Transient Existence of Circulating Mesenchymal Stem Cells in the Deep Veins in Humans Following Long Bone Intramedullary Reaming

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
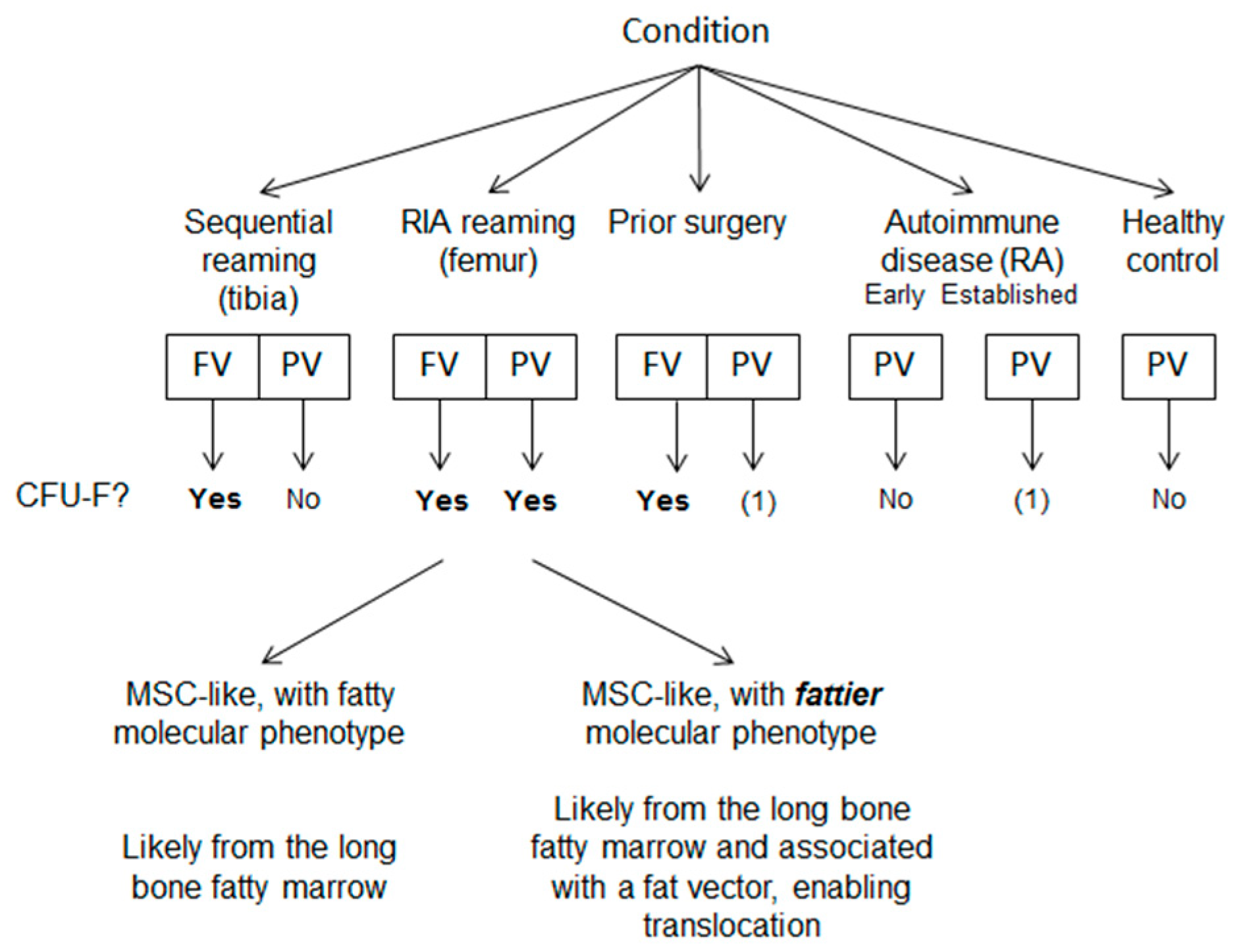
2.1. Donors and Tissue Collection
2.2. Preparation of Tissue and Generation of MSC Cultures
2.3. Colony-Forming Unit-Fibroblast Assay
2.4. Phenotypic Analysis of Putative Blood-Derived MSCs
2.5. Osteogenic Differentiation Assay
2.6. Gene Expression of MSCs
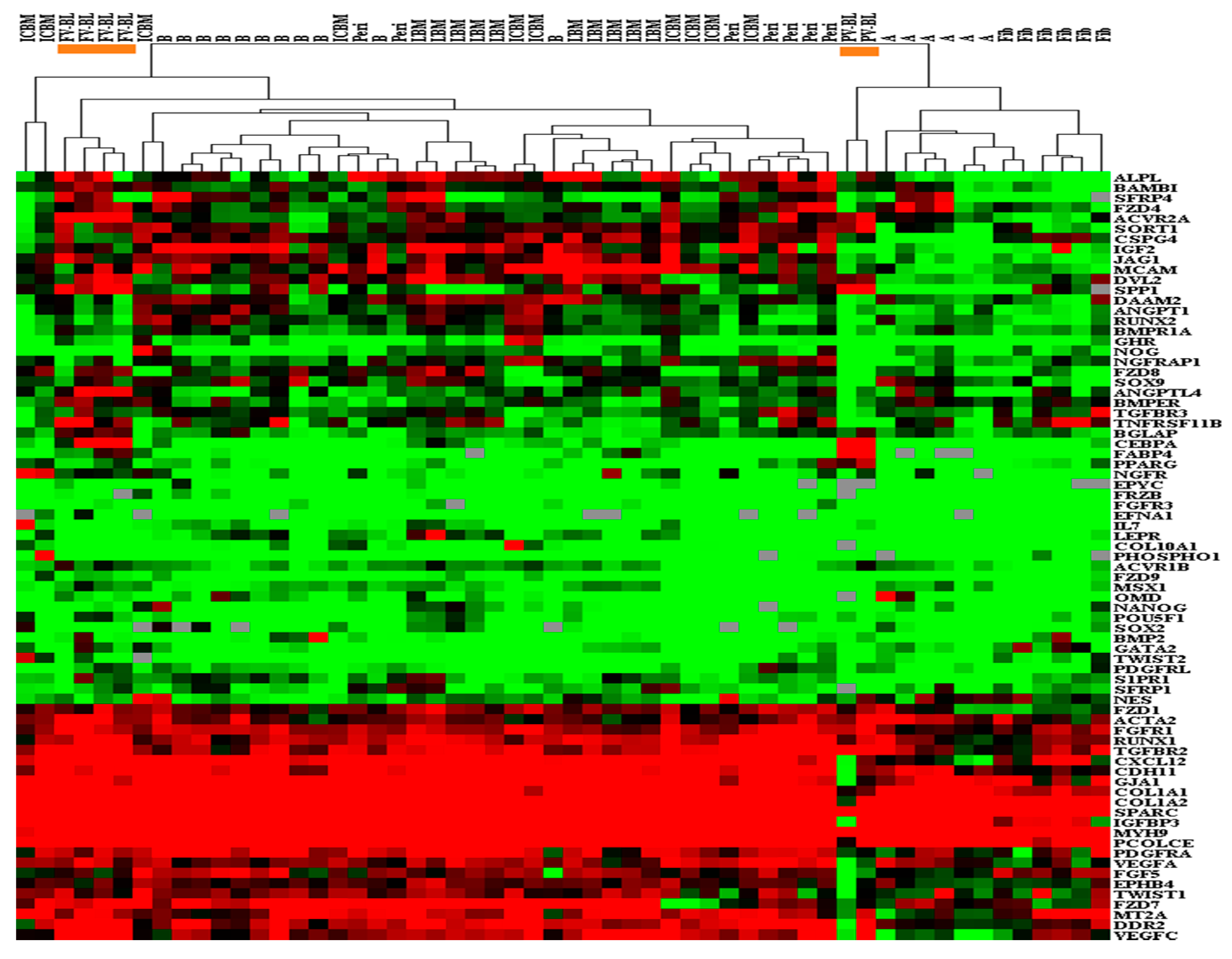
2.7. Hierarchical Clustering
2.8. Statistical Analysis
3. Results
3.1. Identification and Frequency of MSC-Like Colonies
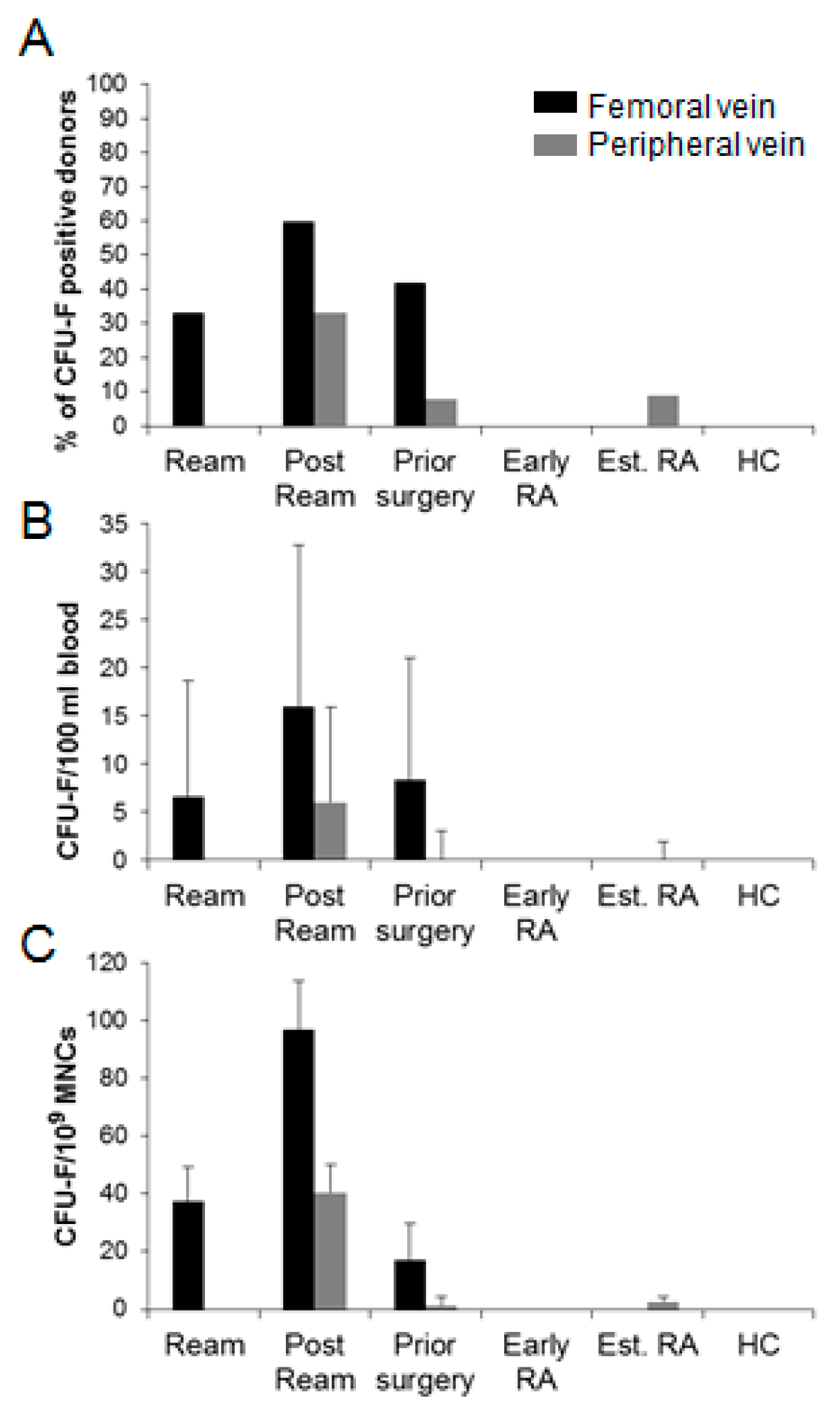
3.2. Circulatory CFU-Fs in Femoral and Peripheral Veins.
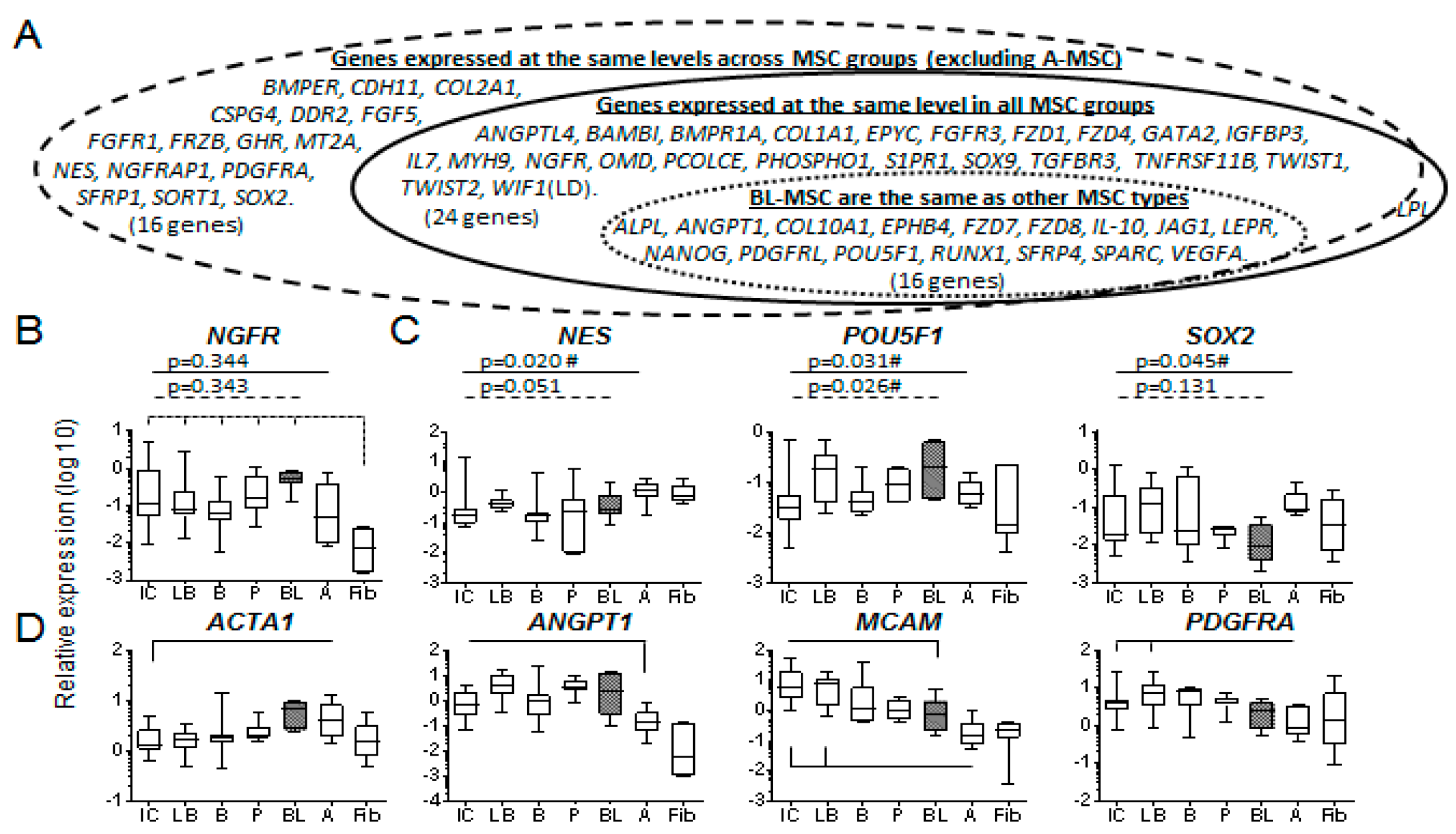
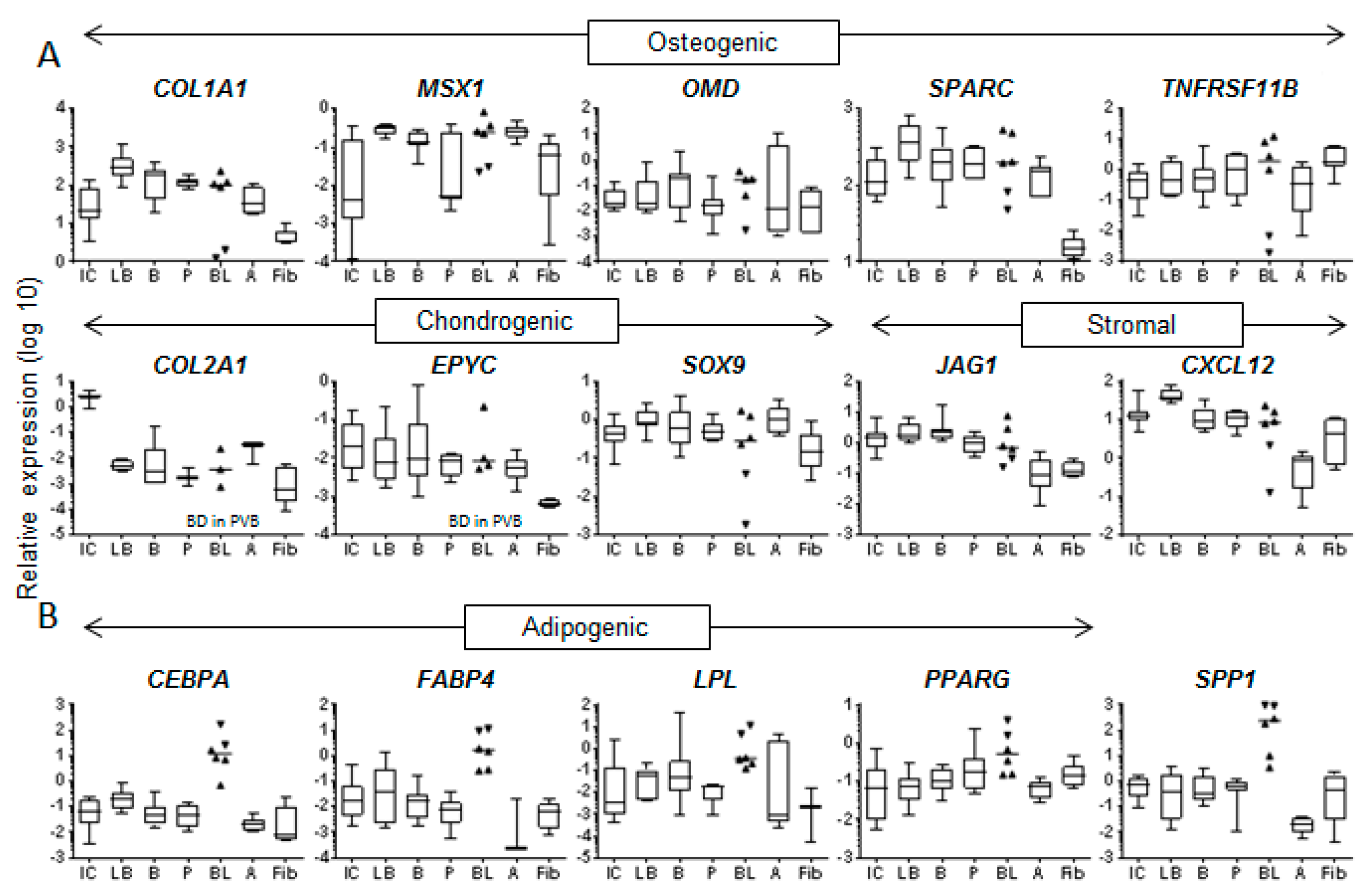
3.3. Molecular Profile of Blood-Derived CFU-Fs.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Ichinose, S.; Shinomiya, K.; Muneta, T. Suspended cells from trabecular bone by collagenase digestion become virtually identical to mesenchymal stem cells obtained from marrow aspirates. Blood 2004, 104, 2728–2735. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Zannettino, A.C.W.; Hay, S.J.; Shi, S.T.; Graves, S.E.; Kortesidis, A.; Simmons, P.J. Molecular and cellular characterisation of highly purified stromal stem cells derived from human bone marrow. J. Cell Sci. 2003, 116, 1827–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.; Crawford, A.; English, A.; Henshaw, K.; Mundy, J.; Corscadden, D.; McGonagle, D.; Chapman, T.; Emery, P.; Hatton, P. Synovial fluid mesenchymal stem cells in health and early osteoarthritis: Detection and functional evaluation at the single-cell level. Arthritis Rheum. 