, 7, 41323. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Kim, D.-H.; Kang, H.-M.; Wang, M.; Jeong, C.-B.; Lee, J.-S. Adverse Effects of Methylmercury (MeHg) on Life Parameters, Antioxidant Systems, and MAPK Signaling Pathways in the Rotifer Brachionus koreanus and the Copepod Paracyclopina nana. Aquat. Toxicol. 2017, 190, 181–189. [Google Scholar] [CrossRef]
- Chávez-Mardones, J.; Gallardo-Escárate, C. Next-Generation Transcriptome Profiling of the Salmon Louse Caligus rogercresseyi Exposed to Deltamethrin (AlphaMaxTM): Discovery of Relevant Genes and Sex-Related Differences. Mar. Biotechnol. 2015, 17, 793–810. [Google Scholar] [CrossRef]
- Dalvin, S.; Eichner, C.; Dondrup, M.; Øvergård, A.-C. Roles of Three Putative Salmon Louse (Lepeophtheirus salmonis) Prostaglandin E2 Synthases in Physiology and Host–Parasite Interactions. Parasites Vectors 2021, 14, 206. [Google Scholar] [CrossRef]
- Roncalli, V.; Turner, J.T.; Kulis, D.; Anderson, D.M.; Lenz, P.H. The Effect of the Toxic Dinoflagellate Alexandrium fundyense on the Fitness of the Calanoid Copepod Calanus finmarchicus. Harmful Algae 2016, 51, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Park, J.S.; Park, Y.; Lee, J.; Shin, H.H.; Lee, K.-W. Effects of Paralytic Shellfish Poisoning Toxin-Producing Dinoflagellate Gymnodinium catenatum on the Marine Copepod Tigriopus japonicus. Mar. Pollut. Bull. 2021, 163, 111937. [Google Scholar] [CrossRef] [PubMed]
- Asai, S.; Sanges, R.; Lauritano, C.; Lindeque, P.K.; Esposito, F.; Ianora, A.; Carotenuto, Y. De Novo Transcriptome Assembly and Gene Expression Profiling of the Copepod Calanus helgolandicus Feeding on the PUA-Producing Diatom Skeletonema Marinoi. Mar. Drugs 2020, 18, 392. [Google Scholar] [CrossRef]
- Lauritano, C.; Carotenuto, Y.; Miralto, A.; Procaccini, G.; Ianora, A. Copepod Population-Specific Response to a Toxic Diatom Diet. PLoS ONE 2012, 7, e47262. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Romano, G.; Roncalli, V.; Amoresano, A.; Fontanarosa, C.; Bastianini, M.; Braga, F.; Carotenuto, Y.; Ianora, A. New Oxylipins Produced at the End of a Diatom Bloom and Their Effects on Copepod Reproductive Success and Gene Expression Levels. Harmful Algae 2016, 55, 221–229. [Google Scholar] [CrossRef]
- Lauritano, C.; Carotenuto, Y.; Procaccini, G.; Turner, J.T.; Ianora, A. Changes in Expression of Stress Genes in Copepods Feeding upon a Non-Brevetoxin-Producing Strain of the Dinoflagellate Karenia brevis. Harmful Algae 2013, 28, 23–30. [Google Scholar] [CrossRef]
- Lauritano, C.; Carotenuto, Y.; Vitiello, V.; Buttino, I.; Romano, G.; Hwang, J.-S.; Ianora, A. Effects of the Oxylipin-Producing Diatom Skeletonema marinoi on Gene Expression Levels of the Calanoid Copepod Calanus sinicus. Mar. Genom. 2015, 24, 89–94. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, G.; Zeng, C.; Wu, L. Comparative Study on the Effects of Two Diatoms as Diets on Planktonic Calanoid and Benthic Harpacticoid Copepods. J. Exp. Zool. Part Ecol. Integr. Physiol. 2018, 329, 140–148. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Wang, G.; Guo, D.; Xing, K.; Zhang, S. Biochemical Responses of the Copepod Centropages tenuiremis to CO2-Driven Acidified Seawater. Water Sci. Technol. 2012, 65, 30–37. [Google Scholar] [CrossRef]
- Glippa, O.; Engström-Öst, J.; Kanerva, M.; Rein, A.; Vuori, K. Oxidative Stress and Antioxidant Defense Responses in Acartia Copepods in Relation to Environmental Factors. PLoS ONE 2018, 13, e0195981. [Google Scholar] [CrossRef] [Green Version]
