
The Black Sea Zooplankton Mortality, Decomposition, and Sedimentation Measurements Using Vital Dye and Short-Term Sediment Traps


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Experimental Design
2.2. Design and Exposure Conditions of Sediment Traps
2.3. Evaluation of Total Abundance of Zooplankton and Fraction of Live Organisms (FLO)
2.4. Estimation of Abundance and Physiological Activity of Bacteria
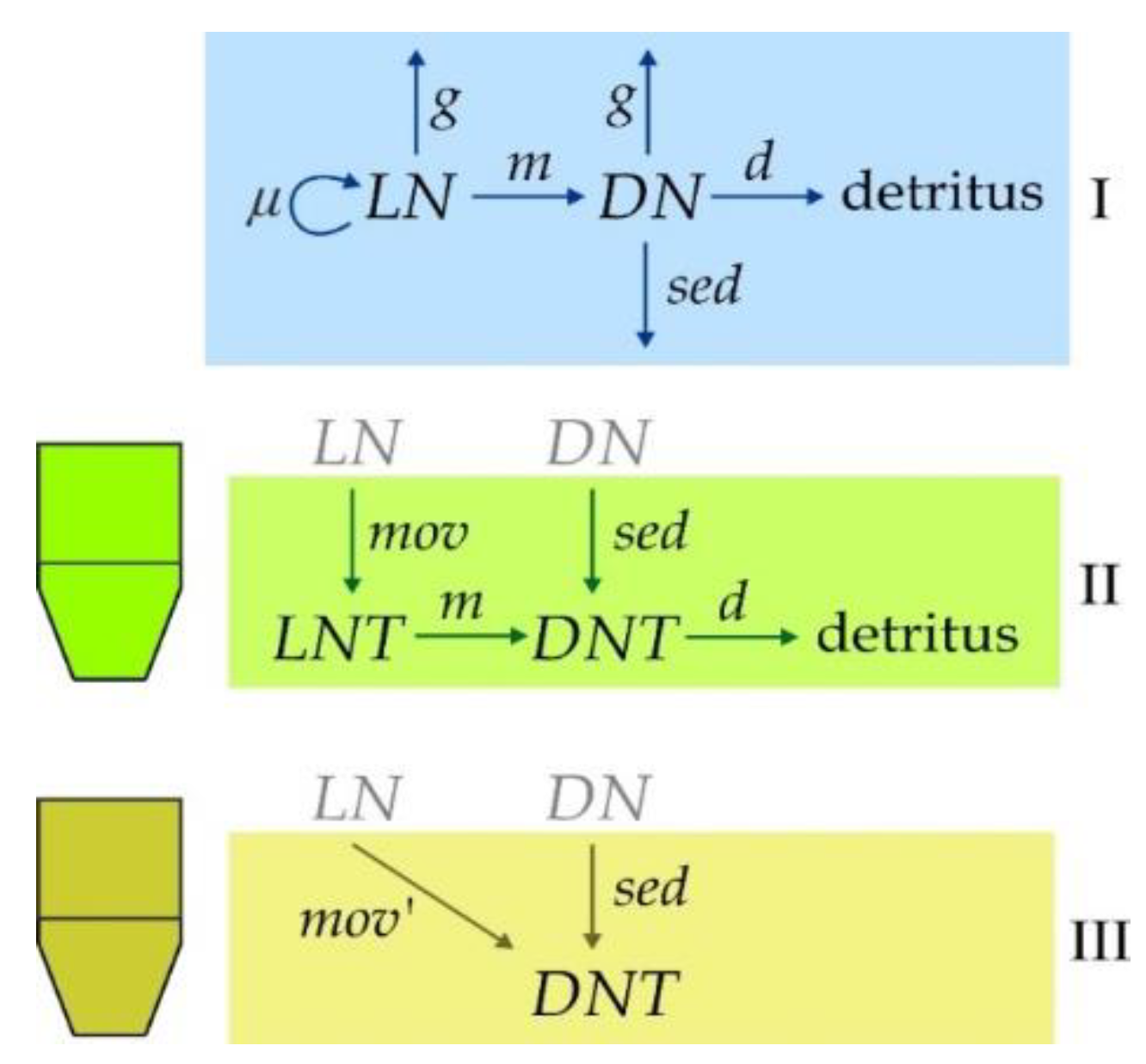
2.5. Simulation Model of Live and Dead Zooplankton Dynamics in Sediment Traps and Water Column above Them
3. Results
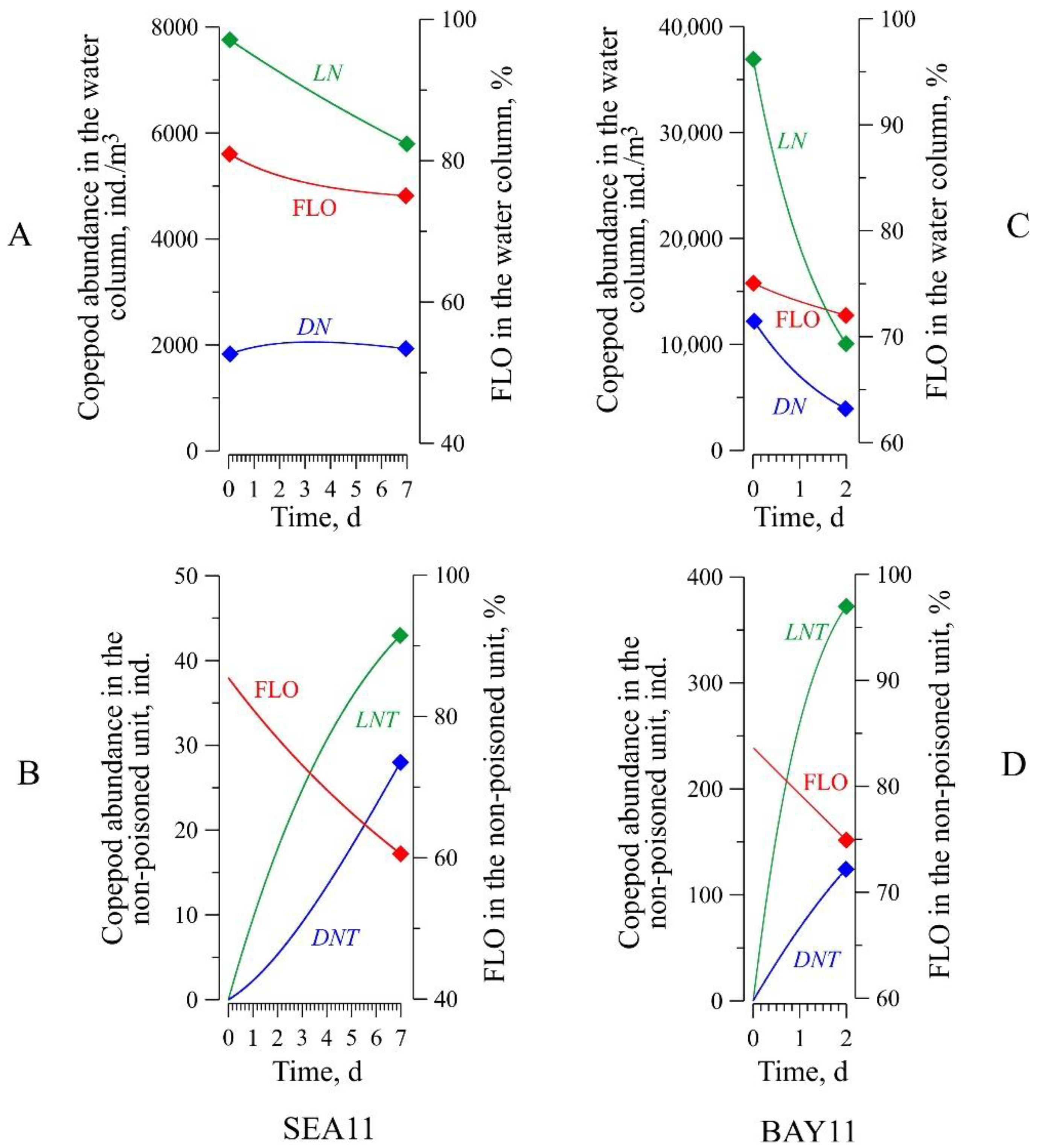
3.1. Species Composition and Dynamics of Zooplankton in the Water Column above the Sediment Trap
3.2. Accumulation of Zooplankton in the Sediment Trap
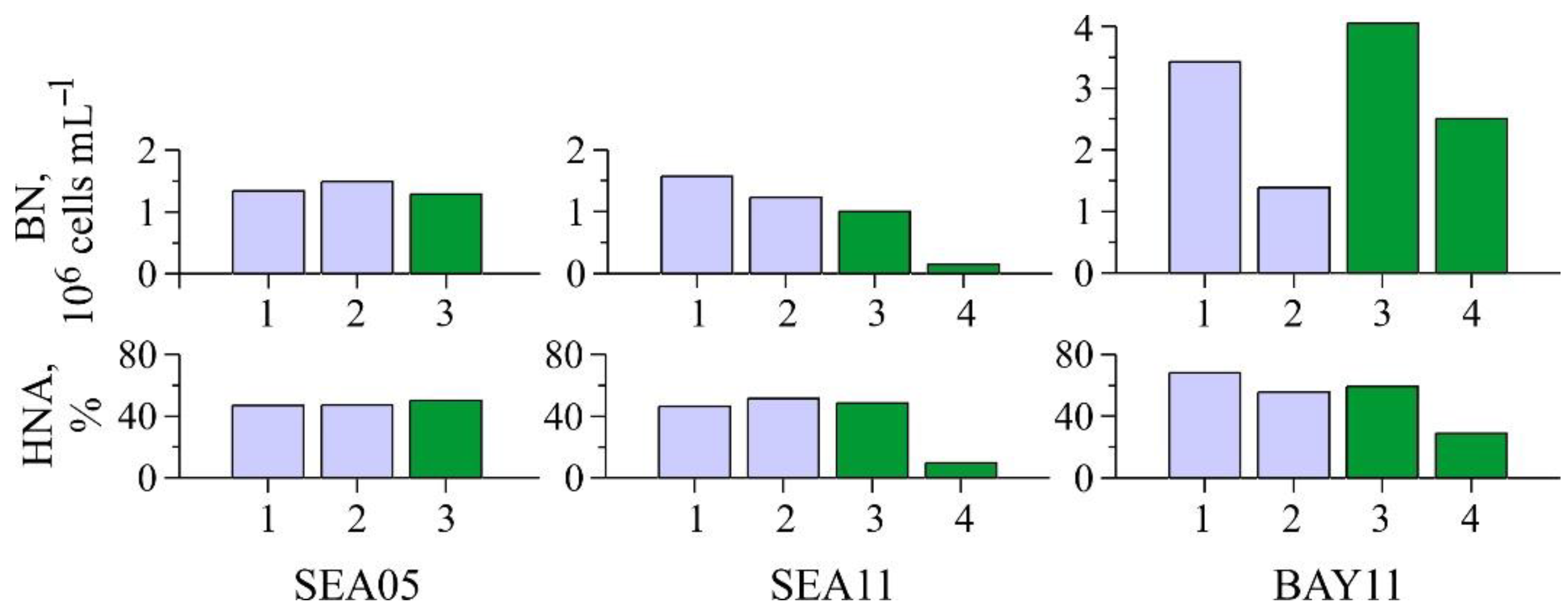
3.3. Dynamics of Bacterioplankton in the Water Column and the Traps
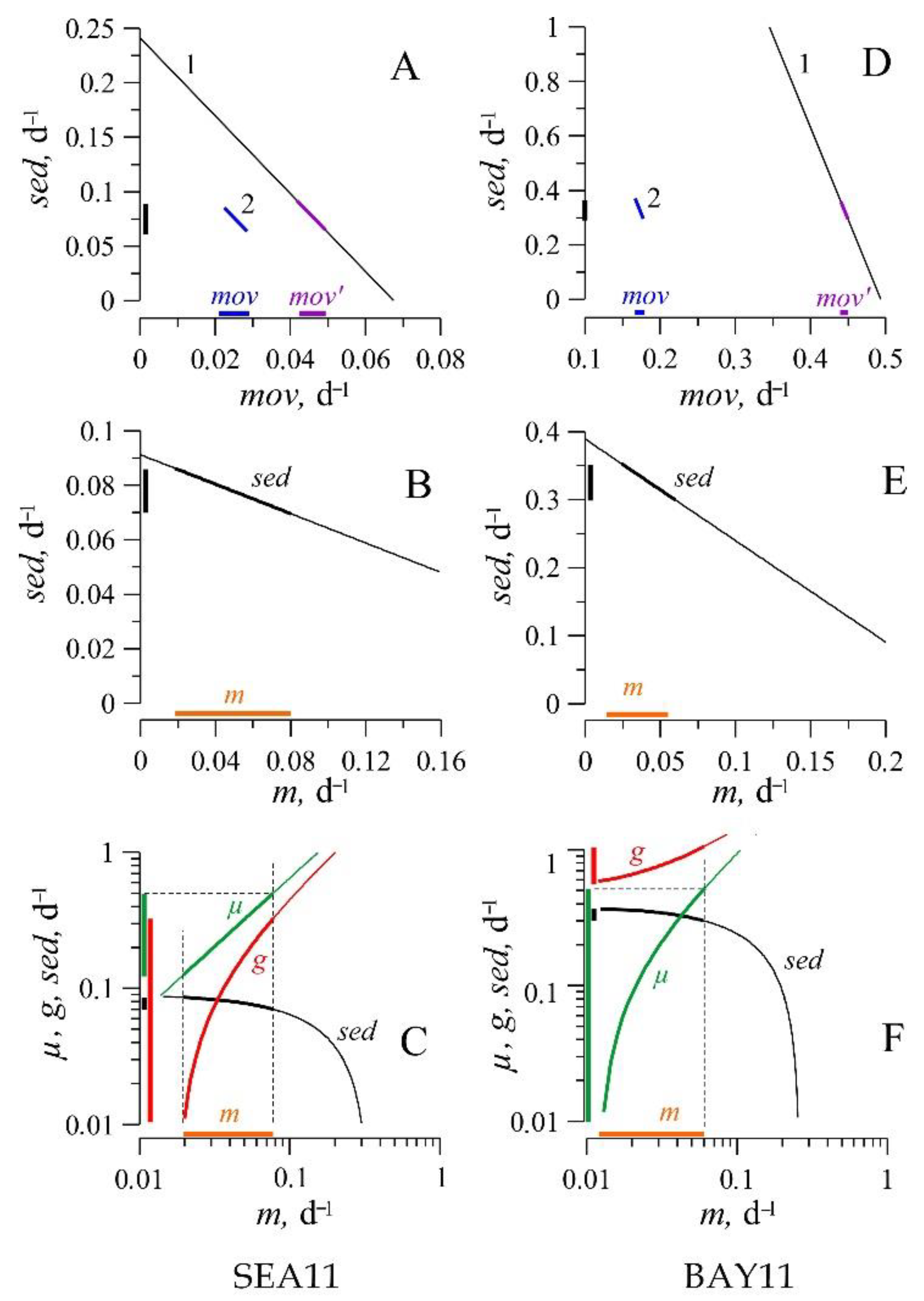
3.4. Results of Numerical Experiments
4. Discussion
4.1. Rates of the Processes Controlling Copepod Carcasses Dynamics in the Water Column
4.2. Validity and Applicability of Existing Field Methods for Measuring Zooplankton Non-Consumptive Mortality
4.3. The Problem of Live Copepods-Swimmers in the Trap
4.4. Applicability of Other Models to Our Field Data
5. Conclusions
- Significant changes in the abundance of copepod carcasses (from 280 to 12,443 ind. m−3) and FLO (53 to 81%) were observed in Sevastopol Bay and adjacent waters over short time periods, which indicated a high variability of zooplankton non-consumptive mortality (m), sedimentation (sed), and decomposition rates of dead organisms (d).
