Oncogenic Role of Secreted Engrailed Homeobox 2 (EN2) in Prostate Cancer

 ,
,  , ,
, , 

Abstract
1. Introduction
2. Material and Methods
2.1. Patients and Samples
2.2. Datasets Analysis
2.3. EN2 Protein, Reagents, and Cell Lines
2.4. RNA Isolation, Reverse-Transcription, and Real-Time Quantitative PCR (qPCR)
2.5. Measurements of Cell Proliferation
2.6. Measurements of Migration Capacity
2.7. Measurement of Free Cytosolic Calcium Concentration ([Ca2+]i)
2.8. Microarray of Gene Expression Profile
2.9. Western Blot
2.10. Determination of PSA and EN2 Levels by ELISA
2.11. Statistical Analysis
3. Results
3.1. EN2 Is Overexpressed in Tissue and Urine Samples from PCa Patients Compared with Controls
3.2. EN2 Is Overexpressed, Secreted, and Modulates Aggressiveness Features in PCa Cells
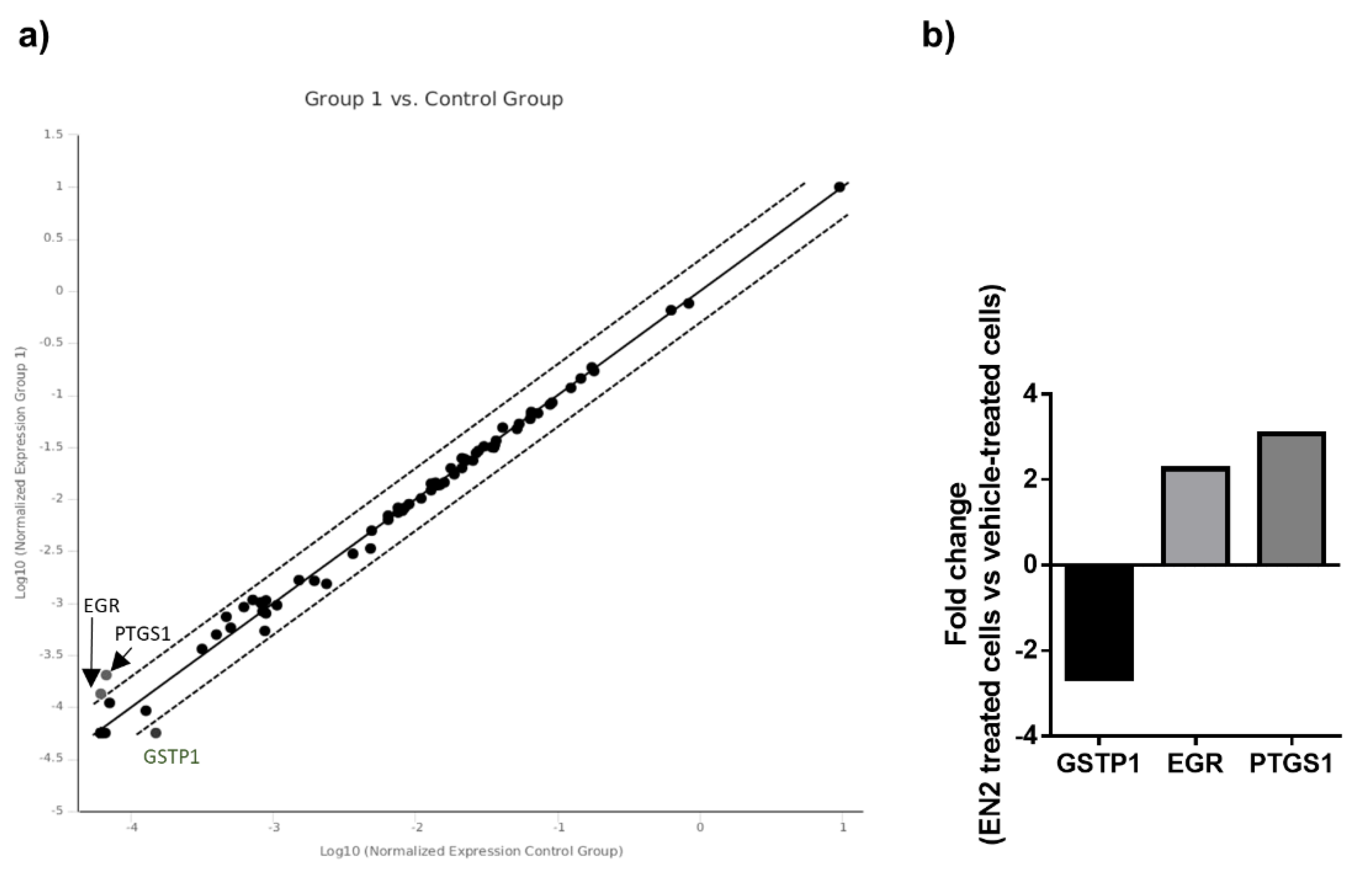
3.3. EN2 Modulates Key Signaling Pathways and Molecular Targets in PCa Cells
4. Discussion
Supplementary Materials
Author Contributions
Financial Support
Acknowledgments
Conflicts of Interest
References
- Center, M.M.; Jemal, A.; Lortet-Tieulent, J.; Ward, E.; Ferlay, J.; Brawley, O.; Bray, F. International variation in prostate cancer incidence and mortality rates. Eur. Urol. 2012, 61, 1079–1092. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Welch, H.G.; Gorski, D.H.; Albertsen, P.C. Trends in Metastatic Breast and Prostate Cancer--Lessons in Cancer Dynamics. N. Engl. J. Med. 2015, 373, 1685–1687. [Google Scholar] [CrossRef]
- Nadler, R.B.; Humphrey, P.A.; Smith, D.S.; Catalona, W.J.; Ratliff, T.L. Effect of inflammation and benign prostatic hyperplasia on elevated serum prostate specific antigen levels. J. Urol. 1995, 154, 407–413. [Google Scholar] [CrossRef]
- Shariat, S.F.; Karam, J.A.; Margulis, V.; Karakiewicz, P.I. New blood-based biomarkers for the diagnosis, staging and prognosis of prostate cancer. BJU Int. 2008, 101, 675–683. [Google Scholar] [CrossRef]
- Fenton, J.J.; Weyrich, M.S.; Durbin, S.; Liu, Y.; Bang, H.; Melnikow, J. Prostate-Specific Antigen-Based Screening for Prostate Cancer: Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2018, 319, 1914–1931. [Google Scholar] [CrossRef]
- Attard, G.; Parker, C.; Eeles, R.A.; Schröder, F.; Tomlins, S.A.; Tannock, I.; Drake, C.G.; de Bono, J.S. Prostate cancer. Lancet 2016, 387, 70–82. [Google Scholar] [CrossRef]
- Wedge, D.C.; Gundem, G.; Mitchell, T.; Woodcock, D.J.; Martincorena, I.; Ghori, M.; Zamora, J.; Butler, A.; Whitaker, H.; Kote-Jarai, Z.; et al. Sequencing of prostate cancers identifies new cancer genes, routes of progression and drug targets. Nat. Genet. 2018, 50, 682. [Google Scholar] [CrossRef]
- Shah, N.; Sukumar, S. The Hox genes and their roles in oncogenesis. Nat. Rev. Cancer 2010, 10, 361–371. [Google Scholar] [CrossRef]
