Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Measurement of Serum and Allergen-Specific IgE Levels
2.3. Plasma and Urine Sample Preparation
2.4. 1H–Nuclear Magnetic Resonance (NMR) Spectroscopy
2.5. NMR Data Processing and Analysis
2.6. Statistical Analysis
3. Results
3.1. Population Characteristics
3.2. Metabolites Sets Categorized by Asthma Outcome and Mite, Food, and IgE Sensitization
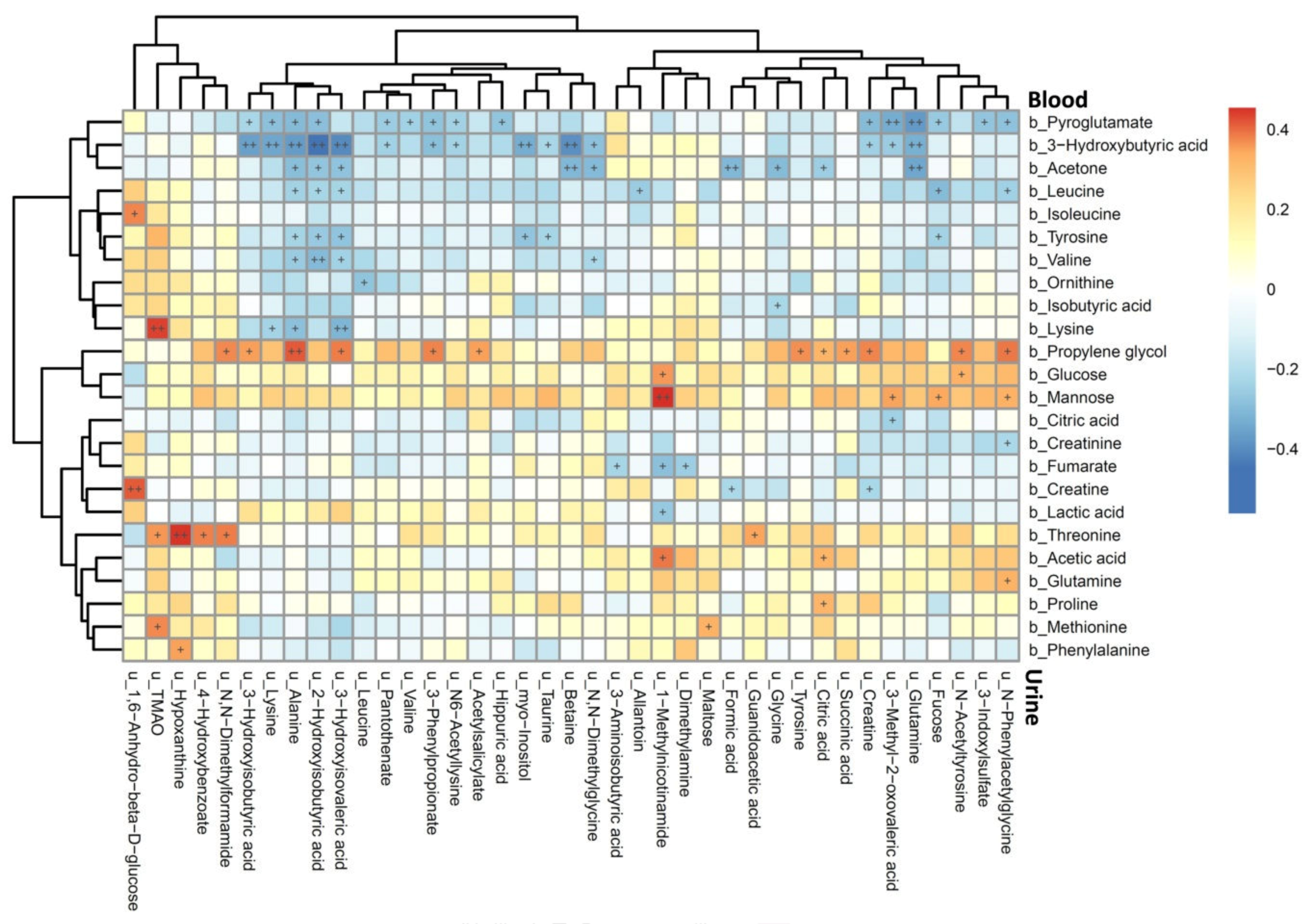
3.3. Association between Plasma and Urine Metabolites
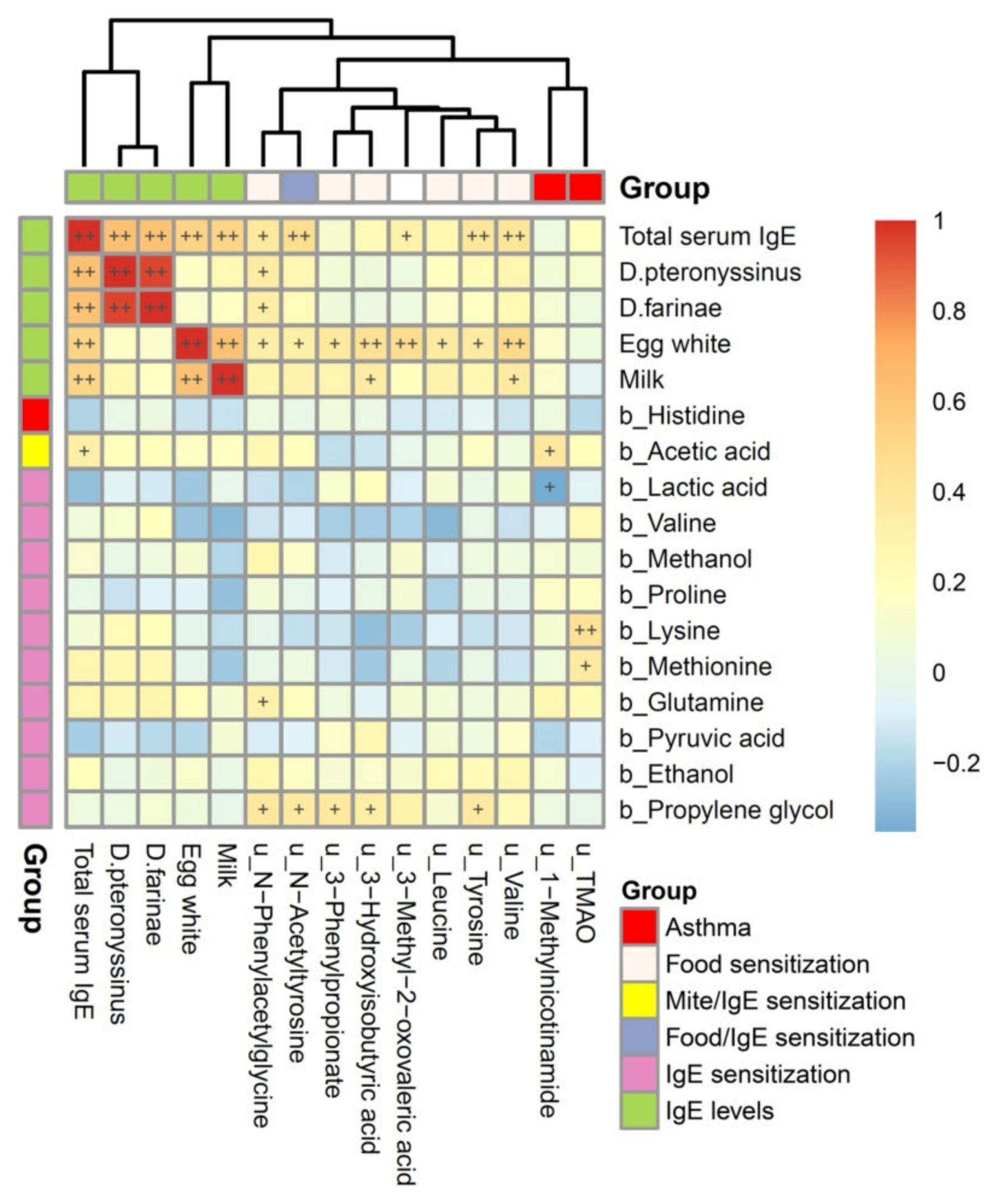
3.4. Association of Atopic Indices Associated Metabolites between Plasma and Urine Metabolome
3.5. Metabolites Associated with Atopic Indices Related to Allergen-Specific IgE Levels
3.6. Metabolic Pathway and Functional Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kauffmann, F.; Demenais, F. Gene-environment interactions in asthma and allergic diseases: Challenges and perspectives. J. Allergy Clin. Immunol. 2012, 130, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Burbank, A.J.; Sood, A.K.; Kesic, M.J.; Peden, D.B.; Hernandez, M.L. Environmental determinants of allergy and asthma in early life. J. Allergy Clin. Immunol. 2017, 140, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pite, H.; Morais-Almeida, M.; Rocha, S.M. Metabolomics in asthma: Where do we stand? Curr. Opin. Pulm. Med. 2018, 24, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Kim, S.H.; Lee, H.S.; Choi, G.S.; Jung, Y.S.; Ryu, D.H.; Park, H.S.; Hwang, G.S. Serum metabolomics reveals pathways and biomarkers associated with asthma pathogenesis. Clin. Exp. Allergy 2013, 43, 425–433. [Google Scholar] [CrossRef]
- Mattarucchi, E.; Baraldi, E.; Guillou, C. Metabolomics applied to urine samples in childhood asthma; differentiation between asthma phenotypes and identification of relevant metabolites. Biomed. Chromatogr. 2012, 26, 89–94. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chan, Y.L.; Tsai, Y.S.; Chen, S.A.; Wang, C.J.; Chen, K.F.; Chung, I.F. Airway Microbial Diversity is Inversely Associated with Mite-Sensitized Rhinitis and Asthma in Early Childhood. Sci. Rep. 2017, 7, 1820. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Huang, S.Y.; Peng, Y.C.; Tsai, M.H.; Hua, M.C.; Yao, T.C.; Yeh, K.W.; Huang, J.L. Maternal vitamin D levels are inversely related to allergic sensitization and atopic diseases in early childhood. Pediatr. Allergy Immunol. 2015, 26, 337–343. [Google Scholar] [CrossRef]