- Engström-Öst, J.; Glippa, O.; Feely, R.A.; Kanerva, M.; Keister, J.E.; Alin, S.R.; Carter, B.R.; McLaskey, A.K.; Vuori, K.A.; Bednaršek, N. Eco-Physiological Responses of Copepods and Pteropods to Ocean Warming and Acidification. Sci. Rep. 2019, 9, 4748. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-M.; Choi, B.-S.; Lee, K.-W.; Ki, J.-S.; Kim, I.-C.; Choi, I.-Y.; Rhee, J.-S.; Lee, J.-S. Expression Profile Analysis of Antioxidative Stress and Developmental Pathway Genes in the Manganese-Exposed Intertidal Copepod Tigriopus japonicus with 6K Oligochip. Chemosphere 2013, 92, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Lee, K. Expression of Glutathione S-Transferase (GST) Genes in the Marine Copepod Tigriopus japonicus Exposed to Trace Metals. Aquat. Toxicol. 2008, 89, 158–166. [Google Scholar] [CrossRef]
- Souza, M.S.; Hansson, L.-A.; Hylander, S.; Modenutti, B.; Balseiro, E. Rapid Enzymatic Response to Compensate UV Radiation in Copepods. PLoS ONE 2012, 7, e32046. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Apple, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Park, J.C.; Hagiwara, A.; Park, H.G.; Lee, J.-S. The Glutathione S-Transferase Genes in Marine Rotifers and Copepods: Identification of GSTs and Applications for Ecotoxicological Studies. Mar. Pollut. Bull. 2020, 156, 111080. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.-B.; Lee, B.-Y.; Choi, B.-S.; Kim, M.-S.; Park, J.C.; Kim, D.-H.; Wang, M.; Park, H.G.; Lee, J.-S. The Genome of the Harpacticoid Copepod Tigriopus japonicus: Potential for Its Use in Marine Molecular Ecotoxicology. Aquat. Toxicol. 2020, 222, 105462. [Google Scholar] [CrossRef]
- Lauritano, C.; Borra, M.; Carotenuto, Y.; Biffali, E.; Miralto, A.; Procaccini, G.; Ianora, A. Molecular Evidence of the Toxic Effects of Diatom Diets on Gene Expression Patterns in Copepods. PLoS ONE 2011, 6, e26850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Borra, M.; Carotenuto, Y.; Biffali, E.; Miralto, A.; Procaccini, G.; Ianora, A. First Molecular Evidence of Diatom Effects in the Copepod Calanus helgolandicus. J. Exp. Mar. Biol. Ecol. 2011, 404, 79–86. [Google Scholar] [CrossRef]
- Carotenuto, Y.; Esposito, F.; Pisano, F.; Lauritano, C.; Perna, M.; Miralto, A.; Ianora, A. Multi-Generation Cultivation of the Copepod Calanus helgolandicus in a Re-Circulating System. J. Exp. Mar. Biol. Ecol. 2012, 418–419, 46–58. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A. Toxigenic Effects of Diatoms on Grazers, Phytoplankton and Other Microbes: A Review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, Y.; Dattolo, E.; Lauritano, C.; Pisano, F.; Sanges, R.; Miralto, A.; Procaccini, G.; Ianora, A. Insights into the Transcriptome of the Marine Copepod Calanus helgolandicus Feeding on the Oxylipin-Producing Diatom Skeletonema marinoi. Harmful Algae 2014, 31, 153–162. [Google Scholar] [CrossRef] [PubMed]
- da Fonseca, R.R.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Molecular Evolution and the Role of Oxidative Stress in the Expansion and Functional Diversification of Cytosolic Glutathione Transferases. BMC Evol. Biol. 2010, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favreau, L.V.; Pickett, C.B. Transcriptional Regulation of the Rat NAD(P)H:Quinone Reductase Gene. Identification of Regulatory Elements Controlling Basal Level Expression and Inducible Expression by Planar Aromatic Compounds and Phenolic Antioxidants. J. Biol. Chem. 1991, 266, 4556–4561. [Google Scholar] [CrossRef]
Figure 1.