- Logan, C.; Hanks, M.C.; Noble-Topham, S.; Nallainathan, D.; Provart, N.J.; Joyner, A.L. Cloning and sequence comparison of the mouse, human, and chicken engrailed genes reveal potential functional domains and regulatory regions. Dev. Genet. 1992, 13, 345–358. [Google Scholar] [CrossRef]
- Morata, G.; Lawrence, P.A. Control of compartment development by the engrailed gene in Drosophila. Nature 1975, 255, 614–617. [Google Scholar] [CrossRef] [PubMed]
- McGrath, S.E.; Michael, A.; Morgan, R.; Pandha, H. EN2: A novel prostate cancer biomarker. Biomark. Med 2013, 7, 893–901. [Google Scholar] [CrossRef]
- Cosgaya, J.M.; Aranda, A.; Cruces, J.; Martín-Blanco, E. Neuronal differentiation of PC12 cells induced by engrailed homeodomain is DNA-binding specific and independent of MAP kinases. J. Cell. Sci. 1998, 111, 2377–2384. [Google Scholar] [PubMed]
- Joliot, A.; Maizel, A.; Rosenberg, D.; Trembleau, A.; Dupas, S.; Volovitch, M.; Prochiantz, A. Identification of a signal sequence necessary for the unconventional secretion of Engrailed homeoprotein. Curr. Biol. 1998, 8, 856–863. [Google Scholar] [CrossRef]
- Morgan, R. Engrailed: Complexity and economy of a multi-functional transcription factor. FEBS Lett. 2006, 580, 2531–2533. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.; Bryan, R.T.; Javed, S.; Launchbury, F.; Zeegers, M.P.; Cheng, K.K.; James, N.D.; Wallace, D.M.; Hurst, C.D.; Ward, D.G.; et al. Expression of Engrailed-2 (EN2) protein in bladder cancer and its potential utility as a urinary diagnostic biomarker. Eur. J. Cancer 2013, 49, 2214–2222. [Google Scholar] [CrossRef]
- Martin, N.L.; Saba-El-Leil, M.K.; Sadekova, S.; Meloche, S.; Sauvageau, G. EN2 is a candidate oncogene in human breast cancer. Oncogene 2005, 24, 6890–6901. [Google Scholar] [CrossRef]
- Abollo-Jiménez, F.; Campos-Sánchez, E.; Toboso-Navasa, A.; Vicente-Dueñas, C.; González-Herrero, I.; Alonso-Escudero, E.; González, M.; Segura, V.; Blanco, O.; Martínez-Climent, J.A.; et al. Lineage-specific function of Engrailed-2 in the progression of chronic myelogenous leukemia to T-cell blast crisis. Cell Cycle 2014, 13, 1717–1726. [Google Scholar] [CrossRef]
- Bose, S.K.; Bullard, R.S.; Donald, C.D. Oncogenic role of engrailed-2 (en-2) in prostate cancer cell growth and survival. Transl. Oncogenomics 2008, 3, 37–43. [Google Scholar]
- Morgan, R.; Boxall, A.; Bhatt, A.; Bailey, M.; Hindley, R.; Langley, S.; Whitaker, H.C.; Neal, D.E.; Ismail, M.; Whitaker, H.; et al. Engrailed-2 (EN2): A tumor specific urinary biomarker for the early diagnosis of prostate cancer. Clin. Cancer Res. 2011, 17, 1090–1098. [Google Scholar] [CrossRef]
- Killick, E.; Morgan, R.; Launchbury, F.; Bancroft, E.; Page, E.; Castro, E.; Kote-Jarai, Z.; Aprikian, A.; Blanco, I.; Clowes, V.; et al. Role of Engrailed-2 (EN2) as a prostate cancer detection biomarker in genetically high risk men. Sci. Rep. 2013, 3, 1–5. [Google Scholar] [CrossRef]