2008, 58, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- Wickham, M.Q.; Erickson, G.R.; Gimble, J.M.; Vail, T.P.; Guilak, F. Multipotent stromal cells derived from the infrapatellar fat pad of the knee. Clin. Orthop. Relat. Res. 2003, 412, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Dowthwaite, G.P.; Bishop, J.C.; Redman, S.N.; Khan, I.M.; Rooney, P.; Evans, D.J.R.; Wolfe, M.S.; Haughton, L.; Bayram, Z.; Boyer, S.; et al. The surface of articular cartilage contains a progenitor cell population. J. Cell Sci. 2004, 117, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Hedrick, M.H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Tempfer, H.; Wagner, A.; Gehwolf, R.; Lehner, C.; Tauber, M.; Resch, H.; Bauer, H.C. Perivascular cells of the supraspinatus tendon express both tendon-and stem cell-related markers. Histochem. Cell Biol. 2009, 131, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Kouroupis, D.; Churchman, S.M.; English, A.; Emery, P.; Giannoudis, P.V.; McGonagle, D.; Jones, E.A. Assessment of umbilical cord tissue as a source of mesenchymal stem cell/endothelial cell mixtures for bone regeneration. Regen. Med. 2013, 8, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Campagnoli, C.; Roberts, I.A.G.; Kumar, S.; Bennett, P.R.; Bellantuono, I.; Fisk, N.M. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 2001, 98, 2396–2402. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.; McGonagle, D. Human bone marrow mesenchymal stem cells in vivo. Rheumatology 2008, 47, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. Adult Mesenchymal Stem Cells: When, Where, and How. Stem Cells Int. 2015, 2015, 628767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraglia, A.; Cancedda, R.; Quarto, R. Clonal mesenchymal progenitors from human bone marrow differentiate in vitro according to a hierarchical model. J. Cell Sci. 2000, 113, 1161–1166. [Google Scholar]
- Kuznetsov, S.A.; Mankani, M.H.; Gronthos, S.; Satomura, K.; Bianco, P.; Robey, P.G. Circulating skeletal stem cells. J. Cell Biol. 2001, 153, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Rochefort, G.Y.; Delorme, B.; Lopez, A.; Herault, O.; Bonnet, P.; Charbord, P.; Domenech, J.; Eder, V. Multipotential mesenchymal stem cells are mobilized into peripheral blood by hypoxia. Stem Cells 2006, 24, 2202–2208. [Google Scholar] [CrossRef]