- Lee, A.J.; Thalayasingam, M.; Lee, B.W. Food allergy in Asia: How does it compare? Asia Pac. Allergy 2013, 3, 3–14. [Google Scholar] [CrossRef]
- Ballardini, N.; Nilsson, C.; Nilsson, M.; Lilja, G. ImmunoCAP Phadiatop Infant—A new blood test for detecting IgE sensitisation in children at 2 years of age. Allergy 2006, 61, 337–343. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Yeh, K.W.; Lin, G.; Chiang, M.H.; Yang, S.C.; Chao, W.J.; Yao, T.C.; Tsai, M.H.; Hua, M.C.; Liao, S.L.; et al. Metabolomics Reveals Dynamic Metabolic Changes Associated with Age in Early Childhood. PLoS ONE 2016, 11, e0149823. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Lin, G.; Cheng, M.L.; Chiang, M.H.; Tsai, M.H.; Lai, S.H.; Wong, K.S.; Hsieh, S.Y. Metabolomic Profiling of Infectious Parapneumonic Effusions Reveals Biomarkers for Guiding Management of Children with Streptococcus pneumoniae Pneumonia. Sci. Rep. 2016, 6, 24930. [Google Scholar] [CrossRef]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Meyer, T.; Sinnaeve, D.; Van Gasse, B.; Tsiporkova, E.; Rietzschel, E.R.; De Buyzere, M.L.; Gillebert, T.C.; Bekaert, S.; Martins, J.C.; Van Criekinge, W. NMR-based characterization of metabolic alterations in hypertension using an adaptive, intelligent binning algorithm. Anal. Chem. 2008, 80, 3783–3790. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.J.; van Duijnhoven, J.P.M.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [Green Version]
- Barrios, C.; Spector, T.D.; Menni, C. Blood, urine and faecal metabolite profiles in the study of adult renal disease. Arch. Biochem. Biophys. 2016, 589, 81–92. [Google Scholar] [CrossRef]
- Manoli, I.; Venditti, C.P. Disorders of branched chain amino acid metabolism. Transl. Sci. Rare Dis. 2016, 1, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Leitner, M.; Fragner, L.; Danner, S.; Holeschofsky, N.; Leitner, K.; Tischler, S.; Doerfler, H.; Bachmann, G.; Sun, X.; Jaeger, W.; et al. Combined Metabolomic Analysis of Plasma and Urine Reveals AHBA, Tryptophan and Serotonin Metabolism as Potential Risk Factors in Gestational Diabetes Mellitus (GDM). Front. Mol. Biosci. 