- Pandha, H.; Sorensen, K.D.; Orntoft, T.F.; Langley, S.; Hoyer, S.; Borre, M.; Morgan, R. Urinary engrailed-2 (EN2) levels predict tumour volume in men undergoing radical prostatectomy for prostate cancer. BJU Int. 2012, 110, E287–E292. [Google Scholar] [CrossRef]
- Pandha, H.; HJaved, S.; Sooriakumaran, P.; Bott, S.; Montgomery, B.; Hutton, A.; Eden, C.; Langley, S.E.; Morgan, R. Correlation of Urinary Engrailed-2 Levels to Tumour Volume and Pathological Stage in Men Undergoing Radical Prostatectomy. J. Cancer Ther. 2013, 24, 726–733. [Google Scholar] [CrossRef][Green Version]
- Marszałł, M.P.; Sroka, W.; Adamowski, M.; Słupski, P.; Jarzemski, P.; Siódmiak, J.; Odrowąż-Sypniewska, G. Engrailed-2 protein as a potential urinary prostate cancer biomarker: A comparison study before and after digital rectal examination. Eur. J. Cancer Prev. 2015, 24, 51–56. [Google Scholar] [CrossRef]
- Egevad, L.; Srigley, J.R.; Delahunt, B. International Society of Urological Pathology (ISUP) consensus conference on handling and staging of radical prostatectomy specimens: Rationale and organization. Mod. Pathol. 2011, 24, 1–5. [Google Scholar] [CrossRef]
- Epstein, J.I.; Allsbrook, W.C.; Amin, M.B.; Egevad, L.L.; Committee, I.G. The 2005 International Society of Urological Pathology (ISUP) Consensus Conference on Gleason Grading of Prostatic Carcinoma. Am. J. Surg. Pathol. 2005, 29, 1228–1242. [Google Scholar] [CrossRef]
- Hormaechea-Agulla, D.; Gahete, M.D.; Jiménez-Vacas, J.M.; Gómez-Gómez, E.; Ibáñez-Costa, A.; L-López, F.; Rivero-Cortés, E.; Sarmento-Cabral, A.; Valero-Rosa, J.; Carrasco-Valiente, J.; et al. The oncogenic role of the In1-ghrelin splicing variant in prostate cancer aggressiveness. Mol. Cancer 2017, 16, 146. [Google Scholar] [CrossRef]
- Hormaechea-Agulla, D.; Jiménez-Vacas, J.M.; Gómez-Gómez, E.; L-López, F.; Carrasco-Valiente, J.; Valero-Rosa, J.; Moreno, M.M.; Sánchez-Sánchez, R.; Ortega-Salas, R.; Gracia-Navarro, F.; et al. The oncogenic role of the spliced somatostatin receptor sst5TMD4 variant in prostate cancer. FASEB J. 2017, 31, 4682–4696. [Google Scholar] [CrossRef]
- Hormaechea-Agulla, D.; Gómez-Gómez, E.; Ibáñez-Costa, A.; Carrasco-Valiente, J.; Rivero-Cortés, E.; L-López, F.; Pedraza-Arevalo, S.; Valero-Rosa, J.; Sánchez-Sánchez, R.; Ortega-Salas, R.; et al. Ghrelin O-acyltransferase (GOAT) enzyme is overexpressed in prostate cancer, and its levels are associated with patient’s metabolic status: Potential value as a non-invasive biomarker. Cancer Lett. 2016, 383, 125–134. [Google Scholar] [CrossRef]