- Liu, L.; Yu, Q.; Hu, K.; Wang, B.; Zhang, Y.; Xu, Y.; Huang, H.; Fu, S.; Yu, X. Electro-Acupuncture Promotes Endogenous Multipotential Mesenchymal Stem Cell Mobilization into the Peripheral Blood. Cell. Physiol. Biochem. 2016, 38, 1605–1617. [Google Scholar] [CrossRef]
- Pitchford, S.C.; Furze, R.C.; Jones, C.P.; Wengner, A.M.; Rankin, S.M. Differential mobilization of subsets of progenitor cells from the bone marrow. Cell Stem Cell 2009, 4, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Tondreau, T.; Meuleman, N.; Delforge, A.; Dejeneffe, M.; Leroy, R.; Massy, M.; Lagneaux, L.; Mortier, C.; Bron, D. Mesenchymal stem cells derived from CD133-positive cells in mobilized peripheral blood and cord blood: Proliferation, Oct4 expression, and plasticity. Stem Cells 2005, 23, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, K.; Vasanji, A.; Drazba, J.A.; Butler, R.S.; Muschler, G.F. Circulating cells with osteogenic potential are physiologically mobilized into the fracture healing site in the parabiotic mice model. J. Orthop. Res. 2008, 26, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.; Seo, E.Y.; Chen, J.Y.; Lo, D.; McArdle, A.; Sinha, R.; Lu, W.J.; Tevlin, R.; Seita, J.; Chuang, M.T.; et al. Identification and specification of the mouse skeletal stem cell. Cell 2015, 160, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eseonu, O.I.; De Bari, C. Homing of mesenchymal stem cells: Mechanistic or stochastic? Implications for targeted delivery in arthritis. Rheumatology 2015, 54, 210–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogduijn, M.J.; Verstegen, M.M.; Engela, A.U.; Korevaar, S.S.; Roemeling-van Rhijn, M.; Merino, A.; Betjes, M.G.; Franquesa, M.; de Jonge, J.; Baan, C.C.; et al. No evidence for circulating mesenchymal stem cells in patients with organ injury. Stem Cells Dev. 2014, 23, 2328–2335. [Google Scholar] [CrossRef] [PubMed]