- Despite the high concentrations of copepod carcasses in the water column, the rates of their enrichment in the traps proved to be extremely low (no more than 20 specimens per day per trap unit), which could be due to intense turbulent mixing of the waters. The rates of non-consumptive mortality (m) and sedimentation (sed) of copepods were comparable with each other.
- The obtained estimates of the sedimentation rate of copepod carcasses (0.012 to 0.39 d−1) were comparable in value with the rate of their microbial decomposition (0.13 and 0.05 d−1 in the bay and adjacent waters, respectively), which confirmed the hypothesis on microbial decomposition as one of the key controls of FLO in zooplankton. The influence of sedimentation processes on the dynamics of carcasses in coastal waters seems to be greatly overestimated.
- The carcass sedimentation rate (sed) and the flows of swimmers into the traps (mov) were significantly higher in the bay than in the adjacent waters, which may be explained by a difference in hydrological regimes at the stations. Weaker turbulent mixing appeared to increase the contribution of the above processes to the control of FLO in zooplankton.
- The models used to process and interpret the results of the short-term sedimentation experiments should take into account the zooplankton swimmers and their death in the sedimentation trap. Otherwise, mortality and sedimentation rates may be estimated incorrectly.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M. The key role of zooplankton in ecosystem services: A perspective of interaction between zooplankton and fish recruitment. Ecol. Indic. 2021, 129, 107867. [Google Scholar] [CrossRef]
- Ohman, M.D.; Hirche, H.-J. Density-dependent mortality in an oceanic copepod population. Nature 2001, 412, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Fasham, M.J.R. (Ed.) Ocean Biogeochemistry: The Role of the Ocean Carbon Cycle in Global Change, Global Change; Springer: New York, NY, USA, 2003; Volume XVIII, p. 297. [Google Scholar]
