
Transcriptome Analysis Reveals the Endocrine Regulation on the Expression of IAG in Litopenaeus vannamei


1
Key Laboratory of Experimental Marine Biology, Institute of Oceanology Chinese Academy of Sciences, Qingdao 266071, China
2
Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266000, China
3
University of Chinese Academy of Sciences, Beijing 100049, China
4
Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
5
Department of Life Sciences and the National Institute for Biotechnology in the Negev, Ben-Gurion University of the Negev, Beersheba 84105, Israel
6
The Innovation of Seed Design, Chinese Academy of Sciences, Wuhan 430072, China
*
Authors to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2021, 9(6), 677; https://doi.org/10.3390/jmse9060677
Submission received: 30 April 2021
/
Revised: 29 May 2021
/
Accepted: 16 June 2021
/
Published: 19 June 2021
(This article belongs to the Section Marine Biology)
Abstract
:The insulin-like androgenic gland hormone (IAG) plays a key role in male sexual differentiation and spermatogenesis in crustaceans. The expression of IAG is usually negatively regulated by neuropeptide hormones through the “eyestalk-AG” endocrine axis. However, the underlying mechanism is still largely unknown. In the present study, we performed a comparative transcriptome analysis on the androgenic gland (AG), the main secretory organ of IAG, of L. vannamei before and after unilateral eyestalk ablation. A total of 67 differentially expressed genes (DEGs) were identified, including some putative genes involved in sexual development. Interestingly, several genes related to molting and endocrine processes were found differentially expressed between the two treatments. Further investigation on the expression profiles of these genes and Lv-IAG showed that their expression patterns were closely correlated throughout the molting cycle. In addition, injection of 20-hydroxyecdysone (20E) could inhibit the expression of Lv-IAG at different molting stages in a dosage-dependent manner. The data indicated that ecdysteroids played important roles in regulating the Lv-IAG expression and might bridge the endocrine axis between eyestalk and AG. The present study provided new insights into understanding the regulation of sexual development in male crustacean.
1. Introduction
Sexual dimorphism is a common phenomenon in decapod crustaceans [1]. Litopenaeus vannamei is one of the most successful commercial crustaceans in the world due to its high growth rate, resistance to diseases, and environmental stress, and adaptability to low salinity cultivation [2]. The female L. vannamei shows obvious superiority in growth rate comparing to male shrimp after sexual maturity [3]. Therefore, clarifying the molecular mechanism of shrimp sex determination and sex differentiation is essential for the development of its monosex culture techniques.
In crustacean, the androgenic gland (AG) is an essential player in male sex differentiation processes. The insulin-like androgenic gland hormone (IAG), which is secreted by AG, is regarded as the sexual “IAG-switch” during sex differentiation of crustaceans [4]. The silencing of IAG could induce full functional sex reversal in Macrobrachium rosenbergii [5]. Except for its function in sex differentiation at the early developmental stage, IAG also plays important roles in the maintenance of male characters and spermatogenesis at the adult stage of male crustaceans [6,7,8,9]. The implantation of AG or injection of an active extract of AG induces masculinization and inhibits the development of female secondary sex characteristics in crustaceans [10,11].
Although several homologs in the sex determination pathway, including Dsx, Sxl, Dmrt11E, and Sox9, have been identified to participate in IAG expression in some crustaceans [12,13,14], it seems that the expression of IAG is mainly regulated by the “eyestalk-AG-testis” endocrine system [15]. Eyestalk ablation could cause hypertrophy and alter the cell types of AG and enhance RNA synthesis in AG cells of Pandalus platyceros, L. vannamei, Portunus pelagicus, Macrobrachium rosenbergii, and Cherax quadricarinatus [16,17,18,19]. Additionally, eyestalk ablation could also greatly promote the expression of IAG in Callinectes sapidus and Portunus trituberculatus [17,20].
The X-organ/sinus-gland (XO-SG) complex, located in the eyestalk of crustaceans, works as the center of the neuroendocrine system to regulate various physiological processes, including metabolism, molting cycle, sexual differentiation, and reproductive development [4,21,22]. Recently, several neuro-hormones secreted by the XO-SG complex, such as the crustacean hyperglycemic hormones (CHHs) [23,24], crustacean female sex hormone (CFSH) [25,26], gonad-inhibiting hormone (GIH), and molt-inhibiting hormone (MIH) [27], have been proved to be negative regulatory factors of IAG expression. As the XO-SG complex locates in the eyestalk, while the AG attaches to the distal vas deferens and terminal ampoule, there should be an efficient regulatory strategy to achieve an accurate regulation. However, the underlying molecular mechanism is still largely unknown.
In order to learn how the factors inside the eyestalk affect the expression of IAG, we performed a comparative transcriptome analysis on AG of L. vannamei after unilateral eyestalk ablation. We found that some genes related to molting and endocrine processes showed differential expressions in AG of shrimp after eyestalk ablation, as well as several putative genes involved in male sexual development. Further study showed that IAG expression in AG could be regulated by ecdysteroids. The data indicated the important roles of ecdysteroids in regulating Lv-IAG expression. These results might bridge the endocrine axis between eyestalk and AG, which will provide new insights into understanding the regulation of sex differentiation in male crustaceans.
2. Results
2.1. Transcriptome Data Mapping and Analysis
Six samples, including eAG1, eAG2, eAG3, cAG1, cAG2, and cAG3, were sequenced on the Illumina Hiseq 2500 sequencing platform. A total of 114,939,060 and 126,419,470 raw reads from a unilateral eyestalk ablation (ESA) group and a control group were obtained by RNA-sequencing, respectively. After filtering low-quality reads and adaptors, 113,214,646 and 123,993,288 clean reads were obtained in each group. About 87.85% and 87.76% of the total clean reads from two groups were mapped to the reference genome (Genebank accession: PRJNA438564). The sequencing and mapping output of all samples are summarized in Table 1.
2.2. Identification of Differentially Expressed Genes (DEGs)
As shown in Figure 1A, 67 genes were identified with a significant difference (p < 0.05) between the two groups, including 45 up-regulated genes and 22 down-regulated genes, respectively. The gene accession number, annotation, and differential expression changes of all the identified DEGs are summarized in Table S1. Among these DEGs, 61 genes could be annotated with known functions. As expected, the expression level of Lv-IAG in the ESA group was significantly higher (Table S1) than that in the control group, which was consistent with the expression trends of Lv-IAG in the two groups detected by qPCR analysis (Figure 1B). The data showed that the identified DEGs were reliable.