- Alm, J.J.; Koivu, H.M.; Heino, T.J.; Hentunen, T.A.; Laitinen, S.; Aro, H.T. Circulating plastic adherent mesenchymal stem cells in aged hip fracture patients. J. Orthop. Res. 2010, 28, 1634–1642. [Google Scholar] [CrossRef]
- Kuznetsov, S.A.; Mankani, M.H.; Leet, A.I.; Ziran, N.; Gronthos, S.; Robey, P.G. Circulating connective tissue precursors: Extreme rarity in humans and chondrogenic potential in guinea pigs. Stem Cells 2007, 25, 1830–1839. [Google Scholar] [CrossRef]
- Mansilla, E.; Marin, G.H.; Drago, H.; Sturla, F.; Salas, E.; Gardiner, C.; Gil, M.A.; Bossi, S.; Lamonega, R.; Ibar, R.; et al. Bloodstream cells phenotypically identical to human mesenchymal bone marrow stem cells circulate in large amounts under the influence of acute large skin damage: New evidence for their use in regenerative medicine. Transplant. Proc. 2006, 38, 967–969. [Google Scholar] [CrossRef]
- Tan, H.B.; Giannoudis, P.V.; Boxall, S.A.; McGonagle, D.; Jones, E. The systemic influence of platelet-derived growth factors on bone marrow mesenchymal stem cells in fracture patients. BMC Med. 2015, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Lefevre, S.; Knedla, A.; Tennie, C.; Kampmann, A.; Wunrau, C.; Dinser, R.; Evans, C.H.; Korb, A.; Schnäker, E.-M.; Tarner, I.H.; et al. Synovial fibroblasts spread rheumatoid arthritis to unaffected joints. Nat. Med. 2009, 15, 1414. [Google Scholar] [CrossRef] [Green Version]
- Marinova-Mutafchieva, L.; Williams, R.O.; Funa, K.; Maini, R.N.; Zvaifler, N.J. Inflammation is preceded by tumor necrosis factor-dependent infiltration of mesenchymal cells in experimental arthritis. Arthritis Rheum. 2002, 46, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Zvaifler, N.J.; Marinova-Mutafchieva, L.; Adams, G.; Edwards, C.J.; Moss, J.; Burger, J.A.; Maini, R.N. Mesenchymal precursor cells in the blood of normal individuals. Arthritis Res. 2000, 2, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinova-Mutafchieva, L.; Taylor, P.; Funa, K.; Maini, R.N.; Zvaifler, N.J. Mesenchymal cells expressing bone morphogenetic protein receptors are present in the rheumatoid arthritis joint. Arthritis Rheum. 2000, 43, 2046–2055. [Google Scholar] [CrossRef]