- Sarmento-Cabral, A.; L-López, F.; Gahete, M.D.; Castaño, J.P.; Luque, R.M. Metformin Reduces Prostate Tumor Growth, in a Diet-Dependent Manner, by Modulating Multiple Signaling Pathways. Mol. Cancer Res. 2017, 15, 862–874. [Google Scholar] [CrossRef]
- L-López, F.; Sarmento-Cabral, A.; Herrero-Aguayo, V.; Gahete, M.D.; Castaño, J.P.; Luque, R.M. Obesity and metabolic dysfunction severely influence prostate cell function: Role of insulin and IGF1. J. Cell Mol. Med. 2017, 21, 1893–1904. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-Arévalo, S.; Hormaechea-Agulla, D.; Gómez-Gómez, E.; Requena, M.J.; Selth, L.A.; Gahete, M.D.; Castaño, J.P.; Luque, R.M. Somatostatin receptor subtype 1 as a potential diagnostic marker and therapeutic target in prostate cancer. Prostate 2017, 77, 1499–1511. [Google Scholar] [CrossRef] [PubMed]
- Schröder, F.H.; Hugosson, J.; Roobol, M.J.; Tammela, T.L.; Zappa, M.; Nelen, V.; Kwiatkowski, M.; Lujan, M.; Määttänen, L.; Lilja, H.; et al. Screening and prostate cancer mortality: Results of the European Randomised Study of Screening for Prostate Cancer (ERSPC) at 13 years of follow-up. Lancet 2014, 384, 2027–2035. [Google Scholar] [CrossRef]
- Gómez-Gómez, E.; Jiménez-Vacas, J.M.; Carrasco-Valiente, J.; Herrero-Aguayo, V.; Blanca-Pedregosa, A.M.; León-González, A.J.; Valero-Rosa, J.; Fernández-Rueda, J.L.; González-Serrano, T.; López-Miranda, J.; et al. Plasma ghrelin O-acyltransferase (GOAT) enzyme levels: A novel non-invasive diagnosis tool for patients with significant prostate cancer. J. Cell. Mol. Med. 2018. [Google Scholar] [CrossRef]
- McGrath, S.E.; Michael, A.; Morgan, R.; Pandha, H. EN2 in Prostate Cancer. Adv. Clin. Chem 2015, 71, 47–76. [Google Scholar] [PubMed]
- Morgan, R.; Boxall, A.; Harrington, K.J.; Simpson, G.R.; Michael, A.; Pandha, H.S. Targeting HOX transcription factors in prostate cancer. BMC Urol. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Cantile, M.; Franco, R.; Schiavo, G.; Procino, a.; Cindolo, L.; Botti, G.; Cillo, C. The HOX Genes Network in Uro-Genital Cancers: Mechanisms and Potential Therapeutic Implications. Curr. Med. Chem. 2011, 18, 4872–4884. [Google Scholar] [CrossRef]
- Cunningham, D.; You, Z. In vitro and in vivo model systems used in prostate cancer research. J. Biol. Methods 2015, 2, 17. [Google Scholar] [CrossRef]
- Courtney, K.D.; Corcoran, R.B.; Engelman, J.A. The PI3K pathway as drug target in human cancer. J. Clin. Oncol. 2010, 28, 1075–1083. [Google Scholar] [CrossRef]
- Network, C.G.A.R. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar]
