
Review on Gilthead Seabream (Sparus aurata) Aquaculture: Life Cycle, Growth, Aquaculture Practices and Challenges

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Gilthead Seabream’s Taxonomy and Habitat
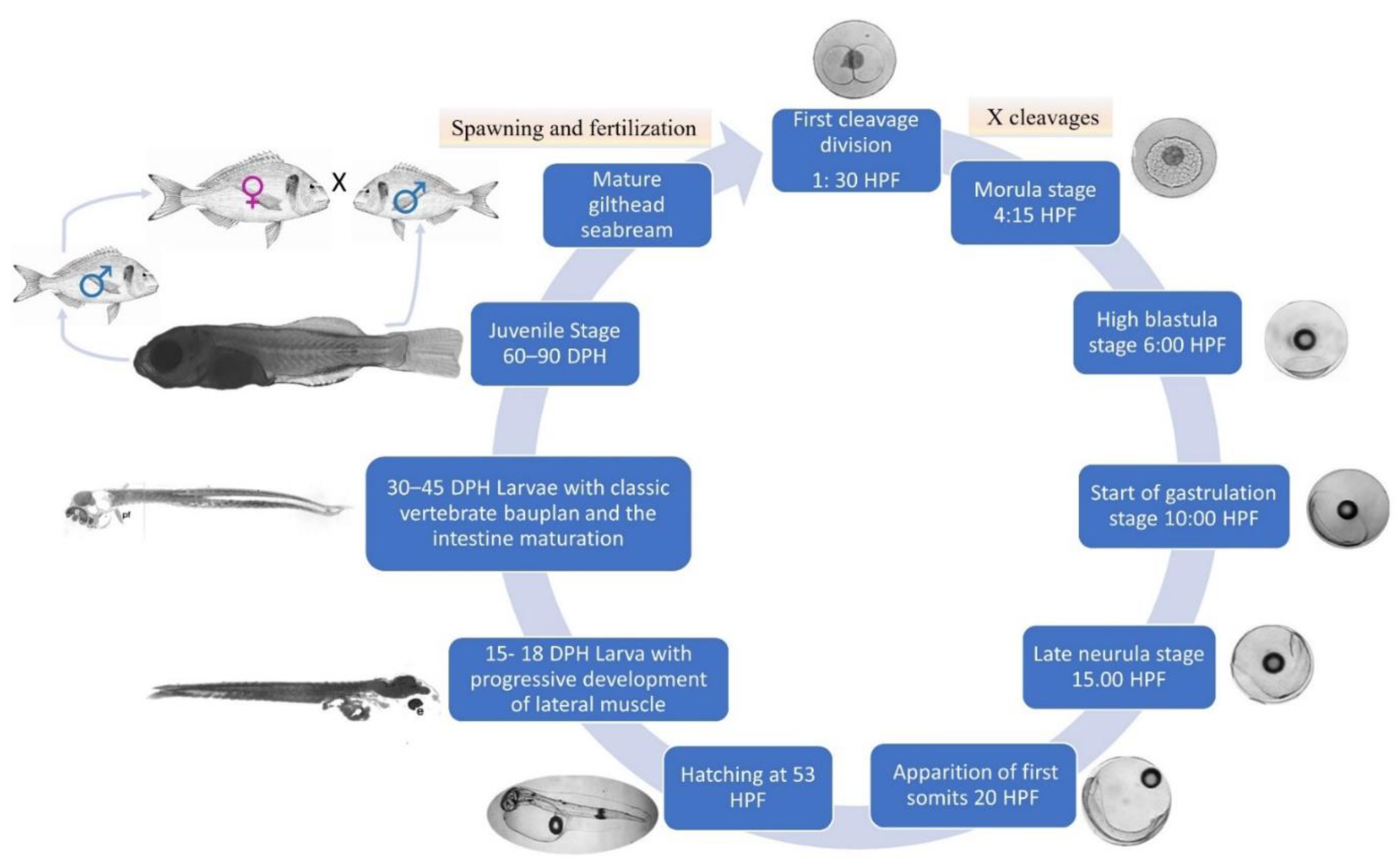
3. Gilthead Seabream’ Developmental Stages

4. Growth Characteristics and Requirements

{kind=link}
{kind=link}
{kind=link}
| Mongile et al., 2014 [54] | Fountoulaki et al., 2005 [55] | Pelusio et al., 2021 [56] | Basto-Silva et al., 2022 [59] | |||
|---|---|---|---|---|---|---|
| Average weight (g) | 75 | 2.1 | 2.8 | 67.50 ± 1.66 | 9.10 ± 0.01 | |
| Experimental condition | 27 ± 1 °C Recirculating flow of 16.6 L/min Water renewal: 5% daily Photoperiod: 12/12 DL Light intensity: 200 lx Dissolved oxygen: 100% saturation ad libitum | Open flow Feeding to satiation | Closed recirculating aquaculture system Oxygen level: 8.0 ± 1.0 mg/L Total ammonia nitrogen ≤ 0.1 mg L–1, NO2 ≤ 0.2 mg/L NO3 ≤ 50 mg/L Salinity: 25–30 g/L | 24 ± 1 °C Continuous flow: 6 L/min Dissolved oxygen: near saturation | ||
| 25 °C | 10–14 °C | 23.17 ± 1.11 °C to 17.34 ± 0.92 °C | 17 °C to 23 °C | |||
| Protein | 46% | 51% | 46% | 40 | ||
| Lipids | 18% | 16–21% | 21% | 16% | - | |
| Carbohydrate | - | - | - | 20 | ||
| Energy content of the feed | - | 21 | 20 | - | ||
| Manganese [62] | Zinc [61] | Selenium [61] | Cobalt [61] | Calcium [61] | |
|---|---|---|---|---|---|
| Average weight (g) | 12.6 ± 1.5 | 25.5 ± 2.7 | |||
| Experimental conditions | Temperature: 19.4 ± 0.4 °C Photoperiod: 12 h L/12 h D Diets high in vegetable ingredients (fish meal: 10%; fish oil: 6%) | Temperature: 19.4 ± 0.4 °C Photoperiod: 12 h L/12 h D Diets with low FM and FO levels | |||
| Requirements | 19 | 150 mg/kg | 0.77 mg/kg | 1.9 mg/kg | ≤7.6 g/kg |
5. Genetic Insights into Gilthead Seabream Populations
6. Breeding and Culture Practices
6.1. History
6.2. Production and Trade
6.3. Gilthead Seabream Farming from Hatchery to Harvest
7. Aquaculture Challenges
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Meaning |
| DPH | Day post hatching |
| HPF | Hour post fertilization |
| HUFAs | Highly unsaturated fatty acids |
| SGR | Specific growth rate |
| Se | Selenium |
| Zn | Zinc |
| Mn | Manganese |
| Ca | Calcium |
| SNP | Single-nucleotide polymorphism |
| VER | Viral encephalopathy and retinopathy |
| VNN | Viral nervous necrosis |
| EAFP | European Association of Fish Pathologists |
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Aragona, M.; Mhalhel, K.; Montalbano, G.; Germanà, A. Morphological Characteristics of the Blackspot Seabream (Pagellus bogaraveo) Tongue: A Structural and Immunohistochemical Study. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2022, 51, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary Inclusion of Tenebrio Molitor Larvae Meal: Effects on Growth Performance and Final Quality Treats of Blackspot Sea Bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Aragona, M.; Mhalhel, K.; Montalbano, G.; Germanà, A. Anatomical, Histological and Immunohistochemical Study of the Tongue in the Rainbow Trout (Oncorhynchus mykiss). Anat. Histol. Embryol. 2020, 49, 848–858. [Google Scholar] [CrossRef]