- Bucala, R.; Spiegel, L.A.; Chesney, J.; Hogan, M.; Cerami, A. Circulating fibrocytes define a new leukocyte subpopulation that mediates tissue repair. Mol. Med. 1994, 1, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosla, S.; Eghbali-Fatourechi, G.Z. Circulating cells with osteogenic potential. Ann. N. Y. Acad. Sci. 2006, 1068, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Riseborough, E.J.; Herndon, J.H. Alterations in pulmonary function, coagulation and fat metabolism in patients with fractures of the lower limbs. Clin. Orthop. Relat. Res. 1976, 115, 248–267. [Google Scholar] [CrossRef]
- Jones, E.A.; English, A.; Kinsey, S.E.; Straszynski, L.; Emery, P.; Ponchel, F.; McGonagle, D. Optimization of a flow cytometry-based protocol for detection and phenotypic characterization of multipotent mesenchymal stromal cells from human bone marrow. Cytom. Part B Clin. Cytom. 2006, 70, 391–399. [Google Scholar] [CrossRef]
- Giannoudis, P.V.; Tzioupis, C.; Green, J. Surgical techniques: How I do it? The Reamer/Irrigator/Aspirator (RIA) System. Inj. Int. J. Care Inj. 2009, 40, 1231–1236. [Google Scholar] [CrossRef]
- Churchman, S.M.; Ponchel, F.; Boxall, S.A.; Cuthbert, R.; Kouroupis, D.; Roshdy, T.; Jones, E.A.; Giannoudis, P.V.; Emery, P.; McGonagle, D. Transcriptional profile of native CD271+ mesenchymal stem cells: Evidence for multiple fates, with prominent osteogenic and Wnt pathway signaling activity. Arthritis Rheum. 2012, 64, 2632–2643. [Google Scholar] [CrossRef]
- De Hoon, M.J.; Imoto, S.; Nolan, J.; Miyano, S. Open source clustering software. Bioinformatics 2004, 20, 1453–1454. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, A.J. Java Treeview—Extensible visualization of microarray data. Bioinformatics 2004, 20, 3246–3248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012, 21, 2724–2752. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.; Riminucci, M.; Gronthos, S.; Robey, P.G. Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells 2001, 19, 180–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomiyama, T.; Perez-Tilve, D.; Ogawa, D.; Gizard, F.; Zhao, Y.; Heywood, E.B.; Bruemmer, D.; Jones, K.L.; Kawamori, R.; Cassis, L.A.; et al. Osteopontin mediates obesity-induced adipose tissue macrophage infiltration and insulin resistance in mice. J. Clin. Investig. 2007, 117, 2877–2888. [Google Scholar] [CrossRef]
- Risau, W.; Flamme, I. Vasculogenesis. Annu. Rev. Cell Dev. Biol. 1995, 11, 73–91. [Google Scholar] [CrossRef]
- Wiegner, R.; Rudhart, N.E.; Barth, E.; Gebhard, F.; Lampl, L.; Huber-Lang, M.S.; Brenner, R.E. Mesenchymal stem cells in peripheral blood of severely injured patients. Eur. J. Trauma Emerg. Surg. 2018, 44, 627–636. [Google Scholar] [CrossRef]
- Di Bernardo, G.; Messina, G.; Capasso, S.; Del Gaudio, S.; Cipollaro, M.; Peluso, G.; Galderisi, U.; Casale, F.; Monda, M. Sera of overweight people promote in vitro adipocyte differentiation of bone marrow stromal cells. Stem Cell Res. Ther. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Blazsek, I.; Chagraoui, J.; Peault, B. Ontogenic emergence of the hematon, a morphogenetic stromal unit that supports multipotential hematopoietic progenitors in mouse bone marrow. Blood 2000, 96, 3763–3771. [Google Scholar] [CrossRef]
- Soukas, A.; Socci, N.D.; Saatkamp, B.D.; Novelli, S.; Friedman, J.M. Distinct Transcriptional Profiles of Adipogenesisin Vivo and in vitro. J. Biol. Chem. 2001, 276, 34167–34174. [Google Scholar] [CrossRef] [Green Version]
- Husebye, E.E.; Lyberg, T.; Madsen, J.E.; Eriksen, M.; Roise, O. The influence of a one-step reamer-irrigator-aspirator technique on the intramedullary pressure in the pig femur. Injury 2006, 37, 935–940. [Google Scholar] [CrossRef] [Green Version]