- Koval, L.G. Zoo- and Necrozooplankton of the Black Sea; Naukova: Kiev, Ukraine, 1984; p. 128. (In Russian) [Google Scholar]
- Dubovskaya, O.P.; Tang, K.W.; Gladyshev, M.I.; Kirillin, G.; Buseva, Z.; Kasprzak, P.; Tolomeev, A.P.; Grossart, H.P. Estimating in situ zooplankton non-predation mortality in an oligo-mesotrophic lake from sediment trap data: Caveats and Reality Check. PLoS ONE 2015, 10, e0131431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, D.T.; Tang, K.W. Spatial and temporal distributions of live and dead Copepods in the lower Chesapeake Bay (Virginia, USA). Estuaries Coasts 2011, 34, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Farran, G.P. Biscayan plankton collected during a cruise of H.M.S. Research, 1900. Pt. 14: The Copepoda. Zool. J. Linn. Soc. 1926, 36, 219–310. [Google Scholar] [CrossRef]
- Tang, K.W.; Gladyshev, M.I.; Dubovskaya, O.P.; Kirillin, G.; Grossart, H.P. Zooplankton carcasses and non-predatory mortality in freshwater and inland sea environments. J. Plankton Res. 2014, 36, 597–612. [Google Scholar] [CrossRef] [Green Version]
- Di Capua, I.; Mazzocchi, M.G. Non-predatory mortality in Mediterranean coastal copepods. Mar. Biol. 2017, 164, 198. [Google Scholar] [CrossRef]
- Tang, K.W.; Freund, C.S.; Schweitzer, C.L. Occurrence of copepod carcasses in the lower Chesapeake Bay and their decomposition by ambient microbes. Estuar. Coast. Shelf Sci. 2006, 68, 499–508. [Google Scholar] [CrossRef]
- Tolomeev, A.P.; Dubovskaya, O.P.; Kirillin, G.; Buseva, Z.; Kolmakova, O.V.; Grossart, H.-P.; Tang, K.W.; Gladyshev, M.I. Degradation of dead cladoceran zooplankton and their contribution to organic carbon cycling in stratified lakes: Field observation and model prediction. J. Plankton Res. 2022, 44, 386–400. [Google Scholar] [CrossRef]