- D’Agaro, E.; Gibertoni, P.; Esposito, S. Recent Trends and Economic Aspects in the Rainbow Trout (Oncorhynchus mykiss) Sector. Appl. Sci. 2022, 12, 8773. [Google Scholar] [CrossRef]
- Guerrera, M.C.; Aragona, M.; Briglia, M.; Porcino, C.; Mhalhel, K.; Cometa, M.; Abbate, F.; Montalbano, G.; Laurà, R.; Levanti, M.; et al. The Alimentary Tract of African Bony-Tongue, Heterotis Niloticus (Cuvier, 1829): Morphology Study. Animals 2022, 12, 1565. [Google Scholar] [CrossRef]
- Monentcham, S.-E.; Kouam, J.; Pouomogne, V.; Kestemont, P. Biology and Prospect for Aquaculture of African Bonytongue, Heterotis Niloticus (Cuvier, 1829): A Review. Aquaculture 2009, 289, 191–198. [Google Scholar] [CrossRef]
- NFISS. FishStatJ-Software for Fishery and Aquaculture Statistical Time Series. 2022. Available online: https://www.fao.org/fishery/en/topic/166235?lang=en (accessed on 5 August 2023).
- Dellacqua, Z.; Di Biagio, C.; Costa, C.; Pousão-Ferreira, P.; Ribeiro, L.; Barata, M.; Gavaia, P.J.; Mattei, F.; Fabris, A.; Izquierdo, M.; et al. Distinguishing the Effects of Water Volumes versus Stocking Densities on the Skeletal Quality during the Pre-Ongrowing Phase of Gilthead Seabream (Sparus aurata). Animals 2023, 13, 557. [Google Scholar] [CrossRef]
- Espírito-Santo, C.; Guardiola, F.A.; Ozório, R.O.A.; Magnoni, L.J. Short-Term Swimming up-Regulates pro-Inflammatory Mediators and Cytokines in Gilthead Seabream (Sparus aurata). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2023, 284, 111487. [Google Scholar] [CrossRef] [PubMed]
- Mhalhel, K.; Germanà, A.; Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Montalbano, G. The Effect of Orally Supplemented Melatonin on Larval Performance and Skeletal Deformities in Farmed Gilthead Seabream (Sparus aurata). Int. J. Mol. Sci. 2020, 21, 9597. [Google Scholar] [CrossRef]
- Mhalhel, K.; Montalbano, G.; Giurdanella, G.; Abbate, F.; Laurà, R.; Guerrera, M.C.; Germanà, A.; Levanti, M. Histological and Immunohistochemical Study of Gilthead Seabream Tongue from the Early Stage of Development: TRPV4 Potential Roles. Ann. Anat. 2022, 244, 151985. [Google Scholar] [CrossRef]
- Sola, L.; Moretti, A.; Crosetti, D.; Karaiskou, N.; Magoulas, A.; Rossi, A.R.; Rye, M.; Triantafyllidis, A.; Tsigenopoulos, C.S. Gilthead Seabream—Sparus aurata. Genetic Impact of Aquaculture Activities on Native Populations. 2007, pp. 47–54. Available online: https://www.vliz.be/imisdocs/publications/318461.pdf (accessed on 2 July 2023).