- Husebye, E.E.; Lyberg, T.; Opdahl, H.; Roise, O. Intravasation of bone marrow content. Can its magnitude and effects be modulated by low pressure reaming in a porcine model? Injury 2010, 41 (Suppl. 2), S9–S15. [Google Scholar] [CrossRef]
- Ganong, R.B. Fat emboli syndrome in isolated fractures of the tibia and femur. Clin. Orthop. Relat. Res. 1993, 291, 208–214. [Google Scholar] [CrossRef]
- Mahajan, M.M.; Cheng, B.; Beyer, A.I.; Mulvaney, U.S.; Wilkinson, M.B.; Fomin, M.E.; Muench, M.O. A quantitative assessment of the content of hematopoietic stem cells in mouse and human endosteal-bone marrow: A simple and rapid method for the isolation of mouse central bone marrow. BMC Hematol. 2015, 15, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Surgery/Sampling Time | Bloods | Total Blood Volumes (mL) | n | F:M | Median Age (Range) |
|---|---|---|---|---|---|---|
| Reaming | Patients undergoing reaming | FV+PV | 510 + 510 | 24 | 10:14 | 44.5 (18–66) |
| a) Sequential conventional reaming for IM nailing of closed tibial shaft fracture. | FV+PV | 360 + 360 | 9 | 3:6 | 40 (18–53) | |
| b) All fracture non-union patients requiring RIA for bone graft acquisition. | FV+PV | 150 + 150 | 15 | 7:8 | 45 (19–66) | |
| Prior surgery | Current surgery not relevant #, samples taken before surgical intervention | FV+PV | 120 + 120 | 12 | 5:7 | 40 (18–67) |
| Early RA | N/A | PV | 168 | 15 | 11:4 | 51 (23–77) |
| Established RA | N/A | PV | 168 | 11 | 7:4 | 47 (32–66) |
| HC | N/A | PV | 120 | 12 | 4:8 | 33.5 (24–54) |
| OVERALL | FV + PV | 630 + 1086 | 74 | 37:37 | 43.5 (18–77) |
| ICBM-MSCs | LBM-MSCs | CB-MSCs | Peri-MSCs | A-MSCs | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Comparator | BL-MSCs | FVB- MSCs | BL-MSCs | FVB- MSCs | BL-MSCs | FVB- MSCs | BL-MSCs | FVB- MSCs | BL-MSCs | FVB-MSCs |
| ACTA2 | 0.028 | |||||||||
| ACVR1B | 0.005 | 0.009 | ||||||||
| ACVR2A | 0.010 | 0.032 | <0.001 | 0.002 | ||||||
| BGLAP | 0.006 | 0.008 | 0.010 | 0.014 | ||||||
| BMP2 | 0.012 | 0.042 | ||||||||
| CEBPA | 0.027 | <0.001 | 0.002 | 0.018 | 0.001 | 0.004 | ||||