2.3. Identification of DEGs Related to Male Differentiation
Based on the functional annotation and published literature, 20 DGEs with annotation were identified as putative target genes related to sexual development and endocrine system (Table 2). Among them, most DEGs related to sexual development were up-regulated after the unilateral eyestalk ablation, like the expression trend of Lv-IAG. Only two DEGs in this category, zinc finger and SCAN domain-containing protein 31 and zinc finger/CW type with PWWP domain 1, were down-regulated after unilateral eyestalk ablation. Eleven DEGs related to endocrinology were identified, including the molting-related hormone receptor 4 (Lv-Hr4), several dopamine-metabolism-related genes, and one co-receptor of the androgen receptor proliferation-associated protein 2G4. Lv-Hr4 was down-regulated, whereas all dopamine-metabolism-related genes were up-regulated.
2.4. Expression Profiles of Selected Genes at Different Molt Stages
As the transcriptome data showed that some genes related to endocrine, especially the molting process, were identified as DEGs, we firstly detected the expression pattern of Lv-IAG in AG of the shrimp throughout the molting cycle. As shown in Figure 2, the expression of Lv-IAG showed the highest level at the late pre-molt stage (D3–4) and rapidly declined at the post-molt stage (P1–2) and inter-molt stage (C), whereas it started to increase at the early and middle pre-molt stage (D0–2).
In order to know the expression correlation between Lv-IAG and endocrine-related DEGs, we detected the expression profiles of Lv-Hr4 and three dopamine beta-hydroxylase genes (Lv-DBH1, Lv-DBH2, Lv-DBH3) throughout the molting cycle. As shown in Figure 2, the expression pattern of Lv-Hr4 showed almost an opposite trend. It showed the highest expression level at the inter-molt stage, declined to the lowest level at the late pre-molt stage, and started to increase at the post-molt stage. The expression of three Lv-DBH genes showed similar expression profiles as that of Lv-IAG.
2.5. Lv-IAG Expression Profile in Molting Cycle with 20E Treatment
As the expression trends of endocrine-related genes were closely related to the expression pattern of Lv-IAG throughout the molting cycle, we further detected whether a regulation relation existed between the molting and male development. The expression level of the internal gene 18S rRNA among all groups was shown in Figure S1. The results showed that a low dosage (1 μg) of 20E injection could inhibit the expression of Lv-IAG at pre-molt and post-molt stages, and high dosage (5 μg) of 20E injection could significantly decrease the expression levels of Lv-IAG throughout the whole molting cycle (Figure 3A). As the ecdysteroids activating gene, Lv-Hr4 was positively regulated by 20E injection at different molting stages (Figure 3B).
3. Discussion
More and more evidence indicates that the “eyestalk-AG-testis” endocrine axis in crustaceans plays an important role in regulating male sexual development by secreting peptide hormones from the X-organ/sinus-gland (XO-SG) complex of eyestalk [15]. However, it is not clear whether these neurohormones directly function on AG. In order to know how the eyestalk affects Lv-IAG expression, we performed a comparative transcriptome analysis between AG tissues in shrimp before and after unilateral eyestalk ablation to identify genes related to Lv-IAG expression inside the AG tissue.
Some identified DEGs have been reported to be involved in sex development in crustaceans or other animals. Besides Lv-IAG, two genes encoding male reproductive-related (Mrr) protein A were also up-regulated after unilateral eyestalk ablation. Mar-Mrr was identified as a sex-specific gene that was first detected in the epithelial cells of the sperm duct and terminal ampulla of M. rosenbergii [28] and later found to be more likely involved in acrosomal activation during fertilization in P. pelagicus [29]. Three heat shock proteins were also up-regulated upon unilateral eyestalk ablation, which have been reported to be involved in sexual differentiation of medaka since they could induce cortisol levels by elevating the expression of the corticotropin-releasing hormone b in the hypothalamus [30].
Two genes related to vitamin K metabolism, vitamin K-dependent gamma-carboxylase, and calumenin were also positively regulated by unilateral eyestalk ablation. Vitamin K-dependent gamma-carboxylase is a kind of enzyme catalyzing γ-carboxylation with vitamin K as a co-factor, which is evolutionary conservative and widely found in both vertebrates and invertebrates. In the lobster Sagmariasus verreauxi, Vitamin K-dependent gamma-carboxylase showed an AG-biased expression profile [31]. In fish, it is exclusively expressed in pituitary and gonad tissues, indicating its function in reproduction [32,33]. In mice, it is essential for epididymal sperm maturation [34]. The data showed that all the above-mentioned genes exhibited up-regulation trends like Lv-IAG after the inhibitory effect of eyestalk decreased by unilateral eyestalk ablation. These results indicated that these genes, like Lv-IAG, might be regulatory targets of the “eyestalk-AG” endocrine axis.
According to the endocrine axis hypothesis, some genes related to the transduction of extracellular signals from eyestalk should have existed in the androgenic gland. Interestingly, several genes related to endocrinology were differentially expressed after unilateral eyestalk ablation. Lv-Hr4 encodes a downstream nuclear receptor of the ecdysteroid signaling transition [35,36], and its expression could be induced by ecdysteroids. Ecdysteroids are a kind of steroid hormone and play important roles in many biological processes including molting and reproduction in arthropods. In crustaceans, ecdysteroids are secreted by the Y-organ, and the process is mainly under the control of neuropeptide hormones, including MIH and CHH, which are produced in the XO-SG complex [37,38,39,40]. The coordination of reproduction and molt exists both in the female decapods [41] and in male ones. In the spider crab Libinia emarginat, ecdysteroids were mainly found in the testis of non-reproductive unabraded males rather than that in the actively mating abraded ones, indicating the role of ecdysteroids in gonad maturation in unabraded animals [42]. In L. vannamei, the reproductive cycle of male individuals is strictly associated with their molting cycle [43]. These data suggest that a coordination mechanism probably exists in shrimps to regulate the relationship between the molt cycle and male sexual development process, which is also supported by the expression profile of Lv-IAG and the inhibitory effect of 20E on Lv-IAG expression throughout the whole molting cycle.