- Verhaegen, Y.; Adriaens, D.; Wolf, T.D.; Dhert, P.; Sorgeloos, P. Deformities in Larval Gilthead Sea Bream (Sparus aurata): A Qualitative and Quantitative Analysis Using Geometric Morphometrics. Aquaculture 2007, 268, 156–168. [Google Scholar] [CrossRef]
- Sadek, S.; Osman, M.F.; Mansour, M.A. Growth, Survival and Feed Conversion Rates of Sea Bream (Sparus aurata) Cultured in Earthen Brackish Water Ponds Fed Different Feed Types. Aquac. Int. 2004, 12, 409–421. [Google Scholar] [CrossRef]
- Kır, M. Thermal Tolerance and Standard Metabolic Rate of Juvenile Gilthead Seabream (Sparus aurata) Acclimated to Four Temperatures. J. Therm. Biol. 2020, 93, 102739. [Google Scholar] [CrossRef]
- Manchado, M.; Planas, J.V.; Cousin, X.; Rebordinos, L.; Claros, M.G. 8-Current Status in Other Finfish Species: Description of Current Genomic Resources for the Gilthead Seabream (Sparus aurata) and Soles (Solea senegalensis and Solea solea). In Genomics in Aquaculture; MacKenzie, S., Jentoft, S., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 195–221. ISBN 978-0-12-801418-9. [Google Scholar]
- Rowlerson, A.; Mascarello, F.; Radaelli, G.; Veggetti, A. Differentiation and Growth of Muscle in the Fish Sparus aurata (L.): II. Hyperplastic and Hypertrophic Growth of Lateral Muscle from Hatching to Adult. J. Muscle Res. Cell Motil. 1995, 16, 223–236. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Animal Welfare Aspects of Husbandry Systems for Farmed European Seabass and Gilthead Seabream-Scientific Opinion of the Panel. EFSA J. 2008, 6, 844. [Google Scholar] [CrossRef]
- Ortiz-Delgado, J.B.; Fernández, I.; Sarasquete, C.; Gisbert, E. Normal and Histopathological Organization of the Opercular Bone and Vertebrae in Gilthead Sea Bream Sparus aurata. Aquat. Biol. 2014, 21, 67–84. [Google Scholar] [CrossRef]
- Sfakianakis, D.G.; Katharios, P.; Tsirigotakis, N.; Doxa, C.K.; Kentouri, M. Lateral Line Deformities in Wild and Farmed Sea Bass (Dicentrarchus labrax, L.) and Sea Bream (Sparus aurata, L.). J. Appl. Ichthyol. 2013, 29, 1015–1021. [Google Scholar] [CrossRef]
- Beraldo, P.; Canavese, B. Recovery of Opercular Anomalies in Gilthead Sea Bream, Sparus aurata L.: Morphological and Morphometric Analysis. J. Fish Dis. 2011, 34, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Negrín-Báez, D.; Navarro, A.; Lee-Montero, I.; Soula, M.; Afonso, J.M.; Zamorano, M.J. Inheritance of Skeletal Deformities in Gilthead Seabream (Sparus aurata)-Lack of Operculum, Lordosis, Vertebral Fusion and LSK Complex. J. Anim. Sci. 2015, 93, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Pavlidis, M.A.; Mylonas, C.C. Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; ISBN 1-4051-9772-2. [Google Scholar]
- FAO. Aquaculture Feed and Fertilizer Resources Information System: Gilthead Seabream-Sparus aurata. Available online: https://www.fao.org/fishery/affris/species-profiles/gilthead-seabream/gilthead-seabream-home/en/ (accessed on 5 August 2023).
- Pita, C.; Gamito, S.; Erzini, K. Feeding Habits of the Gilthead Seabream (Sparus aurata) from the Ria Formosa (Southern portugal) as Compared to the Black Seabream (Spondyliosoma cantharus) and the Annular Seabream (Diplodus annularis). J. Appl. Ichthyol. 2002, 18, 81–86. [Google Scholar] [CrossRef]
- Wassef, E.; Eisawy, A. Food and Feeding Habits of Wild and Reared Gilthead Bream Sparus aurata L. Cybium 1985, 9, 233–242. [Google Scholar]
- The European Commission. AquaTrace Species Leaflet: Gilthead Sea Bream (Sparus aurata). 2014. Available online: https://arquivo.pt/wayback/20141123083703/https://aquatrace.eu/ (accessed on 12 August 2023).