- Xin, L.; Teitell, M.A.; Lawson, D.A.; Kwon, A.; Mellinghoff, I.K.; Witte, O.N. Progression of prostate cancer by synergy of AKT with genotropic and nongenotropic actions of the androgen receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 7789–7794. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, I.; Day, T.K.; Tilley, W.D.; Selth, L.A. Androgen receptor signaling in castration-resistant prostate cancer: A lesson in persistence. Endocr. Relat. Cancer 2016, 23, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Baron, V.T.; Pio, R.; Jia, Z.; Mercola, D. Early Growth Response 3 regulates genes of inflammation and directly activates IL6 and IL8 expression in prostate cancer. Br. J. Cancer 2015, 112, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Wang, H.; Peskar, B.M.; Levin, E.; Itani, R.M.; Sarfeh, I.J.; Tarnawski, A.S. Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs: Insight into mechanisms and implications for cancer growth and ulcer healing. Nat. Med. 1999, 5, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Pio, R.; Jia, Z.; Baron, V.T.; Mercola, D. Early growth response 3 (Egr3) is highly over-expressed in non-relapsing prostate cancer but not in relapsing prostate cancer. PLoS ONE 2013, 8, e54096. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, R.J.; Dijkstra, S.; Smit, F.P.; Vandersmissen, J.; Van de Voorde, H.; Mulders, P.F.A.; van Oort, I.M.; Van Criekinge, W.; Schalken, J.A. Epigenetic markers in circulating cell-free DNA as prognostic markers for survival of castration-resistant prostate cancer patients. Prostate 2018, 78, 336–342. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Overall |
|---|---|
| Number of patients | 33 |
| Age at diagnosis | |
| Median (IQR) | 62 (58–66) |
| BMI | |
| Median (IQR) | 27.7 (25.8–31.3) |
| PSA level, ng/mL | |
| Median (IQR) | 6 (4.4–9.5) |
| Gleason score in prostatectomy specimen (%) | |
| 6 | 9 (27.3) |
| 7 | 23 (69.7) |
| 8 | 1 (3) |
| EE, n° (%) | 21 (63.6) |
| PI, n° (%) | 28 (84.8) |
| VI, n° (%) | 8 (24.2) |
| Relative EN2 mRNA expression in FFPE piece a | |
| Tumor tissue | |
| Median (IQR) | 0.173 (0.002–1.473) |
| Non-tumor adjacent tissue | |
| Median (IQR) | 0.008 (0.000–0.477) |
| Ratio tumor/non-tumor tissue | |
| Median (IQR) | 3.451 (1.260–12.212) |
| Relative EN1 mRNA expression in FFPE piece * | |
| Tumor tissue | |
| Median (IQR) | 0.723 (0.209–2.828) |
| Non-tumor adjacent tissue | |
| Median (IQR) | 0.421 (0.149–1.145) |
| Ratio tumor/non-tumor tissue | |
| Median (IQR) | 1.197 (0.325–3.249) |
| Variable | Overall | Control | PCa |
|---|---|---|---|
| Patients | 30 | 7 | 23 |
| Age at diagnosis | |||
| Median (IQR) | 73 (64–79) | 67 (59–79) | 76 (67.0–80.0) |
| PSA level, ng/mL | |||
| Median (IQR) | - | - | 40 (22–70) |
| Dyslipidemia (%) | 7 (23.3) | 2 (28.6) | 5 (21.7) |
| Diabetes (%) | 8 (26.7) | 2 (28.6) | 6 (26.1) |
| * BMI | |||
| Median (IQR) | 27.19 (25.2–29.83) | 25.87 (24.50–34.24) | 27.44 (25.46–29.65) |
| Gleason score | |||
| =7 | - | - | 8 (34.8) |
| >7 | - | - | 15 (65.2) |