- Kydd, J.; Rajakaruna, H.; Briski, E.; Bailey, S. Examination of a high resolution laser optical plankton counter and FlowCAM for measuring plankton concentration and size. J. Sea Res. 2018, 133, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Colas, F.; Tardivel, M.; Perchoc, J.; Lunven, M.; Forest, B.; Guyader, G.; Danielou, M.M.; Le Mestre, S.; Bourriau, P.; Antajan, E.; et al. The ZooCAM, a new in-flow imaging system for fast onboard counting, sizing and classification of fish eggs and metazooplankton. Prog. Oceanogr. 2018, 166, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Litvinyuk, D.A.; Altukhov, D.A.; Mukhanov, V.S.; Popova, E.V. Dynamics of live Copepoda in plankton of Sevastopol Bay and open coastal waters (the Black Sea) in 2010–2011. Mar. Ecol. J. 2011, 10, 56–65. (In Russian) [Google Scholar]
- Mukhanov, V.; Litvinyuk, D. Microbial control of live/dead zooplankton ratio in Sevastopol Bay. Ecol. Montenegrina 2017, 11, 42–48. [Google Scholar] [CrossRef]
- Pavlova, E.V.; Melnikova, E.B. Zooplankton in inshore waters of the south-western Crimea (1998–2006). Mar. Ecol. J. 2011, 10, 33–42. (In Russian) [Google Scholar]
- Hirst, A.G.; Kiørboe, T. Mortality of marine planktonic copepods: Global rates and patterns. Mar. Ecol. Prog. Ser. 2002, 230, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Dubovskaya, O.P.; Gladyshev, M.I.; Gubanov, V.G.; Makhutova, O.N. Study of non-consumptive mortality of Crustacean zooplankton in a Siberian reservoir using staining for live/dead sorting and sediment traps. Hydrobiologia 2003, 504, 223–227. [Google Scholar] [CrossRef]