- Kamacı, H.O.; Saka, Ş.; Fırat, K. The Cleavage and Embryonic Phase of Gilthead Sea Bream (Sparus aurata Linnaeus, 1758) Eggs. Su Ürünleri Derg. 2005, 22, 205–209. [Google Scholar]
- Desoutter, M.; Quero, J.C.; Hureau, J.C.; Karrer, C.; Post, A.; Saldanha, L. Check-List of the Fishes of the Eastern Tropical Atlantic (CLOFETA); UNESCO: Paris, France, 1990; ISBN 92-3-002620-4. [Google Scholar]
- Patruno, M.; Radaelli, G.; Mascarello, F.; Candia Carnevali, M.D. Muscle Growth in Response to Changing Demands of Functions in the Teleost Sparus aurata (L.) during Development from Hatching to Juvenile. Anat. Embryol. 1998, 198, 487–504. [Google Scholar] [CrossRef] [PubMed]
- Yúfera, M.; Halm, S.; Beltran, S.; Fusté, B.; Planas, J.V.; Martínez-Rodríguez, G. Transcriptomic Characterization of the Larval Stage in Gilthead Seabream (Sparus aurata) by 454 Pyrosequencing. Mar. Biotechnol. 2012, 14, 423–435. [Google Scholar] [CrossRef]
- Colloca, F.; Cerasi, S. Sparus aurata. Cultured Aquatic Species Information Programme; FAO: Rome, Italy, 2022. [Google Scholar]
- Mehanna, S.F. A Preliminary Assessment and Management of Gilthead Bream Sparus aurata in the Port Said Fishery, the Southeastern Mediterranean, Egypt. Turk. J. Fish. Aquat. Sci. 2007, 7, 123–130. [Google Scholar]
- Chaoui, L.; Kara, M.H.; Faure, E.; Quignard, J.P. Growth and Reproduction of the Gilthead Seabream Sparus aurata in Mellah Lagoon (North-Eastern Algeria). Sci. Mar. 2006, 70, 545–552. [Google Scholar] [CrossRef]
- Jerez, S.; Rodríguez, C.; Cejas, J.R.; Martín, M.V.; Bolaños, A.; Lorenzo, A. Influence of Age of Female Gilthead Seabream (Sparus aurata L.) Broodstock on Spawning Quality throughout the Reproductive Season. Aquaculture 2012, 350–353, 54–62. [Google Scholar] [CrossRef]
- Fernández-Palacios, H.; Izquierdo, M.S.; Robaina, L.; Valencia, A.; Salhi, M.; Vergara, J. Effect of n − 3 HUFA Level in Broodstock Diets on Egg Quality of Gilthead Sea Bream (Sparus aurata L.). Aquaculture 1995, 132, 325–337. [Google Scholar] [CrossRef]
- Zohar, Y.; Abraham, M.; Gordin, H. The Gonadal Cycle of the Captivity-Reared Hermaphroditic Teleost Sparus aurata (L.) during the First Two Years of Life. Ann. Biol. Anim. Biochim. Biophys. 1978, 18, 877–882. [Google Scholar] [CrossRef]
- Pauletto, M.; Manousaki, T.; Ferraresso, S.; Babbucci, M.; Tsakogiannis, A.; Louro, B.; Vitulo, N.; Quoc, V.H.; Carraro, R.; Bertotto, D.; et al. Genomic Analysis of Sparus aurata Reveals the Evolutionary Dynamics of Sex-Biased Genes in a Sequential Hermaphrodite Fish. Commun. Biol. 2018, 1, 119. [Google Scholar] [CrossRef]
- Tsakogiannis, A.; Manousaki, T.; Lagnel, J.; Papanikolaou, N.; Papandroulakis, N.; Mylonas, C.C.; Tsigenopoulos, C.S. The Gene Toolkit Implicated in Functional Sex in Sparidae Hermaphrodites: Inferences from Comparative Transcriptomics. Front. Genet. 2019, 9, 749. [Google Scholar] [CrossRef]
- Laiz-Carrión, R.; Sangiao-Alvarellos, S.; Guzmán, J.M.; Martín del Río, M.P.; Soengas, J.L.; Mancera, J.M. Growth Performance of Gilthead Sea Bream Sparus aurata in Different Osmotic Conditions: Implications for Osmoregulation and Energy Metabolism. Aquaculture 2005, 250, 849–861. [Google Scholar] [CrossRef]
- Torno, C.; Staats, S.; Fickler, A.; de Pascual-Teresa, S.; Soledad Izquierdo, M.; Rimbach, G.; Schulz, C. Combined Effects of Nutritional, Biochemical and Environmental Stimuli on Growth Performance and Fatty Acid Composition of Gilthead Sea Bream (Sparus aurata). PLoS ONE 2019, 14, e0216611. [Google Scholar] [CrossRef]
- Ayala, M.D.; Balsalobre, N.; Chaves-Pozo, E.; Sáez, M.I.; Galafat, A.; Alarcón, F.J.; Martínez, T.F.; Arizcun, M. Long-Term Effects of a Short Juvenile Feeding Period with Diets Enriched with the Microalgae Nannochloropsis Gaditana on the Subsequent Body and Muscle Growth of Gilthead Seabream, Sparus aurata L. Animals 2023, 13, 482. [Google Scholar] [CrossRef] [PubMed]
- Salmerón, C.; Navarro, I.; Johnston, I.A.; Gutiérrez, J.; Capilla, E. Characterisation and Expression Analysis of Cathepsins and Ubiquitin-Proteasome Genes in Gilthead Sea Bream (Sparus aurata) Skeletal Muscle. BMC Res. Notes 2015, 8, 149. [Google Scholar] [CrossRef]
- García de la Serrana, D.; Codina, M.; Capilla, E.; Jiménez-Amilburu, V.; Navarro, I.; Du, S.-J.; Johnston, I.A.; Gutiérrez, J. Characterisation and Expression of Myogenesis Regulatory Factors during in Vitro Myoblast Development and in Vivo Fasting in the Gilthead Sea Bream (Sparus aurata). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 167, 90–99. [Google Scholar] [CrossRef]