In crustaceans, the ecdysteroids level changes periodically along with the molting process. Starting from basal levels at the post-molt stage, the level of ecdysteroids increases rapidly at the early pre-molt stage, followed by a sharp decline back to basal levels prior to ecdysis [44,45]. In the present study, Lv-IAG was mainly detected at the late pre-molt stage, which is also the time when ecdysteroids decline. The hemolymph titer of MIH, the inhibitory neuropeptide for both ecdysteroids synthesis and IAG expression, was decreased to the lowest level at the early pre-molt stage but kept at high levels at other molting stages in Procambarus clarkii [45]. Accordingly, the decrease in neuropeptide hormones secreted by the X-organ/sinus gland complex activates synthesis of ecdysteroids in the Y-organ, while the decline in ecdysteroids levels leads to the high expression of IAG in the AG tissue (Figure 4A). Therefore, we deduce that ecdysteroids might mediate the inhibitory regulation of neuropeptide hormones on the expression of Lv-IAG. In other words, an “X-organ/sinus gland–Y-organ–AG” endocrine axis (Figure 4B) should exist in shrimp to regulate the IAG expression and the subsequent male sexual differentiation. Further investigations are necessary to provide more evidence on the hypothesis.
4. Materials and Methods
4.1. Experimental Animals
Healthy adult male shrimps were bought from an aquaculture farm nearby Qingdao city and kept in the aerated aquarium of our laboratory at a temperature of 25 °C for at least one week before experiments. Different sizes of shrimp were used since different experiments were not performed at the same time. Male individuals with a bodyweight of 10.5 ± 1.5 g were used for a unilateral eyestalk ablation experiment, male individuals with a bodyweight of 7.0 ± 1.0 g were used for collecting AG tissues at different molting stages, and male shrimps with a bodyweight of 9.0 ± 0.8 g were used in the 20-hydroxyecdysone (20E) injection experiment.
4.2. Unilateral Eyestalk Ablation
Eighty male shrimp individuals at the intermolt stage were used and separated into two groups: the unilateral eyestalk ablation (ESA) group and the control group. In the ESA group, the left eyestalk of shrimp was ablated. The AG tissue of shrimp in two groups was collected individually at 7 days after the ablation treatment.
4.3. Total RNA Preparation and cDNA Synthesis
The total RNA from each AG sample was extracted with RNAisol reagent (Takara, Japan) using the recommended manufacturer’s instruction. The quality of RNA samples was identified by NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and electrophoresis on 1% agarose gel. One μg total RNA from each sample was used for cDNA synthesis. The cDNA was synthesized following the manufacturer’s protocol of the PrimeScripxt RT reagent Kit with a gDNA Eraser kit (Takara, Japan).
4.4. RNA-Sequencing
Before performing the RNA-sequence, the expression levels of Lv-IAG in all AG samples were detected via RT-qPCR and calculated according to the 2−ΔΔCt method [46]. After analyzing the relative expression of Lv-IAG in all AG samples, nine samples with high Lv-IAG expression levels in the ESA group and nine samples with low Lv-IAG expression levels in the control group were selected, respectively. Then the equal amount of total RNA from three samples in each group was mixed, and three biological replicates were prepared for each group. Three replicates from the ESA group were named eAG1, eAG2, and eAG3, and three replicates from the control group were named cAG1, cAG2, and cAG3.
The quantity and integrity of RNA samples were evaluated by Qubit2.0 Fluorometer and Agilent 2100 Bioanalyzer. Then, mRNA was enriched using magnetic beads with Oligo(dT) and broken into short fragments by ultrasound. The first-strand cDNA was synthesized in an M-MuLV reverse transcriptase system with random oligonucleotides as primers. Then the second cDNA strand was synthesized in the DNA polymerase I system after the RNA strands were eliminated by RNaseH. After purification, end repairment Poly(A) tail was added and connected to Illumina sequencing adapters. The ligation products were size selected by agarose gel electrophoresis, PCR amplified, and sequenced on the Illumina Hiseq 2500 by Gene Denovo Biotechnology Co., Ltd. (Guangzhou, China).
4.5. Bioinformatic Analyses and DEGs Identification
Clean reads were obtained after removing the adaptor and filtering raw reads with more than 10% of unknown nucleotides and low-quality raw reads (Q-value ≤ 20) by fastp (version 0.18.0). Ribosome RNA (rRNA) was mapped to the ribosome RNA (rRNA) database and removed by using the short reads alignment tool Bowtie2 (version 2.2.8). Then paired-end clean reads were mapped to the reference genome (Accession number: PRJNA438564) using HISAT2. Gene expression levels were obtained by calculating the fragments per kilobase of exon model per million mapped reads (FPKM) value. The differentially expressed genes (DEGs) were analyzed by DESeq2 [47] software. Genes/transcripts with the parameter of false discovery rate (FDR) less than 0.05 and absolute log2 (fold change) ≥1 were considered differentially expressed genes/transcripts. The method for adjusting p-values was according to Benjamini and Hochberg [48].
4.6. AG Collection at Different Molting Stages
In order to detect the expression profiles of selected genes in AG of shrimp at different molting stages, AG tissues of five males were collected from each of the four different molting stages separately, including inter-molt stage (C), early/middle pre-molt stage (D0–2), late pre-molt stage (D3–4), and post-molt stage (P1–2) according to Gao et al. [49]. Extraction of the total RNA and synthesis of cDNA was performed on each sample according to 2.3.
4.7. 20-Hydroxyecdysone (20E) Injection
The pure powder of 20-hydroxyecdysone was bought from Aladdin Industrial Corporation (Shanghai, China) and dissolved in DMSO as a stock solution of 5 μg/μL. Before injection, the stock solution was dissolved with 1 × PBS into a suitable dosage. Animals at different molt stages were divided into four groups with 24 individuals in each group at one molting stage: inter-molt stage (C), early/middle pre-molt stage (D0–2), late pre-molt stage (D3–4), and post-molt stage (P1–2). In each group, eight individuals were injected with 1 μg 20E, 5 μg 20E, or equal volume PBS, respectively. The doses of injected 20E were used according to previous studies [50]. At 48 h post-injection, AG tissue from six individuals was collected from each group and used as three biological replicates. The samples were used for total RNA extraction and cDNA synthesis according to Section 4.3.