- Gries, T.; Güde, H. Estimates of the nonconsumptive mortality of mesozooplankton by measurement of sedimentation losses. Limnol. Oceanogr. 1999, 44, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Dubovskaya, O.P.; Tolomeev, A.P.; Kirillin, G.; Buseva, Z.; Tang, K.W.; Gladyshev, M.I. Effects of water column processes on the use of sediment traps to measure zooplankton non-predatory mortality: A mathematical and empirical assessment. J. Plankton Res. 2018, 40, 91–106. [Google Scholar] [CrossRef]
- Kirillin, G.; Grossart, H.-P.; Tang, K.W. Modeling sinking rate of zooplankton carcasses: Effects of stratification and mixing. Limnol. Oceanogr. 2012, 57, 881–894. [Google Scholar] [CrossRef] [Green Version]
- Lukashin, V.N.; Klyuvitkin, A.A.; Lisitzin, A.P.; Novigatsky, A.N. The MSL-110 small sediment trap. Oceanology 2011, 51, 699–703. [Google Scholar] [CrossRef]
- Orekhova, N.A.; Varenik, A.V. Carrent hydrochemical regime of the Sevastopol Bay. Morskoy Gidrofiz. Zhurnal 2018, 34, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Gubanov, V.I.; Gubanova, A.D.; Rodionova, N.Y. Diagnosis of water trophicity in the Sevastopol bay and its offshore. In Current Issues in Aquaculture, Proceedings of the International Scientific Conference, Rostov-on-Don, Russia,28 September–2 October 2015; FGBNU “AzNIIRKH”: Rostov-on-Don, Russia, 2015; pp. 64–67. [Google Scholar]
- Tikhonova, E.A.; Burdiyan, N.V.; Soloveva, O.V.; Doroshenko, Y.V. The estimation of the sevastopol bays ecological state on basic chemical and microbiological criteria. Ecol. Environ. Conserv. 2018, 24, 1574–1584. [Google Scholar]
- Aleksandrov, B.; Arashkevich, E.G.; Gubanova, A.D.; Korshenko, A. Black Sea monitoring guidelines: Mesozooplankton. Publ. EMBLAS Project 2014, BSC, 31. Available online: http://emblasproject.org/wp-content/uploads/2017/01/Mesozooplankton_Final-July2015-PA3-f.pdf (accessed on 29 April 2020).