- Kraljević, M.; Dulčić, J. Age and Growth of Gilt-Head Sea Bream (Sparus aurata L.) in the Mirna Estuary, Northern Adriatic. Fish. Res. 1997, 31, 249–255. [Google Scholar] [CrossRef]
- Aydin, M. Maximum Length and Age Report of Sparus aurata (Linnaeus, 1758) in the Black Sea. J. Appl. Ichthyol. 2018, 34, 964–966. [Google Scholar] [CrossRef]
- Caterina, F.; Piccione, G.; Marafioti, S.; Arfuso, F.; Fortino, G.; Fazio, F. Metabolic Response to Monthly Variations of Sparus aurata Reared in Mediterranean On-Shore Tanks. Turk. J. Fish. Aquat. Sci. 2014, 14, 567–574. [Google Scholar] [CrossRef]
- Ginés, R.; Afonso, J.M.; Argüello, A.; Zamorano, M.J.; López, J.L. The Effects of Long-Day Photoperiod on Growth, Body Composition and Skin Colour in Immature Gilthead Sea Bream (Sparus aurata L.). Aquac. Res. 2004, 35, 1207–1212. [Google Scholar] [CrossRef]
- Vardar, H.; Yildirim, Ş. Effects of Long-Term Extended Photoperiod on Somatic Growth and Husbandry Parameters on Cultured Gilthead Seabream (Sparus aurata, L.) in the Net Cages. Turk. J. Fish. Aquat. Sci. 2012, 12, 225–231. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bortoletti, M.; Olivotto, I.; Ratti, S.; Poltronieri, C.; Negrato, E.; Caberlotto, S.; Radaelli, G.; Bertotto, D. Salinity, Temperature and Ammonia Acute Stress Response in Seabream (Sparus aurata) Juveniles: A Multidisciplinary Study. Animals 2021, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, H.; Nour, A. al-Aziz Effect of Reducing Water Salinity on Survival, Growth Performance, Chemical Composition and Nutrients Gain of Gilthead Sea Bream (Sparus aurata) Larvae. J. King Abdulaziz Univ. Mar. Sci. 2011, 22, 15–29. [Google Scholar]
- Lupatsch, I.; Kissil, G.; Sklan, D. Defining Energy and Protein Requirements Of Gilthead Seabream (Sparus aurata) To Optimize Feeds And Feeding Regimes. Isr. J. Aquac. Bamidgeh 2003, 55, 243–257. [Google Scholar] [CrossRef]
- Mongile, F.; Bonaldo, A.; Fontanillas, R.; Mariani, L.; Badiani, A.; Bonvini, E.; Parma, L. Effects of Dietary Lipid Level on Growth and Feed Utilisation of Gilthead Seabream (Sparus aurata L.) Reared at Mediterranean Summer Temperature. Ital. J. Anim. Sci. 2014, 13, 2999. [Google Scholar] [CrossRef]
- Fountoulaki, E.E.; Alexis, M.N.; Nengas, I. Protein and Energy Requirements of Gilthead Bream (Sparus aurata, L.) Fingerlings: Preliminary Results. In Mediterranean Fish Nutrition; Basurco, B., Izquierdo, M., Montero, D., Nengas, I., Alexis, M., Eds.; Cahiers Options Méditerranéennes; CIHEAM: Zaragoza, Spain, 2005; Volume 63, pp. 19–26. [Google Scholar]
- Pelusio, N.F.; Scicchitano, D.; Parma, L.; Dondi, F.; Brini, E.; D’Amico, F.; Candela, M.; Yúfera, M.; Gilannejad, N.; Moyano, F.J. Interaction between Dietary Lipid Level and Seasonal Temperature Changes in Gilthead Sea Bream Sparus aurata: Effects on Growth, Fat Deposition, Plasma Biochemistry, Digestive Enzyme Activity, and Gut Bacterial Community. Front. Mar. Sci. 2021, 8, 664701. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Dietary Carbohydrate Utilization by European Sea Bass (Dicentrarchus labrax L.) and Gilthead Sea Bream (Sparus aurata L.) Juveniles. Rev. Fish. Sci. 2011, 19, 201–215. [Google Scholar] [CrossRef]
- Caballero, M.J.; López-Calero, G.; Socorro, J.; Roo, F.J.; Izquierdo, M.S.; Férnandez, A.J. Combined Effect of Lipid Level and Fish Meal Quality on Liver Histology of Gilthead Seabream (Sparus aurata). Aquaculture 1999, 179, 277–290. [Google Scholar] [CrossRef]
- Basto-Silva, C.; Enes, P.; Oliva-Teles, A.; Capilla, E.; Guerreiro, I. Dietary Protein/Carbohydrate Ratio and Feeding Frequency Affect Feed Utilization, Intermediary Metabolism, and Economic Efficiency of Gilthead Seabream (Sparus aurata) Juveniles. Aquaculture 2022, 554, 738182. [Google Scholar] [CrossRef]
- Atalah, E.; Hernández-Cruz, C.M.; Ganga, R.; Ganuza, E.; Benítez-Santana, T.; Roo, J.; Fernández-Palacios, H.; Izquierdo, M.S. Enhancement of Gilthead Seabream (Sparus aurata) Larval Growth by Dietary Vitamin E in Relation to Two Different Levels of Essential Fatty Acids. Aquac. Res. 2012, 43, 1816–1827. [Google Scholar] [CrossRef]
- Domínguez, D.; Montero, D.; Robaina, L.; Hamre, K.; Terova, G.; Karalazos, V.; Izquierdo, M. Effects of Graded Levels of Minerals in a Multi-Nutrient Package on Gilthead Sea Bream (Sparus aurata) Fed a Plant-Based Diet. Aquac. Nutr. 2020, 26, 1007–1018. [Google Scholar] [CrossRef]