2017, 4, 84. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Chan, Y.L.; Tsai, M.H.; Wang, C.J.; Chiang, M.H.; Chiu, C.C. Gut microbial dysbiosis is associated with allergen-specific IgE responses in young children with airway allergies. World Allergy Organ. J. 2019, 12, 100021. [Google Scholar] [CrossRef] [Green Version]
- Guasch-Ferre, M.; Bhupathiraju, S.N.; Hu, F.B. Use of Metabolomics in Improving Assessment of Dietary Intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, C.C.; Duarte, I.F.; Gomes, J.; Carrola, J.; Barros, A.S.; Gil, A.M.; Bousquet, J.; Bom, A.T.; Rocha, S.M. Urinary metabolomic changes as a predictive biomarker of asthma exacerbation. J. Allergy Clin. Immunol. 2014, 133, 261–263. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Gogg, S.; Lotta, L.A.; Stancakova, A.; Nerstedt, A.; Boren, J.; Bluher, M.; Ferrannini, E.; Langenberg, C.; Wareham, N.J.; et al. Elevated Plasma Levels of 3-Hydroxyisobutyric Acid Are Associated With Incident Type 2 Diabetes. EBioMedicine 2018, 27, 151–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schadewaldt, P.; Hammen, H.W.; Ott, A.C.; Wendel, U. Renal clearance of branched-chain L-amino and 2-oxo acids in maple syrup urine disease. J. Inherit. Metab. Dis. 1999, 22, 706–722. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Lin, G.; Cheng, M.L.; Chiang, M.H.; Tsai, M.H.; Su, K.W.; Hua, M.C.; Liao, S.L.; Lai, S.H.; Yao, T.C.; et al. Longitudinal urinary metabolomic profiling reveals metabolites for asthma development in early childhood. Pediatr. Allergy Immunol. 2018, 29, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Bjorksten, B.; Engstrand, L.; Jenmalm, M.C. Low gut microbiota diversity in early infancy precedes asthma at school age. Clin. Exp. Allergy 2014, 44, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.S.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsu, H. Pathophysiologic role of histamine: Evidence clarified by histidine decarboxylase gene knockout mice. Int. Arch. Allergy Immunol. 2012, 158, 2–6. [Google Scholar] [CrossRef]
- Staley, K.G.; Stover, C.; Strippoli, M.P.; Spycher, B.D.; Silverman, M.; Kuehni, C.E. Mannan-binding lectin in young children with asthma differs by level of severity. J. Allergy Clin. Immunol. 2007, 119, 503–505. [Google Scholar] [CrossRef]