| EE (%) | - | - | 6 (26.1) |
| PI (%) | - | - | 14 (60.9) |
| # Metastasis (%) | - | - | 13 (56.5) |
| N° samples (%) in whichEN2 was detected | 25 (83.3) | 3 (42.9) | 22 (95.7) |
| ª Median(IQR) EN2 mRNAexpression | 445 (8–2265) | 0 (0–9) | 874 (173–2650) |
| Variable | Control (n = 10) | Negative Biopsy (n = 10) | PCa (n = 24) | p-Value |
|---|---|---|---|---|
| Age, Years | ||||
| Median (IQR) | 56 (52–61) | 56 (53–59) | 68 (59–71) | <0.01 |
| Waist circumference, cm | ||||
| Median (IQR) | 107 (99.5–111.8) | 103 (98.3–109) | 104.5 (100–111.5) | 0.88 |
| BMI | ||||
| Median (IQR) | 32.4 (27.5–33.1) | 30.1 (27.33–31.97) | 29 (26.67–30.48) | 0.15 |
| PSA, ng/mL | ||||
| Median (IQR) | 0.87 (0.6–1.6) | 3.6 (3.0–4.0) | 5.7 (4.6–9.7) | <0.01 |
| DRE, abnormal (%) | 0 (0) | 0 (0) | 9 (37.5) | <0.01 |
| Prostate Vol, cc | ||||
| Median (IQR) | - | 37 (23.1–45.8) | 37 (25.59–52.5) | 0.55 |
| 1° Biopsy (%) | - | 9 (90) | 17 (70.8) | |
| Gleason grade (%) | ||||
| =6 | 6 (25) | |||
| ≥7 | 18 (75) | |||
| Nº Pathologic cores | ||||
| Median (IQR) | 2 (1–4) | |||
| EN2 Urine | ||||
| Median (IQR) | 0 (0–0.21) | 0.02 (0.00–0.30) | 0.19 (0.01–0.43) | 0.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Gómez, E.; Jiménez-Vacas, J.M.; Pedraza-Arévalo, S.; López-López, F.; Herrero-Aguayo, V.; Hormaechea-Agulla, D.; Valero-Rosa, J.; Ibáñez-Costa, A.; León-González, A.J.; Sánchez-Sánchez, R.; et al. Oncogenic Role of Secreted Engrailed Homeobox 2 (EN2) in Prostate Cancer. J. Clin. Med. 2019, 8, 1400. https://doi.org/10.3390/jcm8091400
Gómez-Gómez E, Jiménez-Vacas JM, Pedraza-Arévalo S, López-López F, Herrero-Aguayo V, Hormaechea-Agulla D, Valero-Rosa J, Ibáñez-Costa A, León-González AJ, Sánchez-Sánchez R, et al. Oncogenic Role of Secreted Engrailed Homeobox 2 (EN2) in Prostate Cancer. Journal of Clinical Medicine. 2019; 8(9):1400. https://doi.org/10.3390/jcm8091400
Chicago/Turabian StyleGómez-Gómez, Enrique, Juan M. Jiménez-Vacas, Sergio Pedraza-Arévalo, Fernando López-López, Vicente Herrero-Aguayo, Daniel Hormaechea-Agulla, José Valero-Rosa, Alejandro Ibáñez-Costa, Antonio J. León-González, Rafael Sánchez-Sánchez, and et al. 2019. "Oncogenic Role of Secreted Engrailed Homeobox 2 (EN2) in Prostate Cancer" Journal of Clinical Medicine 8, no. 9: 1400. https://doi.org/10.3390/jcm8091400
APA StyleGómez-Gómez, E., Jiménez-Vacas, J. M., Pedraza-Arévalo, S., López-López, F., Herrero-Aguayo, V., Hormaechea-Agulla, D., Valero-Rosa, J., Ibáñez-Costa, A., León-González, A. J., Sánchez-Sánchez, R., González-Serrano, T., Requena-Tapia, M. J., Castaño, J. P., Carrasco-Valiente, J., Gahete, M. D., & Luque, R. M. (2019). Oncogenic Role of Secreted Engrailed Homeobox 2 (EN2) in Prostate Cancer. Journal of Clinical Medicine, 8(9), 1400. https://doi.org/10.3390/jcm8091400