- Dominguez, D.; Sehnine, Z.; Castro, P.; Zamorano, M.J.; Robaina, L.; Fontanillas, R.; Antony Jesu Prabhu, P.; Izquierdo, M. Dietary Manganese Levels for Gilthead Sea Bream (Sparus aurata) Fingerlings Fed Diets High in Plant Ingredients. Aquaculture 2020, 529, 735614. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Ghrab, W.; Roo, J.; Hamre, K.; Hernández-Cruz, C.M.; Bernardini, G.; Terova, G.; Saleh, R. Organic, Inorganic and Nanoparticles of Se, Zn and Mn in Early Weaning Diets for Gilthead Seabream (Sparus aurata; Linnaeus, 1758). Aquac. Res. 2017, 48, 2852–2867. [Google Scholar] [CrossRef]
- Domínguez, D.; Robaina, L.; Zamorano, M.J.; Karalazos, V.; Izquierdo, M. Effects of Zinc and Manganese Sources on Gilthead Seabream (Sparus aurata) Fingerlings. Aquaculture 2019, 505, 386–392. [Google Scholar] [CrossRef]
- Domínguez, D.; Rimoldi, S.; Robaina, L.E.; Torrecillas, S.; Terova, G.; Zamorano, M.J.; Karalazos, V.; Hamre, K.; Izquierdo, M. Inorganic, Organic, and Encapsulated Minerals in Vegetable Meal Based Diets for Sparus aurata (Linnaeus, 1758). PeerJ 2017, 5, e3710. [Google Scholar] [CrossRef]
- Hossain, M.A.; Yoshimatsu, T. Dietary Calcium Requirement in Fishes. Aquac. Nutr. 2014, 20, 12135. [Google Scholar] [CrossRef]
- Korkut, Y.K.A.Y.; Gurkan, S. Length-Weight Relationship and Condition Factor as an Indicator of Growth and Feeding Intensity of Sea Bream (Sparus aurata L., 1758) given Feed with Different Protein Contents. Indian J. Anim. Res. 2018, 53, 510–514. [Google Scholar] [CrossRef]
- Akyol, O.; Gamsiz, K. Age and Growth of Adult Gilthead Seabream (Sparus aurata L.) in the Aegean Sea. J. Mar. Biol. Assoc. U. K. 2011, 91, 1255–1259. [Google Scholar] [CrossRef]
- Ceyhan, T.; Akyol, O.; Erdem, M. Length-Weight Relationships of Fishes from Gökova Bay, Turkey (Aegean Sea). Turk. J. Zool. 2009, 33, 69–72. [Google Scholar] [CrossRef]
- Sangun, L.; Akamca, E.; Akar, M.A. Weight-Length Relationships for 39 Fish Species from the North-Eastern Mediterranean Coast of Turkey. Turk. J. Fish. Aquat. Sci. 2007, 7, 37–40. [Google Scholar]
- Cicek, E.; Avsar, D.; Yeldan, H.; Ozutok, M. Length–Weight Relationships for 31 Teleost Fishes Caught by Bottom Trawl Net in the Babadillimani Bight (Northeastern Mediterranean). J. Appl. Ichthyol. 2006, 22, 290–292. [Google Scholar] [CrossRef]
- Eid, A.M.S.; Ali, B.A.; Eldahrawy, A.A.; Salama, F.; Abd El-Naby, A.S. Growth Performance and Survival of Gilthead Seabream Sparus aurata Larvae Fed Rotifer and Artemia. Egypt. J. Nutr. Feed. 2018, 21, 899–907. [Google Scholar] [CrossRef]
- Alarcón, J.A.; Magoulas, A.; Georgakopoulos, T.; Zouros, E.; Alvarez, M.C. Genetic Comparison of Wild and Cultivated European Populations of the Gilthead Sea Bream (Sparus aurata). Aquaculture 2004, 230, 65–80. [Google Scholar] [CrossRef]
- Rossi, A.R.; Perrone, E.; Sola, L. Genetic Structure of Gilthead Seabream, Sparus aurata, in the Central Mediterranean Sea. Cent. Eur. J. Biol. 2006, 1, 636–647. [Google Scholar] [CrossRef]
- Loukovitis, D.; Sarropoulou, E.; Vogiatzi, E.; Tsigenopoulos, C.S.; Kotoulas, G.; Magoulas, A.; Chatziplis, D. Genetic Variation in Farmed Populations of the Gilthead Sea Bream Sparus aurata in Greece Using Microsatellite DNA Markers. Aquac. Res. 2012, 43, 239–246. [Google Scholar] [CrossRef]
- Maroso, F.; Gkagkavouzis, K.; De Innocentiis, S.; Hillen, J.; do Prado, F.; Karaiskou, N.; Taggart, J.B.; Carr, A.; Nielsen, E.; Triantafyllidis, A.; et al. Genome-Wide Analysis Clarifies the Population Genetic Structure of Wild Gilthead Sea Bream (Sparus aurata). PLoS ONE 2021, 16, e0236230. [Google Scholar] [CrossRef] [PubMed]
- Gkagkavouzis, K.; Karaiskou, N.; Katopodi, T.; Leonardos, I.; Abatzopoulos, T.J.; Triantafyllidis, A. The Genetic Population Structure and Temporal Genetic Stability of Gilthead Sea Bream Sparus aurata Populations in the Aegean and Ionian Seas, Using Microsatellite DNA Markers. J. Fish Biol. 2019, 94, 606–613. [Google Scholar] [CrossRef]
- Moretti, A.; Pedini Fernandez-Criado, M.; Cittolin, G.; Guidastri, R. Manual on Hatchery Production of Seabass and Gilthead Seabream; FAO: Rome, Italy, 1999; Volume 1. [Google Scholar]
- Laird, L.M. Mariculture Overview. In Encyclopedia of Ocean Sciences; Steele, J.H., Ed.; Academic Press: Cambridge, MA, USA, 2001; pp. 1572–1577. ISBN 978-0-12-227430-5. [Google Scholar]