- Biedroń, R.; Ciszek, M.; Tokarczyk, M.; Bobek, M.; Kurnyta, M.; Słominska, E.M.; Smoleński, R.T.; Marcinkiewicz, J. 1-Methylnicotinamide and nicotinamide: Two related anti-inflammatory agents that differentially affect the functions of activated macrophages. Arch. Immunol. Ther. Exp. 2008, 56, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Motta, A.; Paris, D.; D’Amato, M.; Melck, D.; Calabrese, C.; Vitale, C.; Stanziola, A.A.; Corso, G.; Sofia, M.; Maniscalco, M. NMR metabolomic analysis of exhaled breath condensate of asthmatic patients at two different temperatures. J. Proteome Res. 2014, 13, 6107–6120. [Google Scholar] [CrossRef] [PubMed]
- Saude, E.J.; Skappak, C.D.; Regush, S.; Cook, K.; Ben-Zvi, A.; Becker, A.; Moqbel, R.; Sykes, B.D.; Rowe, B.H.; Adamko, D.J. Metabolomic profiling of asthma: Diagnostic utility of urine nuclear magnetic resonance spectroscopy. J. Allergy Clin. Immunol. 2011, 127, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.S.; Dahlin, A.; McGeachie, M.J.; Qiu, W.; Sordillo, J.; Wan, E.S.; Wu, A.C.; Lasky-Su, J. Asthma Metabolomics and the Potential for Integrative Omics in Research and the Clinic. Chest 2017, 151, 262–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Characteristics | Asthmatics (n = 28) | Controls (n = 26) | p Value |
|---|---|---|---|
| Age (yr) | 3.62 ± 0.66 | 3.61 ± 0.73 | 0.971 |
| Sex, male | 19 (67.9%) | 16 (61.5%) | 0.627 |
| Maternal atopy | 13 (48.1%) | 9 (34.6%) | 0.318 |
| Passive smoking | 9 (32.1%) | 10 (38.5) | 0.627 |
| Household income | 0.559 | ||
| Low, ≤ 500,000 NTD | 10 (37.0%) | 12 (46.2%) | |
| Medium, 500,000–1,000,000 NTD | 11 (40.7%) | 11 (42.3%) | |
| High, > 1,000,000 NTD | 6 (22.2%) | 3 (11.5%) | |
| Allergen-specific IgE, kU/L | |||
| D. pteronyssinus | 29.67 ± 53.07 | 0.34 ± 0.55 | 0.001 |
| D. farinae | 14.41 ± 28.04 | 0.17 ± 0.26 | 0.001 |
| Egg white | 0.63 ± 0.83 | 0.33 ± 0.52 | 0.203 |
| Cow’s milk | 0.58 ± 0.90 | 0.22 ± 0.27 | 0.052 |
| Sensitization | |||
| Mite | 19 (67.9%) | 5 (19.2%) | <0.001 |
| Food | 15 (53.6%) | 7 (26.9%) | 0.046 |
| IgE > 100 kU/L | 14 (50.0%) | 2 (7.7%) | 0.001 |
| Total serum IgE, kU/L | 224.00 ± 292.43 | 47.79 ± 60.67 | 0.001 |