4.8. RT-qPCR Analysis
RT-qPCR was performed to detect the expression levels of Lv-IAG, hormone receptor 4 (Lv-Hr4), and three dopamine beta-hydroxylase genes (Lv-DBH1, Lv-DBH2, Lv-DBH3). The qPCR was performed on Eppendorf Mastercycler® ep realplex (Eppendorf, Germany) by using SuperReal PreMix Plus (SYBR Green, Toyobo, Japan) under the steps described below: denaturation at 94 °C for 1 min; 40 cycles of 94 °C for 20 s, annealing temperature for 20 s, and 72 °C for 20 s. As the internal reference gene, 18S rRNA was used. Primers were designed using Primer Premier 5 and are listed in Table 3. The specificity and accuracy of primers were determined by PCR assay and single peak in the melting curve of RT-qPCR assay.
4.9. Statistical Analysis
All real-time quantitative PCR experiments were performed with three replicates and normalized by using geometric averaging of the host gene 18S rRNA. The comparative Ct method was used with the formula 2−△△Ct to calculate the relative expression level of the candidate genes [46]. Data of real-time quantitative PCR were statistically tested by one-way ANOVA, and p-value < 0.05 denoted a statistically significant difference.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jmse9060677/s1, Figure S1: Expression levels (Ct values) of the internal gene 18S rRNA in all groups after 20E injection. Table S1: Differentially expressed genes identified from the transcriptome analysis.
Author Contributions
Conceptualization, S.L. and F.L.; methodology, validation, formal analysis and data curation, K.C. and S.L.; writing—original draft preparation, K.C. and S.L.; writing—review and editing, S.L., J.X., A.S. and F.L.; funding acquisition, S.L. and F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Joint NSFC-ISF Research Program (31861143047), the National Key R&D Program of China (2018YFD0900202), and the China Agriculture Research System of MOF and MARA.
Institutional Review Board Statement
This study used shrimp and experimental animals, which is not endangered invertebrates. In addition, there is no genetically modified organism used in the study. According to the national regulation (Fisheries Law of the People’s Republic of China), no permission is required to collect the animals and no formal ethics approval is required for this study.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sagi, A.; Aflalo, E.D. The androgenic gland and monosex culture of freshwater prawn Macrobrachium rosenbergii (De Man): A biotechnological perspective. Aquac. Res. 2005, 36, 231–237. [Google Scholar] [CrossRef]
- Laramore, S.; Laramore, C.R.; ScarpaEffect, J. Effect of low salinity on growth and survival of postlarvae and juvenile Litopenaeus vannamei. J. World Aquac. Soc. 2001, 32, 385–390. [Google Scholar] [CrossRef]
- Gitterle, T.; Rye, M.; Salte, R.; Cock, J.; Johansen, H.; Lozano, C.; Arturo Suárez, J.; Gjerde, B. Genetic (co)variation in harvest body weight and survival in Penaeus (Litopenaeus) vannamei under standard commercial conditions. Aquaculture 2005, 243, 83–92. [Google Scholar] [CrossRef]
- Levy, T.; Sagi, A. The “IAG-Switch”—A key controlling element in decapod crustacean sex differentiation. Front. Endocrinol. 2020, 11, 651. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing sexual differentiation: Full functional sex reversal achieved through silencing of a single insulin-like gene in the prawn, Macrobrachium rosenbergii. Biol. Reprod. 2012, 86, 90. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Manor, R.; Weil, S.; Gafni, O.; Linial, A.; Aflalo, E.D.; Ventura, T.; Sagi, A. A sexual shift induced by silencing of a single insulin-like gene in crayfish: Ovarian upregulation and testicular degeneration. PLoS ONE 2010, 5, e15281. [Google Scholar] [CrossRef] [PubMed]
- Manor, R.; Weil, S.; Oren, S.; Glazer, L.; Aflalo, E.D.; Ventura, T.; Chalifa-Caspi, V.; Lapidot, M.; Sagi, A. Insulin and gender: An insulin-like gene expressed exclusively in the androgenic gland of the male crayfish. Gen. Comp. Endocrinol. 2007, 150, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Sagi, A. The insulin-like androgenic gland hormone in crustaceans: From a single gene silencing to a wide array of sexual manipulation-based biotechnologies. Biotechnol. Adv. 2012, 30, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Khalaila, I.; Katz, T.; Abdu, U.; Yehezkel, G.; Sagi, A. Effects of implantation of hypertrophied androgenic glands on sexual characters and physiology of the reproductive system in the female red claw crayfish, Cherax quadricarinatus. Gen. Comp. Endocrinol. 2001, 121, 242–249. [Google Scholar] [CrossRef]
- Katakura, Y. Masculinization of females of the isopod crustacean, Armadillidium vulgare, following injections of an active extract of the androgenic gland. Gen. Comp. Endocrinol. 1983, 49, 57–62. [Google Scholar] [CrossRef]
- Yu, Y.-Q.; Ma, W.-M.; Zeng, Q.-G.; Qian, Y.-Q.; Yang, J.-S.; Yang, W.-J. Molecular cloning and sexually dimorphic expression of two dmrt genes in the giant freshwater prawn, Macrobrachium rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Zheng, J.; Cheng, S.; Jia, Y.; Gu, Z.; Li, F.; Chi, M.; Liu, S.; Jiang, W. Molecular identification and expression profiles of four splice variants of sex-lethal gene in Cherax quadricarinatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 234, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, F.; Yu, K.; Xiang, J. Identification and characterization of a doublesex gene which regulates the expression of insulin-like androgenic gland hormone in Fenneropenaeus chinensis. Gene 2018, 649, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Khalaila, I.; Manor, R.; Weil, S.; Granot, Y.; Keller, R.; Sagi, A. The eyestalk-androgenic gland-testis endocrine axis in the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol. 2002, 127, 147–156. [Google Scholar] [CrossRef]