- Postel, L.; Fock, H.; Hagen, W. Biomass and abundance. In ICES Zooplankton Methodology Manual; Harris, R.P., Wiebe, P.H., Lenz, J., Skjoldal, H.R., Huntley, M., Eds.; Academic Press: London, UK, 2000; pp. 83–174. [Google Scholar]
- Lytvynyuk, D.A.; Mukhanov, V.S. Advanced method for identifying alive organisms in marine zooplankton stained with neutral red and fluorescein diacetate. Mar. Ecol. J. 2012, 11, 45–54. (In Russian) [Google Scholar]
- Brookes, J.D.; Geary, S.M.; Ganf, G.G.; Burch, M.D. Use of FDA and flow cytometry to assess metabolic activity as an indicator of nutrient status in phytoplankton. Mar. Freshw. Res. 2000, 51, 817–823. [Google Scholar] [CrossRef]
- Litvinyuk, D.; Aganesova, L.; Mukhanov, V. Identifying alive versus dead Copepods in culture of Calanipeda aquae dulcis after staining them with neutral red and fluorescein diacetate. Ekologiyamorya 2009, 78, 65–69. (In Russian) [Google Scholar]
- Marie, D.; Partensky, F.; Jacquet, S.; Vaulot, D. Enumeration and cell cycle analysis of natural populations of marine picoplankton by flow cytometry using the nucleic acid stain SYBR Green, I. Appl. Environ. Microbiol. 1997, 63, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasol, J.M.; Del Giorgio, P.A. Using flow cytometry for counting natural planktonic bacteria and understanding the structure of planktonic bacterial communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Servais, P.; Casamayor, E.O.; Courties, C.; Catala, P.; Parthuisot, N.; Lebaron, P. Activity and diversity of bacterial cells with high and low nucleic acid content. Aquat. Microb. Ecol. 2003, 33, 41–51. [Google Scholar] [CrossRef]
- Tang, K.W.; Bickel, S.L.; Dziallas, C.; Grossart, H.P. Microbial activities accompanying decomposition of cladoceran and copepod carcasses under different environmental conditions. Aquat. Microb. Ecol. 2009, 57, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Gubanova, A.D.; Garbazey, O.A.; Popova, E.V.; Altukhov, D.A.; Mukhanov, V.S. Oithona davisae: Naturalization in the Black Sea, interannual and seasonal dynamics, and effect on the structure of the planktonic copepod community. Oceanology 2019, 59, 912–919. [Google Scholar] [CrossRef]
- Garbazey, O.A.; Popova, E.V.; Gubanova, A.D.; Altukhov, D.A. First report of the occurrence of Pseudodiaptomus marinus Sato, 1913 (Copepoda: Calanoida: Pseudodiaptomidae) in the Black Sea (Sevastopol Bay). Mar. Biol. J. 2016, 1, 78–80. [Google Scholar] [CrossRef]
- Richardson, A.J.; Verheye, H.M. The relative importance of food and temperature to copepod egg production and somatic growth in the southern Benguela upwelling system. J. Plankton Res. 1998, 20, 2379–2399. [Google Scholar] [CrossRef]
- Richardson, A.J.; Verheye, H.M. Growth rates of copepods in the southern Benguela upwelling system: The interplay between body size and food. Limnol. Oceanogr. 1999, 44, 382–392. [Google Scholar] [CrossRef]
- Richardson, A.J.; Verheye, H.M.; Herbert, V.; Rogers, C.; Arendse, L.M. Egg production, somatic growth and productivity of copepods in the Benguela Current system and Angola-Benguela Front: BENEFIT Marine Science. S. Afr. J. Sci. 2001, 97, 251–257. Available online: https://hdl.handle.net/10520/EJC97315 (accessed on 19 June 2022).