| Asthma | Mite Sensitization | Food Sensitization | IgE Sensitization | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolites | Chemical Shift, ppm | VIP Score * | Fold Change † | p ‡ | VIP Score | Fold Change | p | VIP Score | Fold Change | p | VIP Score | Fold Change | p |
| Plasma | |||||||||||||
| Histidine | 7.030–7.078 (s) | 1.28 | 1.13 | 0.009 | 0.83 | 1.08 | 0.081 | 0.04 | 0.98 | 0.934 | 0.36 | 1.05 | 0.293 |
| Acetic acid | 1.850–1.930 (d) | 1.19 | 1.41 | 0.226 | 2.02 | 1.37 | 0.031 | 1.04 | 1.24 | 0.254 | 1.68 | 1.48 | 0.009 |
| Lactic acid | 1.304–1.340 (d) | 1.02 | 0.86 | 0.304 | 0.40 | 0.94 | 0.678 | 1.38 | 0.80 | 0.125 | 1.41 | 0.63 | 0.030 |
| Valine | 3.588–3.615 (d) | 0.45 | 1.04 | 0.309 | 0.69 | 1.06 | 0.097 | 0.12 | 1.01 | 0.768 | 0.64 | 1.10 | 0.027 |
| Methanol | 3.347–3.363 (s) | 0.82 | 1.13 | 0.241 | 0.27 | 1.04 | 0.686 | 0.45 | 1.07 | 0.479 | 1.19 | 1.33 | 0.009 |
| Proline | 3.301–3.347 (dt) | 0.41 | 1.04 | 0.449 | 0.44 | 1.03 | 0.397 | 0.18 | 0.98 | 0.720 | 0.90 | 1.15 | 0.011 |
| Lysine | 2.999–3.024 (t) | 0.02 | 1.00 | 0.947 | 0.36 | 1.02 | 0.291 | 0.12 | 0.99 | 0.722 | 0.38 | 1.04 | 0.036 |
| Methionine | 2.610–2.653 (t) | 0.48 | 1.05 | 0.330 | 0.63 | 1.06 | 0.186 | 0.03 | 1.01 | 0.941 | 1.00 | 1.16 | 0.002 |
| Glutamine | 2.416–2.494 (m) | 0.15 | 1.02 | 0.775 | 0.23 | 1.02 | 0.633 | 0.61 | 1.06 | 0.185 | 0.68 | 1.11 | 0.041 |
| Pyruvic acid | 2.354–2.376 (s) | 0.87 | 0.86 | 0.352 | 0.79 | 0.87 | 0.379 | 0.80 | 0.89 | 0.351 | 1.27 | 0.66 | 0.025 |
| Ethanol | 1.148–1.181 (t) | 1.86 | 1.69 | 0.062 | 0.44 | 1.05 | 0.652 | 0.82 | 0.95 | 0.383 | 1.50 | 1.64 | 0.026 |
| Propylene glycol | 1.123–1.148 (d) | 1.52 | 1.80 | 0.241 | 0.37 | 1.26 | 0.766 | 0.80 | 1.47 | 0.501 | 2.20 | 2.65 | 0.009 |
| Urine | |||||||||||||
| 1-Methylnicotinamide | 8.874–8.906 (d) | 2.44 | 0.64 | 0.011 | 0.21 | 1.00 | 0.818 | 0.31 | 1.32 | 0.737 | 1.04 | 0.80 | 0.289 |
| TMAO | 3.256–3.261 (s) | 1.74 | 0.75 | 0.025 | 0.80 | 1.15 | 0.274 | 0.01 | 1.04 | 0.992 | 1.10 | 1.29 | 0.163 |
| N-Phenylacetylglycine | 7.330–7.370 (m) | 0.05 | 0.99 | 0.946 | 1.01 | 1.20 | 0.151 | 1.39 | 1.36 | 0.024 | 0.93 | 1.22 | 0.224 |
| Tyrosine | 6.874–6.898 (m) | 0.11 | 0.98 | 0.871 | 0.46 | 1.07 | 0.454 | 1.45 | 1.31 | 0.006 | 0.93 | 1.23 | 0.158 |