- Foulks, N.B.; Hoffman, D.L. The effects of eyestalk ablation and B-ecdysone on RNA synthesis in the androgenic glands of the protandric shrimp, Pandalus platyceros brandt. Gen. Comp. Endocrinol. 1974, 22, 439–447. [Google Scholar] [CrossRef]
- Sroyraya, M.; Chotwiwatthanakun, C.; Stewart, M.J.; Soonklang, N.; Kornthong, N.; Phoungpetchara, I.; Hanna, P.J.; Sobhon, P. Bilateral eyestalk ablation of the blue swimmer crab, Portunus pelagicus, produces hypertrophy of the androgenic gland and an increase of cells producing insulin-like androgenic gland hormone. Tissue Cell 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Phoungpetchara, I.; Tinikul, Y.; Poljaroen, J.; Chotwiwatthanakun, C.; Vanichviriyakit, R.; Sroyraya, M.; Hanna, P.J.; Sobhon, P. Cells producing insulin-like androgenic gland hormone of the giant freshwater prawn, Macrobrachium rosenbergii, proliferate following bilateral eyestalk-ablation. Tissue Cell 2011, 43, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Grecia, V.-I.; Danitzia, A.G.-T.; Rodolfo, G.-T.; Píndaro, Á.-R.; Humberto, M.-R.; Rafael, C.-R. Quantitative analysis of hypertrophy and hyperactivity in the androgenic gland of eyestalk-ablated male Pacific white shrimp Litopenaeus vannamei during molt stages. Aquaculture 2015, 439, 7–13. [Google Scholar] [CrossRef]
- Chung, J.S.; Manor, R.; Sagi, A. Cloning of an insulin-like androgenic gland factor (IAG) from the blue crab, Callinectes sapidus: Implications for eyestalk regulation of IAG expression. Gen. Comp. Endocrinol. 2011, 173, 4–10. [Google Scholar] [CrossRef]
- Hopkins, P.M. The eyes have it: A brief history of crustacean neuroendocrinology. Gen. Comp. Endocrinol. 2012, 175, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Lachaise, F.; Le Roux, A.; Hubert, M.; Lafont, R. The molting gland of crustaceans: Localization, activity, and endocrine control (A review). J. Crustacean Biol. 1993, 13, 198–234. [Google Scholar] [CrossRef]
- Guo, Q.; Li, S.; Lv, X.; Xiang, J.; Manor, R.; Sagi, A.; Li, F. Sex-biased CHHs and their putative receptor regulate the expression of IAG gene in the shrimp Litopenaeus vannamei. Front. Physiol. 2019, 10, 1525. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, C.; Greve, P.; Martin, G. Overview on the sub-grouping of the crustacean hyperglycemic hormone family. Neuropeptides 1999, 33, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Liu, J.; Liu, F.; Huang, Y.; Wang, G.; Ye, H. Crustacean female sex hormone from the mud crab Scylla paramamosain is highly expressed in prepubertal males and inhibits the development of androgenic gland. Front. Physiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lu, B.; Wang, G.; Ye, H. Transcriptional Inhibition of Sp-IAG by Crustacean Female Sex Hormone in the Mud Crab, Scylla paramamosain. Int. J. Mol. Sci. 2020, 21, 5300. [Google Scholar] [CrossRef]
- Li, F.; Bai, H.; Zhang, W.; Fu, H.; Jiang, F.; Liang, G.; Jin, S.; Sun, S.; Qiao, H. Cloning of genomic sequences of three crustacean hyperglycemic hormone superfamily genes and elucidation of their roles of regulating insulin-like androgenic gland hormone gene. Gene 2015, 561, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Jun-Xia, C.; Guo-Li, Y.; Wei-Jun, Y. Identification of a novel male reproduction-related gene and its regulated expression patterns in the prawn, Macrobrachium rosenbergii. Peptides 2006, 27, 728–735. [Google Scholar] [CrossRef]
- Sroyraya, M.; Hanna, P.J.; Changklungmoa, N.; Senarai, T.; Siangcham, T.; Tinikul, Y.; Sobhon, P. Expression of the male reproduction-related gene in spermatic ducts of the blue swimming crab, Portunus pelagicus, and transfer of modified protein to the sperm acrosome. Microsc. Res. Tech. 2013, 76, 102–112. [Google Scholar] [CrossRef]
- Uchimura, T.; Hara, S.; Yazawa, T.; Kamei, Y.; Kitano, T. Involvement of heat shock proteins on the transcriptional regulation of corticotropin-releasing hormone in medaka. Front. Endocrinol. 2019, 10, 529. [Google Scholar] [CrossRef]
- Chandler, J.C.; Aizen, J.; Elizur, A.; Battaglene, S.C.; Ventura, T. Male sexual development and the androgenic gland: Novel insights through the de novo assembled transcriptome of the eastern spiny lobster, Sagmariasus verreauxi. Sex. Dev. 2015, 9, 338–354. [Google Scholar] [CrossRef]
- Fernandez, I.; Vijayakumar, P.; Marques, C.; Cancela, M.L.; Gavaia, P.J.; Laize, V. Zebrafish vitamin K epoxide reductases: Expression in vivo, along extracellular matrix mineralization and under phylloquinone and warfarin in vitro exposure. Fish. Physiol. Biochem. 2015, 41, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Beato, S.; Toledo-Solis, F.J.; Fernandez, I. Vitamin K in vertebrates’ reproduction: Further puzzling pieces of evidence from teleost fish species. Biomolecules 2020, 10, 1303. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, B.L.; Liu, B.Y.; Shi, S.; Gao, D.Y.; Zhang, T.C.; Shi, H.J.; Li, Z.; Shum, W.W. Vitamin K2-dependent GGCX and MGP are required for homeostatic calcium regulation of sperm maturation. iScience 2019, 14, 210–225. [Google Scholar] [CrossRef]