- Thorland, I.; Papaioannou, N.; Kottaras, L.; Refstie, T.; Papasolomontos, S.; Rye, M. Family Based Selection for Production Traits in Gilthead Seabream (Sparus aurata) and European Sea Bass (Dicentrarchus labrax) in Greece. Aquaculture 2007, 272, S314. [Google Scholar] [CrossRef]
- Zenger, K.R.; Khatkar, M.S.; Jones, D.B.; Khalilisamani, N.; Jerry, D.R.; Raadsma, H.W. Genomic Selection in Aquaculture: Application, Limitations and Opportunities with Special Reference to Marine Shrimp and Pearl Oysters. Front. Genet. 2018, 9, 693. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Zohar, Y.; Pankhurst, N.; Kagaw, H. Reproduction and Broodstock Management. In Sparidae, Biology and Aquaculture of Gilthead Sea Bream and Other Species; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 95–131. [Google Scholar]
- Cerdà, J.; Fabra, M.; Raldúa, D. Physiological and Molecular Basis of Fish Oocyte Hydration. In The Fish Oocyte; Springer: Berlin/Heidelberg, Germany, 2007; pp. 349–396. [Google Scholar]
- Carnevali, O.; Mosconi, G.; Cardinali, M.; Meiri, I.; Polzonetti-Magni, A. Molecular Components Related to Egg Viability in the Gilthead Sea Bream, Sparus aurata. Mol. Reprod. Dev. 2001, 58, 330–335. [Google Scholar] [CrossRef]
- Uçal, İ.O. Sea Bream (Sparus aurata L., 1758) Fry Production in Turkey. Turk. J. Fish. Aquat. Sci. 2002, 2, 77–81. [Google Scholar]
- Elbal, M.T.; García Hernández, M.P.; Lozano, M.T.; Agulleiro, B. Development of the Digestive Tract of Gilthead Sea Bream (Sparus aurata L.). Light and Electron Microscopic Studies. Aquaculture 2004, 234, 215–238. [Google Scholar] [CrossRef]
- René, F. Rearing of Gilt-Head Sparus aurata. In Early Life History of Fish; Blaxter, J.H.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1974; p. 747. [Google Scholar]
- Papandroulakis, N.; Divanach, P.; Kentouri, M. Enhanced Biological Performance of Intensive Sea Bream (Sparus aurata) Larviculture in the Presence of Phytoplankton with Long Photophase. Aquaculture 2002, 204, 45–63. [Google Scholar] [CrossRef]
- Webster, C.D.; Lim, C.E. Nutrient Requirements and Feeding of Finfish for Aquaculture, 1st ed.; CABI: Wallingford, UK, 2002; ISBN 978-0-85199-519-9. [Google Scholar]
- Lorenzo-Felipe, Á.; Shin, H.S.; León-Bernabeu, S.; Pérez-García, C.; Zamorano, M.J.; Pérez-Sánchez, J.; Afonso-López, J.M. The Effect of the Deformity Genetic Background of the Breeders on the Spawning Quality of Gilthead Seabream (Sparus aurata L.). Front. Mar. Sci. 2021, 8, 656901. [Google Scholar] [CrossRef]
- Galeotti, M.; Beraldo, P.; de Dominis, S.; D’Angelo, L.; Ballestrazzi, R.; Musetti, R.; Pizzolito, S.; Pinosa, M. A Preliminary Histological and Ultrastructural Study of Opercular Anomalies in Gilthead Sea Bream Larvae (Sparus aurata). Fish Physiol. Biochem. 2000, 22, 151–157. [Google Scholar] [CrossRef]
- Koumoundouros, G.; Oran, G.; Divanach, P.; Stefanakis, S.; Kentouri, M. The Opercular Complex Deformity in Intensive Gilthead Sea Bream (Sparus aurata L.) Larviculture. Moment of Apparition and Description. Aquaculture 1997, 156, 165–177. [Google Scholar] [CrossRef]
- Boglione, C.; Gavaia, P.; Koumoundouros, G.; Gisbert, E.; Moren, M.; Fontagné, S.; Witten, P.E. Skeletal Anomalies in Reared European Fish Larvae and Juveniles. Part 1: Normal and Anomalous Skeletogenic Processes. Rev. Aquac. 2013, 5, S99–S120. [Google Scholar] [CrossRef]
- Borrego, J.J.; Labella, A.M.; Ortiz-Delgado, J.B.; Sarasquete, C. Updated of the Pathologies Affecting Cultured Gilthead Seabream, Sparus aurata. Ann. Aquac. Res. 2017, 4, 1033. [Google Scholar] [CrossRef]
- Vendramin, N.; Zrncic, S.; Padrós, F.; Oraic, D.; Le Breton, A.; Zarza, C.; Olesen, N.J. Fish Health in Mediterranean Aquaculture, Past Mistakes and Future Challenges. Bull. Eur. Assoc. Fish Pathol. 2016, 36, 38–45. [Google Scholar]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral Nervous Necrosis in Gilthead Sea Bream (Sparus aurata) Caused by Reassortant Betanodavirus RGNNV/SJNNV: An Emerging Threat for Mediterranean Aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef]
- Castri, J.; Thiéry, R.; Jeffroy, J.; de Kinkelin, P.; Raymond, J.C. Sea Bream Sparus aurata, an Asymptomatic Contagious Fish Host for Nodavirus. Dis. Aquat. Organ. 2001, 47, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Savoca, S.; Abbadi, M.; Toffan, A.; Salogni, C.; Iaria, C.; Capparucci, F.; Quartesan, R.; Alborali, G.L.; Guarnera, S.; Cangemi, G.; et al. Betanodavirus Infection Associated with Larval Enteropathy as a Cause of Mortality in Cultured Gilthead Sea Bream (Sparus aurata, Linnaeus, 1758). Aquaculture 2021, 541, 736844. [Google Scholar] [CrossRef]