| 3-Phenylpropionate | 2.870–2.907 (t) | 0.23 | 0.97 | 0.691 | 0.35 | 1.03 | 0.519 | 1.13 | 1.20 | 0.016 | 0.13 | 1.02 | 0.828 |
| 3-Methyl-2-oxovaleric acid | 1.083–1.105 (d) | 0.25 | 0.98 | 0.725 | 0.26 | 1.03 | 0.692 | 1.68 | 1.40 | 0.003 | 0.28 | 1.07 | 0.693 |
| 3-Hydroxyisobutyric acid | 1.050–1.070 (d) | 0.03 | 1.24 | 0.974 | 0.88 | 1.40 | 0.231 | 1.48 | 1.74 | 0.022 | 1.03 | 1.72 | 0.196 |
| Valine | 1.023–1.041 (d) | 0.18 | 0.99 | 0.772 | 0.80 | 1.12 | 0.164 | 1.64 | 1.35 | 0.001 | 0.37 | 1.07 | 0.555 |
| Leucine | 0.936–0.966 (t) | 0.41 | 0.94 | 0.490 | 0.81 | 1.11 | 0.132 | 0.97 | 1.16 | 0.042 | 0.06 | 1.00 | 0.915 |
| N-Acetyltyrosine | 6.841–6.869 (d) | 0.38 | 1.02 | 0.568 | 0.00 | 0.97 | 1.000 | 1.16 | 1.24 | 0.030 | 1.37 | 1.25 | 0.034 |
| Sample | Metabolites | Pathway Name | Total | Hits | Raw p | FDR | Function |
|---|---|---|---|---|---|---|---|
| Plasma | Histidine, Glutamine, Methionine, Valine, Lysine, Proline | Aminoacyl-tRNA biosynthesis | 75 | 6 | 5.97E-07 | 4.77E-05 | Genetic Information Processing, Translation |
| Ethanol, Pyruvic acid, Lactic acid, Acetic acid | Glycolysis or Gluconeogenesis | 31 | 4 | 1.04E-05 | 3.16E-04 | Carbohydrate metabolism | |
| Pyruvic acid, Propylene glycol, Lactic acid, Acetic acid | Pyruvate metabolism | 32 | 4 | 1.18E-05 | 3.16E-04 | Carbohydrate metabolism | |
| Urine | Leucine, Valine, 3-Hydroxyisobutyric acid | Valine, leucine and isoleucine degradation | 40 | 3 | 4.71E-04 | 2.68E-02 | Amino acid metabolism |
| N-Phenylacetylglycine, Tyrosine, 3-Phenylpropionate | Phenylalanine metabolism | 45 | 3 | 6.69E-04 | 2.68E-02 | Amino acid metabolism |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-Y.; Cheng, M.-L.; Chiang, M.-H.; Wang, C.-J.; Tsai, M.-H.; Lin, G. Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma. J. Clin. Med. 2020, 9, 887. https://doi.org/10.3390/jcm9030887
Chiu C-Y, Cheng M-L, Chiang M-H, Wang C-J, Tsai M-H, Lin G. Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma. Journal of Clinical Medicine. 2020; 9(3):887. https://doi.org/10.3390/jcm9030887
Chicago/Turabian StyleChiu, Chih-Yung, Mei-Ling Cheng, Meng-Han Chiang, Chia-Jung Wang, Ming-Han Tsai, and Gigin Lin. 2020. "Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma" Journal of Clinical Medicine 9, no. 3: 887. https://doi.org/10.3390/jcm9030887