- Beckstead, R.B.; Lam, G.; Thummel, C.S. The genomic response to 20-hydroxyecdysone at the onset of Drosophila metamorphosis. Genome. Biol. 2005, 6, R99. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.Y.; Meng, Q.W.; Deng, P.; Guo, W.C.; Li, G.Q. Leptinotarsa hormone receptor 4 (HR4) tunes ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis. Insect Biochem. Mol. Biol. 2018, 94, 50–60. [Google Scholar] [CrossRef]
- Chan, S.-M.; Gu, P.-L.; Chu, K.-H.; Tobe, S.S. Crustacean neuropeptide genes of the CHH/MIH/GIH family: Implications from molecular studies. Gen. Comp. Endocrinol. 2003, 134, 214–219. [Google Scholar] [CrossRef]
- Chang, E.S.; Mykles, D.L. Regulation of crustacean molting: A review and our perspectives. Gen. Comp. Endocrinol. 2011, 172, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Webster, S.G.; Keller, R.; Dircksen, H. The CHH-superfamily of multifunctional peptide hormones controlling crustacean metabolism, osmoregulation, moulting, and reproduction. Gen. Comp. Endocrinol. 2012, 175, 217–233. [Google Scholar] [CrossRef]
- Techa, S.; Chung, J.S. Ecdysteroids regulate the levels of Molt-Inhibiting Hormone (MIH) expression in the blue crab, Callinectes sapidus. PLoS ONE 2015, 10, e0117278. [Google Scholar] [CrossRef]
- Raviv, S.; Parnes, S.; Sagi, A. Coordination of Reproduction and Molt in Decapods; Mente, E., Ed.; Reproductive Biology of Crustaceans Sciences Publishers: Enfield, NH, USA, 2008; pp. 365–390. [Google Scholar]
- Laufer, H.; Wainwright, G.; Young, N.J.; Sagi, A.; Ahl, J.S.B.; Rees, H.H. Ecdysteroids and juvenoids in two male morphotypes of Libinia emarginata. Insect Biochem. Mol. Biol. 1993, 23, 171–174. [Google Scholar] [CrossRef]
- Parnes, S.; Raviv, S.; Shechter, A.; Sagi, A. Males also have their time of the month! Cyclic disposal of old spermatophores, timed by the molt cycle, in a marine shrimp. J. Exp. Biol. 2006, 209, 4974–4983. [Google Scholar] [CrossRef]
- Dominik, M.-C.; Stephanie, A.W.; Klaus, H.H. Ecdysteroid levels in Daphnia magna during a molt cycle: Determination by radioimmunoassay (RIA) and liquid chromatography–mass spectrometry (LC–MS). Gen. Comp. Endocrinol. 2007, 151, 66–71. [Google Scholar] [CrossRef]
- Teruaki, N.; Haruyuki, S. Regulation of ecdysteroid secretion from the Y-organ by molt-inhibiting hormone in the American crayfish, Procambarus clarkii. Gen. Comp. Endocrinol. 2004, 135, 358–364. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, X.; Wei, J.; Sun, X.; Yuan, J.; Xiang, J. Whole transcriptome analysis provides insights into molecular mechanisms for molting in Litopenaeus vannamei. PLoS ONE 2015, 10, e0144350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.H.; Li, S.H.; Wang, Z.W.; Li, F.H.; Xiang, J.H. An eclosion hormone-like gene participates in the molting process of Palaemonid shrimp Exopalaemon carinicauda. Dev. Genes Evol. 2017, 227, 189–199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Principal component analysis (PCA) of the transcriptome, identification of differentially expressed genes (DEGs), and confirmation of Lv-IAG expression by RT-qPCR. (A) shows the PCA result of the ESA group (eAG1, eAG2, and eAG3) and the control group (cAG1, cAG2, and cAG3). The first two axes account for 15.4% (PC1) and 62.6% (PC2) of the variation. (B) shows the expression volcano plot of DEGs between the unilateral eyestalk ablation group and the control group. The abscissa represents the logarithm of the expression fold change of DEGs relative to control. The ordinate represents the negative logarithm of the statistical significance of the expression difference. The red dots represent up-regulated DEGs, the yellow dots represent down-regulated DEGs, and the blue dots represent non-DEGs. (C) shows the Lv-IAG expression profiles validated by RT-qPCR. Data are represented as means of fold change and standard deviations (n = 3). Statistical difference was determined using Student’s t-test (** p < 0.01).
Figure 1.
Principal component analysis (PCA) of the transcriptome, identification of differentially expressed genes (DEGs), and confirmation of Lv-IAG expression by RT-qPCR. (A) shows the PCA result of the ESA group (eAG1, eAG2, and eAG3) and the control group (cAG1, cAG2, and cAG3). The first two axes account for 15.4% (PC1) and 62.6% (PC2) of the variation. (B) shows the expression volcano plot of DEGs between the unilateral eyestalk ablation group and the control group. The abscissa represents the logarithm of the expression fold change of DEGs relative to control. The ordinate represents the negative logarithm of the statistical significance of the expression difference. The red dots represent up-regulated DEGs, the yellow dots represent down-regulated DEGs, and the blue dots represent non-DEGs. (C) shows the Lv-IAG expression profiles validated by RT-qPCR. Data are represented as means of fold change and standard deviations (n = 3). Statistical difference was determined using Student’s t-test (** p < 0.01).

Figure 2.
Expression profiles of Lv-IAG, Lv-Hr4, and three Lv-DBH genes in AG at different molting stages. Data are represented as means and standard deviations. Statistical differences were determined using Student’s t-test. The different letters indicate significant differences in gene expression level at the same molt stage (n = 3).
Figure 2.
Expression profiles of Lv-IAG, Lv-Hr4, and three Lv-DBH genes in AG at different molting stages. Data are represented as means and standard deviations. Statistical differences were determined using Student’s t-test. The different letters indicate significant differences in gene expression level at the same molt stage (n = 3).
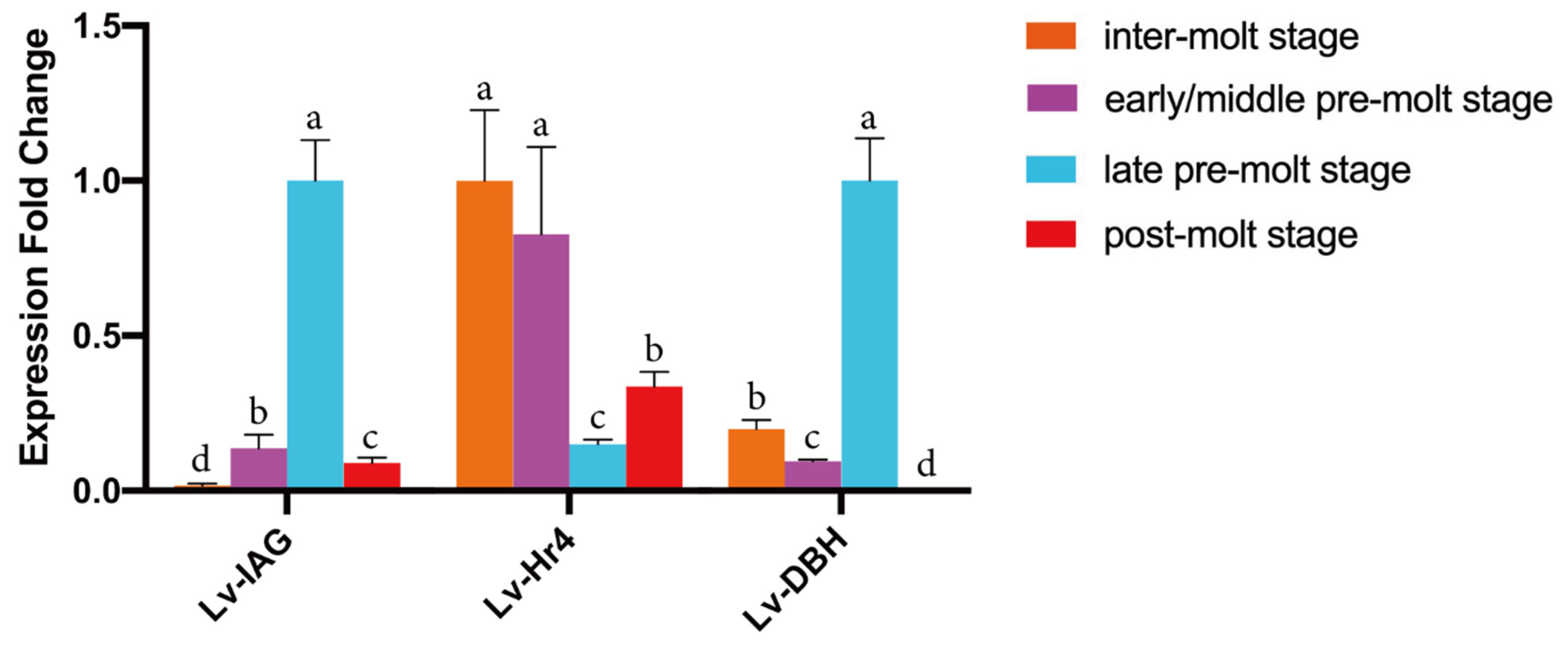
Figure 3.