| COL1A2 | 0.021 | |||||||||
| CXCL12 | 0.013 | |||||||||
| DAAM2 | 0.016 | |||||||||
| DVL2 | 0.041 | 0.003 | 0.011 | |||||||
| EFNA1 | 0.040 | |||||||||
| FABP4 | 0.029 | 0.008 | 0.038 | 0.008 | 0.020 | |||||
| FZD9 | 0.038 | 0.008 | ||||||||
| GJA1 | 0.037 | 0.042 | ||||||||
| IGF2 | 0.048 | |||||||||
| MCAM | 0.038 | |||||||||
| MSX1 | 0.003 | 0.030 | 0.050 | |||||||
| NOG | 0.014 | |||||||||
| PPARG | 0.043 | 0.041 | ||||||||
| RUNX2 | 0.017 | |||||||||
| SORT1 | 0.006 | |||||||||
| SP7 | 0.008 | |||||||||
| SPP1 | 0.017 | 0.041 | <0.001 | 0.038 | ||||||
| TGFBR3 | 0.015 | <0.001 | 0.001 | |||||||
| Differences (BL-MSCs) | 8 | 9 | 9 | 5 | 8 | |||||
| ANGPTL4 | 0.012 | |||||||||
| BMPER | 0.021 | 0.031 | ||||||||
| DDR2 | 0.046 | <0.001 | ||||||||
| FGFR1 | 0.005 | |||||||||
| GHR | 0.012 | |||||||||
| PCOLCE | 0.027 | 0.005 | ||||||||
| S1PR1 | 0.033 | |||||||||
| TGFBR2 | 0.002 | 0.004 | ||||||||
| VEGFC | 0.012 | <0.000 | ||||||||
| Differences (FVB-MSCs) | 2 | 2 | 8 | 4 | 12 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churchman, S.M.; Jones, E.A.; Roshdy, T.; Cox, G.; Boxall, S.A.; McGonagle, D.; Giannoudis, P.V. Transient Existence of Circulating Mesenchymal Stem Cells in the Deep Veins in Humans Following Long Bone Intramedullary Reaming. J. Clin. Med. 2020, 9, 968. https://doi.org/10.3390/jcm9040968
Churchman SM, Jones EA, Roshdy T, Cox G, Boxall SA, McGonagle D, Giannoudis PV. Transient Existence of Circulating Mesenchymal Stem Cells in the Deep Veins in Humans Following Long Bone Intramedullary Reaming. Journal of Clinical Medicine. 2020; 9(4):968. https://doi.org/10.3390/jcm9040968
Chicago/Turabian StyleChurchman, Sarah M, Elena A Jones, Tarek Roshdy, George Cox, Sally A Boxall, Dennis McGonagle, and Peter V Giannoudis. 2020. "Transient Existence of Circulating Mesenchymal Stem Cells in the Deep Veins in Humans Following Long Bone Intramedullary Reaming" Journal of Clinical Medicine 9, no. 4: 968. https://doi.org/10.3390/jcm9040968