- Faggion, S.; Carnier, P.; Franch, R.; Babbucci, M.; Pascoli, F.; Dalla Rovere, G.; Caggiano, M.; Chavanne, H.; Toffan, A.; Bargelloni, L. Viral Nervous Necrosis Resistance in Gilthead Sea Bream (Sparus aurata) at the Larval Stage: Heritability and Accuracy of Genomic Prediction with Different Training and Testing Settings. Genet. Sel. Evol. 2023, 55, 22. [Google Scholar] [CrossRef] [PubMed]
- Muniesa, A.; Basurco, B.; Aguilera, C.; Furones, D.; Reverté, C.; Sanjuan-Vilaplana, A.; Jansen, M.D.; Brun, E.; Tavornpanich, S. Mapping the Knowledge of the Main Diseases Affecting Sea Bass and Sea Bream in Mediterranean. Transbound. Emerg. Dis. 2020, 67, 1089–1100. [Google Scholar] [CrossRef]
- Fernandes, T.; Herlin, M.; Belluga, M.D.L.; Ballón, G.; Martinez, P.; Toro, M.A.; Fernández, J. Estimation of Genetic Parameters for Growth Traits in a Hatchery Population of Gilthead Sea Bream (Sparus aurata L.). Aquac. Int. 2017, 25, 499–514. [Google Scholar] [CrossRef]
- Saura, M.; Caballero, A.; Santiago, E.; Fernández, A.; Morales-González, E.; Fernández, J.; Cabaleiro, S.; Millán, A.; Martínez, P.; Palaiokostas, C.; et al. Estimates of Recent and Historical Effective Population Size in Turbot, Seabream, Seabass and Carp Selective Breeding Programmes. Genet. Sel. Evol. 2021, 53, 85. [Google Scholar] [CrossRef]
- Palaiokostas, C.; Ferraresso, S.; Franch, R.; Houston, R.D.; Bargelloni, L. Genomic Prediction of Resistance to Pasteurellosis in Gilthead Sea Bream (Sparus aurata) Using 2b-RAD Sequencing. G3 Genes Genomes Genet. 2016, 6, 3693–3700. [Google Scholar] [CrossRef]
- Chavanne, H.; Janssen, K.; Hofherr, J.; Contini, F.; Haffray, P.; Komen, H.; Nielsen, E.E.; Bargelloni, L. A Comprehensive Survey on Selective Breeding Programs and Seed Market in the European Aquaculture Fish Industry. Aquac. Int. 2016, 24, 1287–1307. [Google Scholar] [CrossRef]
- Griot, R.; Allal, F.; Phocas, F.; Brard-Fudulea, S.; Morvezen, R.; Haffray, P.; François, Y.; Morin, T.; Bestin, A.; Bruant, J.-S.; et al. Optimization of Genomic Selection to Improve Disease Resistance in Two Marine Fishes, the European Sea Bass (Dicentrarchus labrax) and the Gilthead Sea Bream (Sparus aurata). Front. Genet. 2021, 12, 665920. [Google Scholar] [CrossRef]
- Aslam, M.L.; Carraro, R.; Bestin, A.; Cariou, S.; Sonesson, A.K.; Bruant, J.-S.; Haffray, P.; Bargelloni, L.; Meuwissen, T.H.E. Genetics of Resistance to Photobacteriosis in Gilthead Sea Bream (Sparus aurata) Using 2b-RAD Sequencing. BMC Genet. 2018, 19, 43. [Google Scholar] [CrossRef]
- Knibb, W.; Gorshkova, G.; Gorshkov, S. Selection for Growth in the Gilthead Seabream, Sparus aurata L. Isr. J. Aquac. 1997, 49, 57–66. [Google Scholar]
- Cameron Brown, R.; Woolliams, J.A.; McAndrew, B.J. Factors Influencing Effective Population Size in Commercial Populations of Gilthead Seabream, Sparus aurata. Aquaculture 2005, 247, 219–225. [Google Scholar] [CrossRef]


| Incubation | Hatching | Larval Stage | Fry Stage | |
|---|---|---|---|---|
| Water temperature (°C) | 15–17.5 | 15–17 | 15–20 | 20 |
| Salinity (ppt) | 35–38 | 35–38 | 35 | 30 |
| Photoperiod (h) | - | 16:8 | 16:8 | 14:10 |
| Water renewal (time/day) | 12 | 12 | 8–12 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mhalhel, K.; Levanti, M.; Abbate, F.; Laurà, R.; Guerrera, M.C.; Aragona, M.; Porcino, C.; Briglia, M.; Germanà, A.; Montalbano, G. Review on Gilthead Seabream (Sparus aurata) Aquaculture: Life Cycle, Growth, Aquaculture Practices and Challenges. J. Mar. Sci. Eng. 2023, 11, 2008. https://doi.org/10.3390/jmse11102008
Mhalhel K, Levanti M, Abbate F, Laurà R, Guerrera MC, Aragona M, Porcino C, Briglia M, Germanà A, Montalbano G. Review on Gilthead Seabream (Sparus aurata) Aquaculture: Life Cycle, Growth, Aquaculture Practices and Challenges. Journal of Marine Science and Engineering. 2023; 11(10):2008. https://doi.org/10.3390/jmse11102008
Chicago/Turabian StyleMhalhel, Kamel, Maria Levanti, Francesco Abbate, Rosaria Laurà, Maria Cristina Guerrera, Marialuisa Aragona, Caterina Porcino, Marilena Briglia, Antonino Germanà, and Giuseppe Montalbano. 2023. "Review on Gilthead Seabream (Sparus aurata) Aquaculture: Life Cycle, Growth, Aquaculture Practices and Challenges" Journal of Marine Science and Engineering 11, no. 10: 2008. https://doi.org/10.3390/jmse11102008