The 20E injection regulated the expression of Lv-IAG (A) and Lv-Hr4 (B) throughout the molting cycle. Two dosages of 20E (1 μg and 5 μg per individual, respectively) were used, and an equal volume of PBS was injected as a negative control. Data are represented as means and standard deviations. Statistical differences were determined using Student’s t-test. The different letters indicate significant differences in the gene expression level at the same molt stage (n = 3).
Figure 3.
The 20E injection regulated the expression of Lv-IAG (A) and Lv-Hr4 (B) throughout the molting cycle. Two dosages of 20E (1 μg and 5 μg per individual, respectively) were used, and an equal volume of PBS was injected as a negative control. Data are represented as means and standard deviations. Statistical differences were determined using Student’s t-test. The different letters indicate significant differences in the gene expression level at the same molt stage (n = 3).

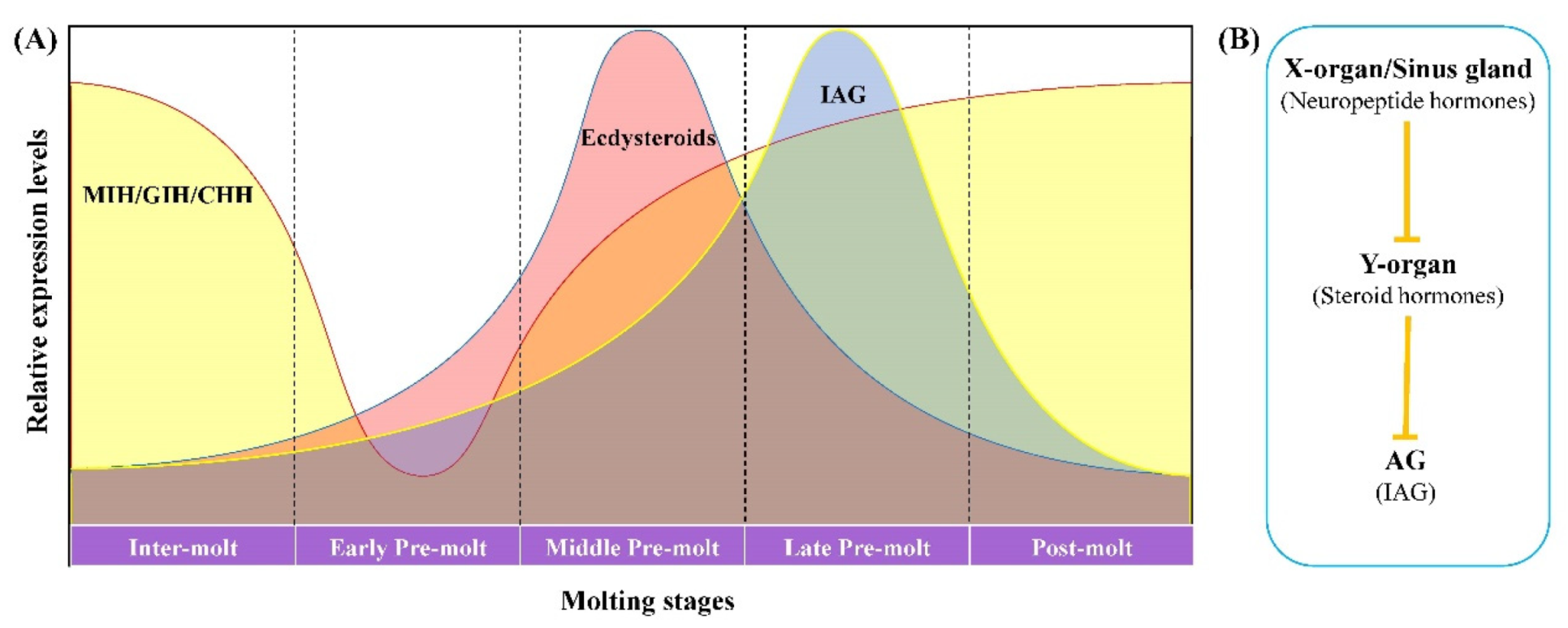
Figure 4.
The hypothesis of the regulatory mechanism between the secretion of neuropeptide hormones and the Lv-IAG expression level via the mediating role of ecdysteroids. (A) shows the expression profile of the inhibitory neuropeptides, ecdysteroids, and Lv-IAG throughout the molting cycle of male L. vannamei. (B) shows a diagram illustrating the hypothetical “X-organ/sinus gland–Y-organ–AG” endocrine axis. The biosynthesis of ecdysteroids was triggered by the decreased secretion of neuropeptide hormones from X-organ/sinus gland complex at the early pre-molt stage. When the concentration of ecdysteroids decreased dramatically at the late pre-molt stage, the suppression effect of ecdysteroids on Lv-IAG expression was eliminated.
Figure 4.