Graphical representation of the glutathione S-transferase genes based on their cellular localization. GSTs belong to three classes (cytosolic, mitochondrial, and microsomal). The cytosolic class includes several subclasses as listed.
Figure 1.
Graphical representation of the glutathione S-transferase genes based on their cellular localization. GSTs belong to three classes (cytosolic, mitochondrial, and microsomal). The cytosolic class includes several subclasses as listed.
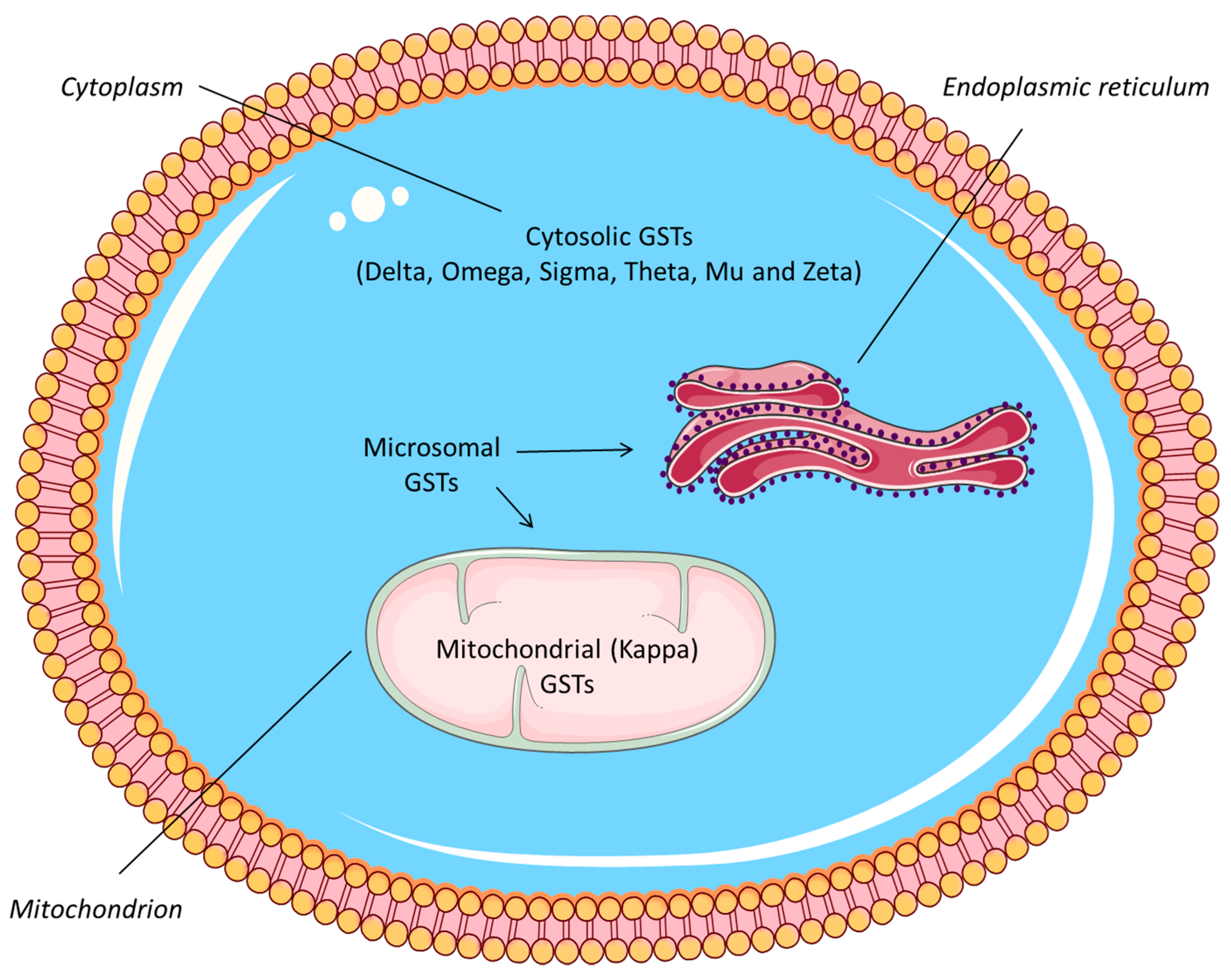
Figure 2.