- Persad, G.; Webber, M. The use of Ecopath software to model trophic interactions within the zooplankton community of Discovery Bay, Jamaica. Open Mar. Biol. J. 2009, 3, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, L.A.; Roff, J.C. Abundances, growth rates, and production of tropical neritic copepods off Kingston, Jamaica. Mar. Biol. 1990, 106, 79–89. [Google Scholar] [CrossRef]
- Stepanov, V.N.; Svetlichnyyi, L.S. Research of Hydromechanical Characteristics of Plankton Copepods; Naukova Dumka: Kiev, Ukraine, 1981; p. 128. (In Russian) [Google Scholar]
- Zelezinskaya, L.M. Natural Mortality of Some Forms of Ichthyo and Zooplankton of the Black Sea. Ph.D. Thesis, IBSS UAS, Odessa, Ukraine, 1966; p. 23. (In Russian). [Google Scholar]
- Dubovskaya, O.P.; Gladyshev, M.I.; Esimbekova, E.N.; Morozova, I.I.; Gol’d, Z.G.; Makhutova, O.N. Study of possible relation between seasonal dynamics of zooplankton non-consumptive mortality and water toxicity in a pond. Inland Water Biol. 2002, 3, 39–43. (In Russian) [Google Scholar]
- Frangoulis, C.; Skliris, N.; Lepoint, G.; Elkalay, K.; Goffart, A.; Pinnegar, J.K.; Hecq, J.-H. Importance of copepod carcasses versus fecal pellets in the upper water column of an oligotrophic area. Estuar. Coast. Shelf Sci. 2011, 92, 456–463. [Google Scholar] [CrossRef]
- Dubovskaya, O.P. Non-predatory mortality of the crustacean zooplankton, and its possible causes (a review). Zhumal Obshch. Biol. 2009, 70, 168–192. (In Russian) [Google Scholar]
- Dubovskaya, O.P.; Gladyshev, M.I.; Gubanov, V.G. Seasonal dynamics of number of alive and dead zooplankton in a small pond and some variants of mortality estimation. J. Gen. Biol. 1999, 60, 543–555, (Translated into English). [Google Scholar]
- Ivory, J.A.; Tang, K.W.; Takahashi, K. Use of Neutral Red in short-term sediment traps to distinguish between zooplankton swimmers and carcasses. Mar. Ecol. Prog. Ser. 2014, 505, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Gladyshev, M.I.; Gubanov, V.G. Seasonal dynamics of specific mortality of Bosmina longirostris in forest pond determined on the basis of counting of dead individuals. Dokl. Akad. Nauk. 1996, 348, 127–128. [Google Scholar]
- Coale, K.H. Labyrinth of doom: A device to minimize the “swimmer” component in sediment trap collections. Limnol. Oceanogr. 1990, 35, 1376–1381. [Google Scholar] [CrossRef]
- Hansell, D.A.; Newton, J.A. Design and evaluation of a “swimmer”-segregating particle interceptor trap. Limnol. Oceanogr. 1994, 39, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Tolomeev, A.P.; Kirillin, G.; Dubovskay, O.P.; Buseva, Z.F.; Gladyshev, M.I. Numerical modeling of vertical distribution of living and dead copepods Arctodiaptomus salinus in Salt Lake Shira. Contemp. Probl. Ecol. 2018, 11, 543–550. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Units |
|---|---|---|
| N0 | Initial zooplankton abundance in the water column * | ind. M−3 |
| Nt | Final zooplankton abundance in the water column * | ind. M−3 |
| FLO | Fraction of live organisms * | % |
| LN | Abundance of live organisms in the water column above the trap | ind. M−3 |
| LN0 | Initial abundance of live copepods in the water column above the trap * | ind. M−3 |
| LNt | Final abundance oflivecopepodsin the watercolumnabovethetrap * | ind. M−3 |
| rlive | Apparent specific rate of growth/loss of live copepods * | d−1 |
| rdead | Apparent specific rate of production/loss of dead copepods * | d−1 |
| DN | Abundance of dead copepods in the water column above the trap | ind. M−3 |
| DN0 | Initial abundance of dead copepods in the water column above the trap * | ind. M−3 |
| DNt | Final abundance of dead copepods in the water column above the trap * | ind. M−3 |
| LNT | Abundance of live organisms (swimmers) in the trap * | ind. |
| DNT | Abundance of carcasses in the trap * | ind. |
| M | Non–consumptive mortality rate | d−1 |
| g | Consumptive mortality rate | d−1 |
| d | Carcass decomposition rate | d−1 |