The hypothesis of the regulatory mechanism between the secretion of neuropeptide hormones and the Lv-IAG expression level via the mediating role of ecdysteroids. (A) shows the expression profile of the inhibitory neuropeptides, ecdysteroids, and Lv-IAG throughout the molting cycle of male L. vannamei. (B) shows a diagram illustrating the hypothetical “X-organ/sinus gland–Y-organ–AG” endocrine axis. The biosynthesis of ecdysteroids was triggered by the decreased secretion of neuropeptide hormones from X-organ/sinus gland complex at the early pre-molt stage. When the concentration of ecdysteroids decreased dramatically at the late pre-molt stage, the suppression effect of ecdysteroids on Lv-IAG expression was eliminated.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the sequencing and mapping output of all samples.
| Sample | Raw Reads | Clean Reads | Q20 (%) | Q30 (%) | Total Mapped | GC Content (%) |
|---|---|---|---|---|---|---|
| eAG1 | 39,144,350 | 38,623,336 | 97.64 | 93.61 | 32,983,645 (88.06%) | 48.65 |
| eAG2 | 38,000,328 | 37,384,452 | 97.48 | 93.28 | 31,424,401 (87.06%) | 47.65 |
| eAG3 | 37,794,382 | 37,206,858 | 97.53 | 93.47 | 32,130,046 (88.44%) | 48.77 |
| cAG1 | 42,634,574 | 41,927,468 | 97.50 | 93.34 | 36,357,115 (88.87%) | 49.85 |
| cAG2 | 38,989,158 | 38,455,262 | 97.74 | 93.84 | 31,867,585 (86.36%) | 49.13 |
| cAG3 | 44,795,738 | 43,610,558 | 97.53 | 93.63 | 37,288,555 (88.04%) | 50.29 |
Table 2.
Identified DEGs related to sexual development and endocrinology.
| Categories | Gene Symbol | Accession Number | Annotation | Log2 (Fold Change ESA/Control) |
|---|---|---|---|---|
| Sexual development | Lv-IAG | MSTRG.32535 | insulin-like androgenic gland hormone | 2.41 |
| Lv-Mrr1 | ROT78373.1 | male reproductive-related protein A | 3.09 | |
| Lv-Mrr2 | MSTRG.11644 | male reproductive-related protein A | 3.3 | |
| Lv-GGCX | ROT82257.1 | Vitamin K-dependent gamma-carboxylase | 3.3 | |
| Lv-Calu | ROT83784.1 | Calumenin | 2.76 | |
| Lv-Hsp90-1 | ROT62469.1 | heat shock protein 90 | 2.76 | |
| Lv-Hsp70 | ROT68844.1 | heat shock protein 70 | 3.1 | |
| Lv-Hsp90-2 | ROT76137.1 | heat shock protein 90 | 2.63 | |
| Lv-ZSCAN31 | ROT76330.1 | zinc finger and SCAN domain-containing protein 31 | −4.18 | |
| Lv-ZCWPW1 | MSTRG.12712 | zinc finger/CW type with PWWP domain 1 | −1.89 | |
| Endocrine system | Lv-Hr4 | ROT82234.1 | hormone receptor 4 | −1.75 |
| Lv-SPI | ROT62953.1 | serine proteinase inhibitor | 2.47 | |
| Lv-SPI3 | ROT75925.1 | serine proteinase inhibitor-3 | −6.43 | |
| Lv-DBH1 | ROT77266.1 | dopamine beta-hydroxylase | 2.35 | |
| Lv-DBH2 | ROT77268.1 | dopamine beta-hydroxylase | 2.75 | |
| Lv-DBH3 | ROT77267.1 | dopamine beta-hydroxylase | 2.54 | |
| Lv-GRWD1 | ROT82204.1 | glutamate-rich WD repeat-containing protein 1 | 2.47 | |
| Lv-LRP2 | ROT73017.1 | lipoprotein receptor-related protein 2 | −4.25 | |
| Lv-PA2G4 | ROT64438.1 | proliferation-associated protein 2G4 | 1.79 | |
| Lv-ANK3 | ROT77030.1 | ankyrin-3-like isoform X1 | −3.05 |
Table 3.
All primers used in this study.
| Gene Symbol | Primer Name | Nucleotide Sequences (5′-3′) | Annealing Temperature (°C) |
|---|---|---|---|
| Lv-IAG | Lv-IAG-qF | AGTGTCAAGGTCAGCCGATAC | 54.5 °C |
| Lv-IAG-qR | CGAGATTCCACGTTGGATTCAG | ||
| Lv-18S | 18S-F | TATACGCTAGTGGAGCTGGAA | 54.9 °C |
| 18S-R | GGGGAGGTAGTGACGAAAAAT | ||
| Lv-Hr4 | Lv-Hr4-F | AGACTCCCCAGCAGCTATGA | 55.0 °C |
| Lv-Hr4-R | GTTGCGCTGTGCCTTTGTAA | ||
| Lv-DBH1 | Lv-DBH1-qF | TCGAAGCCGAAGACAGAACC | 56.0 °C |
| Lv-DBH1-qR | AGGCGCTCACTTCTCTCAAC | ||
| Lv-DBH2 | Lv-DBH2-qF | CAGCACCGATTTCAGCGATG | 55.0 °C |
| Lv-DBH2-qR | ATCCTCTTCCCCTGGAGTCC | ||
| Lv-DBH3 | Lv-DBH3-qF | TATGGTGCCAACATGCCCAT | 54.0 °C |
| Lv-DBH3-qR | GGGACGATTTGGATTGGGGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, K.; Li, S.; Xiang, J.; Sagi, A.; Li, F. Transcriptome Analysis Reveals the Endocrine Regulation on the Expression of IAG in Litopenaeus vannamei. J. Mar. Sci. Eng. 2021, 9, 677. https://doi.org/10.3390/jmse9060677
AMA Style
Chen K, Li S, Xiang J, Sagi A, Li F. Transcriptome Analysis Reveals the Endocrine Regulation on the Expression of IAG in Litopenaeus vannamei. Journal of Marine Science and Engineering. 2021; 9(6):677. https://doi.org/10.3390/jmse9060677
Chicago/Turabian StyleChen, Kangxuan, Shihao Li, Jianhai Xiang, Amir Sagi, and Fuhua Li. 2021. "Transcriptome Analysis Reveals the Endocrine Regulation on the Expression of IAG in Litopenaeus vannamei" Journal of Marine Science and Engineering 9, no. 6: 677. https://doi.org/10.3390/jmse9060677
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.