Glutathione S-transferase diversity in marine copepods. Number of transcripts encoding for glutathione S-transferases from the cytosolic, mitochondrial, and microsomal (MAPEG) class. GST diversity identified in this study (*) for the copepods C. helgolandicus, C. glacialis, P. annandalei, A. tonsa, T. californicus, C. rogercresseyi, and L. salmonis compared with the GST diversity previously reported in literature for C. finmarchicus [5], E. affinis [9], T. japonicus, and P. nana [10]). In bold, the three GST classes: cytosolic, mitochondrial (kappa), and microsomal. The cytosolic class includes multiple subclasses as labeled.
Figure 2.
Glutathione S-transferase diversity in marine copepods. Number of transcripts encoding for glutathione S-transferases from the cytosolic, mitochondrial, and microsomal (MAPEG) class. GST diversity identified in this study (*) for the copepods C. helgolandicus, C. glacialis, P. annandalei, A. tonsa, T. californicus, C. rogercresseyi, and L. salmonis compared with the GST diversity previously reported in literature for C. finmarchicus [5], E. affinis [9], T. japonicus, and P. nana [10]). In bold, the three GST classes: cytosolic, mitochondrial (kappa), and microsomal. The cytosolic class includes multiple subclasses as labeled.
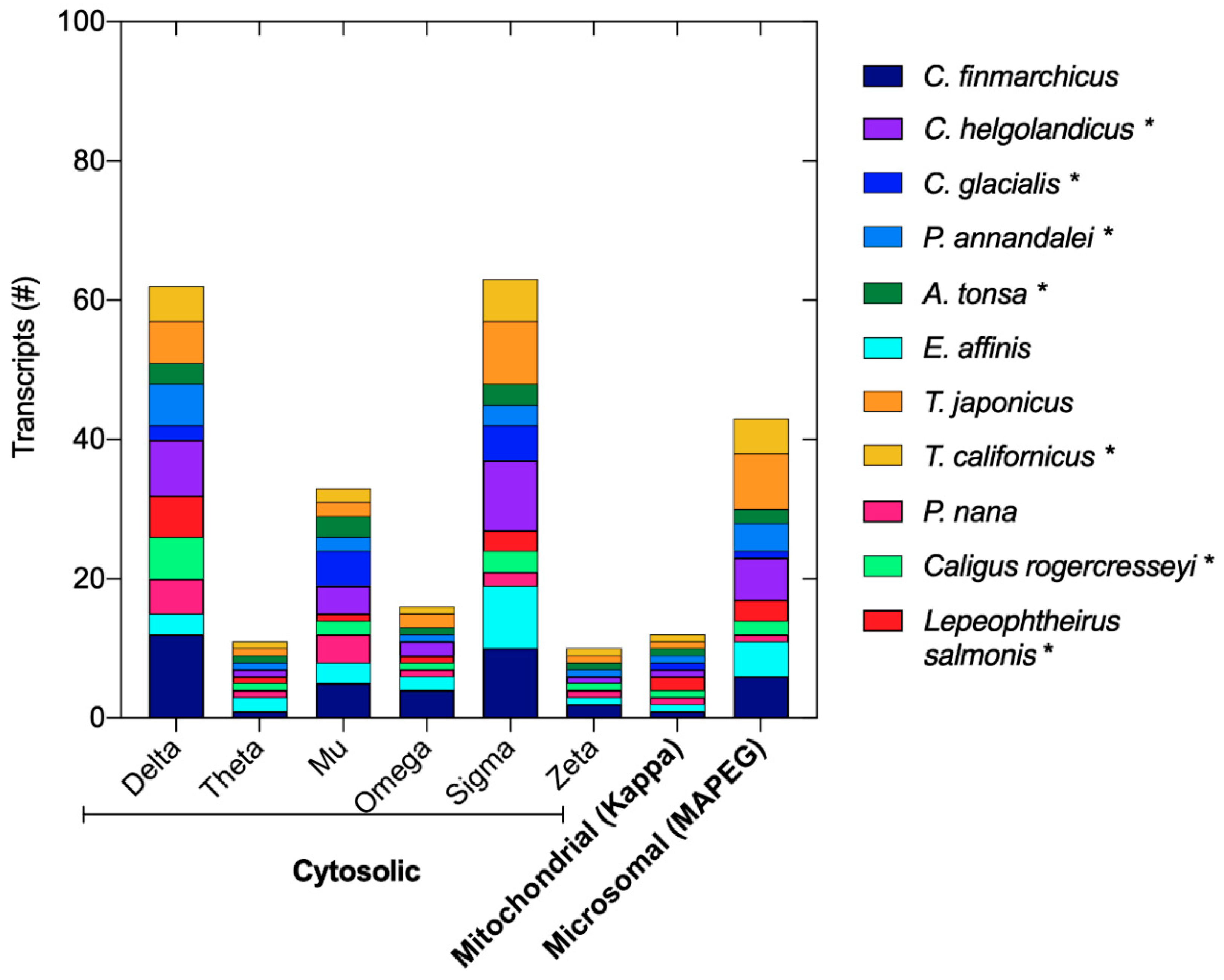
Figure 3.
Phylogenetic tree for the glutathione S-transferases in marine copepods. The unrooted tree shows the relationships between the cytosolic, microsomal (MAPEG), and mitochondrial GSTs from the copepods C. finmarchicus, C. helgolandicus, P. annandalei, A. tonsa, E. affinis, T. californicus, T. japonicus, P. nana, C. rogercressereyi, and