| µ | Specific growth rate | d−1 |
| sed | Sedimentation rate | d−1 |
| mov | Net flow of swimmers into the non–poisoned trap unit | d−1 |
| mov’ | Net flow of swimmers into the poisoned trap unit | d−1 |
| T | Duration of the experiment | d |
| Taxon | Water Column | Water Column | Non-Poisoned Unit | Poisoned Unit | ||||
|---|---|---|---|---|---|---|---|---|
| N0, ind. m−3 | FLO0, % | Nt, ind. m−3 | FLOt, % | LNT, ind. | DNT, ind. | LNT, ind. | DNT, ind. | |
| Experiment SEA05 (St. S; depth: 36 m; time of exposition: 1 day) | ||||||||
| Total Copepoda | Nd * | 53 | 849 | 67 | 12 | 0 | – | – |
| Acartia clausi | nd | 58 | 554 | 65 | 2 | 0 | – | – |
| Pseudocalanus elongatus | nd | 52 | 36 | 71 | 4 | 0 | – | – |
| Oithona similis | nd | 49 | 177 | 60 | 5 | 0 | – | – |
| Copepoda nauplii | nd | 88 | 144 | 94 | 10 | 1 | – | – |
| Pleopis polyphemoides | nd | nd | 29 | nd | 1 | 0 | – | – |
| Noctiluca scintillans | nd | nd | 7806 | nd | 5 | 0 | – | – |
| Cirripedia nauplii | nd | nd | 188 | nd | 3 | 1 | – | – |
| Bivalvia larvae | nd | nd | 87 | nd | 2 | 0 | – | – |
| Experiment SEA11 (St. S; depth: 16 m; time of exposition: 7 d) | ||||||||
| Total Copepoda | 9597 | 81 | 6181 | 75 | 43 | 28 | 0 | 178 |
| Acartia clausi | 801 | 87 | 1527 | 94 | 1 | 0 | 0 | 3 |
| Paracalanus parvus | 1858 | 67 | 1433 | 43 | 5 | 1 | 0 | 91 |
| Oithona similis | 445 | 88 | 203 | 47 | 0 | 0 | 0 | 7 |
| Oithona davisae | 2435 | 86 | 2859 | 86 | 10 | 11 | 0 | 32 |
| Harpacticoida | 0 | nd | 2 | nd | 27 | 4 | 0 | 19 |
| Copepoda nauplii | 861 | 100 | 797 | 95 | 10 | 7 | 0 | 21 |
| Oikopleura dioica | 0 | 85 | 0 | nd | 0 | 0 | 0 | 6 |
| Bivalvia larvae | 56 | nd | 135 | nd | 0 | 0 | 0 | 15 |
| Experiment BAY11 (St. B; depth: 8 m; time of exposition: 11 d) | ||||||||
| Total Copepoda | 49,691 | 75 | 13,981 | 72 | 372 | 125 | 0 | 1054 |
| Acartia clausi | 237 | 87 | 309 | 64 | 2 | 1 | 0 | 3 |
| Paracalanus parvus | 1707 | 32 | 459 | 4 | 0 | 1 | 0 | 11 |
| Pseudocalanus elongatus | 926 | 71 | 60 | 0 | 0 | 3 | 0 | 54 |
| Pseudodiaptomus marinus | 0 | 0 | 0 | 0 | 64 | 2 | 0 | 30 |
| Oithona similis | 250 | 89 | 75 | nd | 0 | 0 | 0 | 3 |
| Oithona davisae | 46,312 | 84 | 13,012 | 91 | 285 | 93 | 0 | 854 |
| Harpacticoida | 1,25 | nd | 0 | nd | 21 | 3 | 0 | 25 |
| Copepoda nauplii | 40 | nd | 89 | nd | 13 | 0 | 0 | 43 |
| Cirripedia nauplii | 584 | nd | 350 | nd | 8 | 2 | 0 | 12 |
| Oikopleura dioica | 0 | nd | 112 | nd | 0 | 0 | 0 | 19 |
| Bivalvia larvae | 1229 | nd | 131 | nd | 0 | 0 | 0 | 22 |
| Polychaeta larvae | 40 | nd | 62 | nd | 1 | 0 | 0 | 39 |
| Gastropoda larvae | 90 | nd | 44 | nd | 0 | 0 | 0 | 5 |
| Experiment | d | mov | mov’ | sed | m | g | µ | |
|---|---|---|---|---|---|---|---|---|
| Copepoda | SEA11 | 0.02 | 0.02–0.04 | 0.08–0.10 | 0.05–0.01 | 0.03–0.13 | 0.00–0.40 | 0.00–0.50 |
| BAY11 | 0.05 | 0.24–0.29 | 0.59–0.64 | 0.16–0.06 | 0.08–0.19 | 0.60–0.97 | 0.00–0.50 | |
| O. davisae | SEA11 | 0.02 | 0.02–0.03 | 0.04–0.05 | 0.08–0.07 | 0.02–0.08 | 0.00–0.32 | 0.12–0.50 |
| BAY11 | 0.05 | 0.17–0.18 | 0.44–0.45 | 0.35–0.30 | 0.01–0.06 | 0.60–1.00 | 0.00–0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Litvinyuk, D.; Mukhanov, V.; Evstigneev, V. The Black Sea Zooplankton Mortality, Decomposition, and Sedimentation Measurements Using Vital Dye and Short-Term Sediment Traps. J. Mar. Sci. Eng. 2022, 10, 1031. https://doi.org/10.3390/jmse10081031
Litvinyuk D, Mukhanov V, Evstigneev V. The Black Sea Zooplankton Mortality, Decomposition, and Sedimentation Measurements Using Vital Dye and Short-Term Sediment Traps. Journal of Marine Science and Engineering. 2022; 10(8):1031. https://doi.org/10.3390/jmse10081031
Chicago/Turabian StyleLitvinyuk, Daria, Vladimir Mukhanov, and Vladislav Evstigneev. 2022. "The Black Sea Zooplankton Mortality, Decomposition, and Sedimentation Measurements Using Vital Dye and Short-Term Sediment Traps" Journal of Marine Science and Engineering 10, no. 8: 1031. https://doi.org/10.3390/jmse10081031