L. salmonis. FAST TREE was used to build maximum-likelihood phylogenetic tree using the protein evolution model JTT + CAT. Color coding: teal = microsomal (MAPEG), orange = mitochondrial, red = sigma, blue = mu, light green = theta, pink = delta, turquoise = omega, purple = zeta.
Figure 3.
Phylogenetic tree for the glutathione S-transferases in marine copepods. The unrooted tree shows the relationships between the cytosolic, microsomal (MAPEG), and mitochondrial GSTs from the copepods C. finmarchicus, C. helgolandicus, P. annandalei, A. tonsa, E. affinis, T. californicus, T. japonicus, P. nana, C. rogercressereyi, and L. salmonis. FAST TREE was used to build maximum-likelihood phylogenetic tree using the protein evolution model JTT + CAT. Color coding: teal = microsomal (MAPEG), orange = mitochondrial, red = sigma, blue = mu, light green = theta, pink = delta, turquoise = omega, purple = zeta.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lauritano, C.; Carotenuto, Y.; Roncalli, V. Glutathione S-Transferases in Marine Copepods. J. Mar. Sci. Eng. 2021, 9, 1025. https://doi.org/10.3390/jmse9091025
AMA Style
Lauritano C, Carotenuto Y, Roncalli V. Glutathione S-Transferases in Marine Copepods. Journal of Marine Science and Engineering. 2021; 9(9):1025. https://doi.org/10.3390/jmse9091025
Chicago/Turabian StyleLauritano, Chiara, Ylenia Carotenuto, and Vittoria Roncalli. 2021. "Glutathione S-Transferases in Marine Copepods" Journal of Marine Science and Engineering 9, no. 9: 1025. https://doi.org/10.3390/jmse9091025
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.