Moderate Cardiovascular Exercise Speeds Up Neural Markers of Stimulus Evaluation During Attentional Control Processes


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
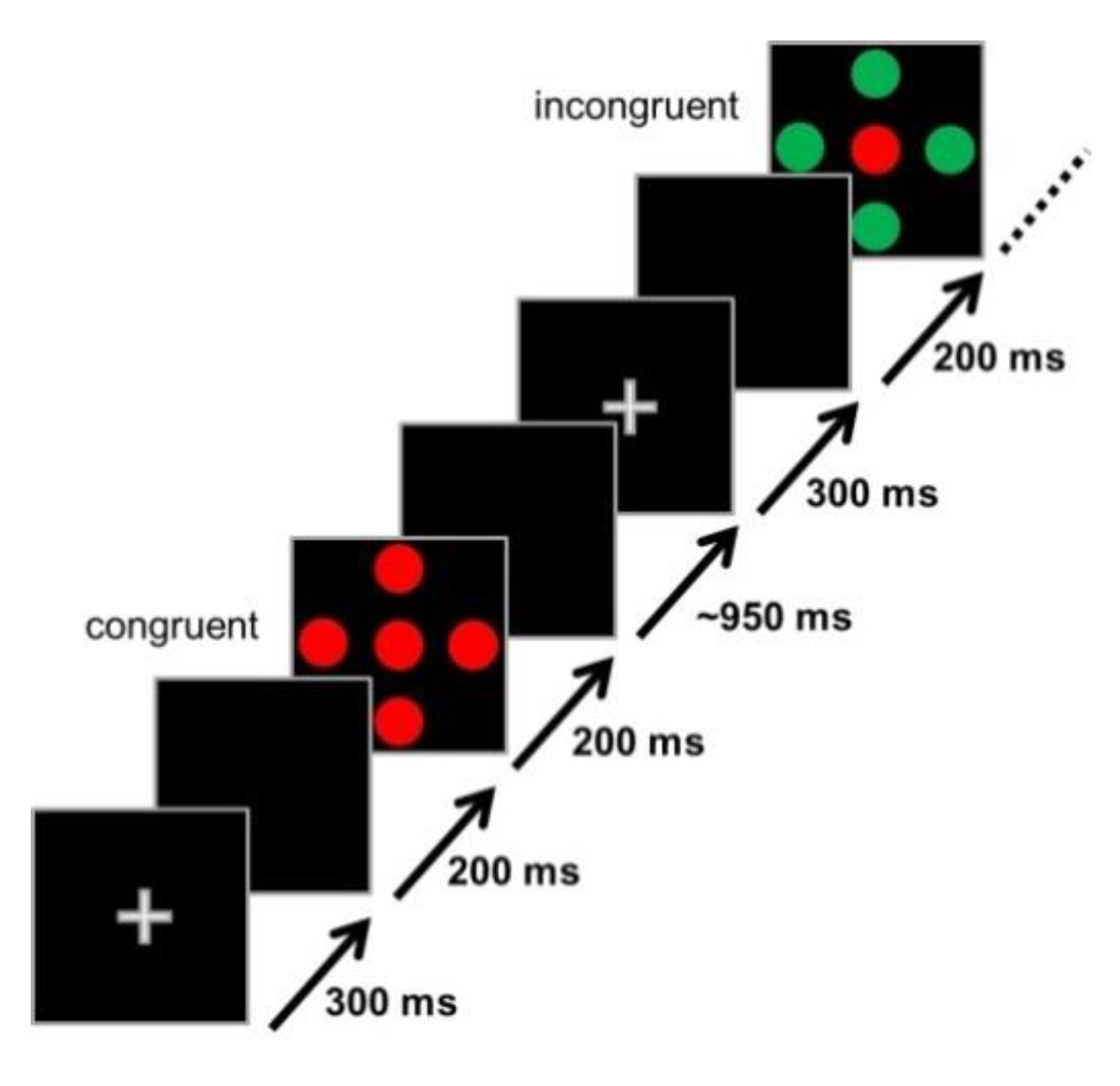
2.2. Stimuli and Experimental Design (Flanker Task)
2.3. Cardiorespiratory Fitness Assessment
2.4. Exercise Intervention
2.5. EEG Data Acquisition and Analyses
2.6. Testing Procedure
2.7. Statistics
3. Results
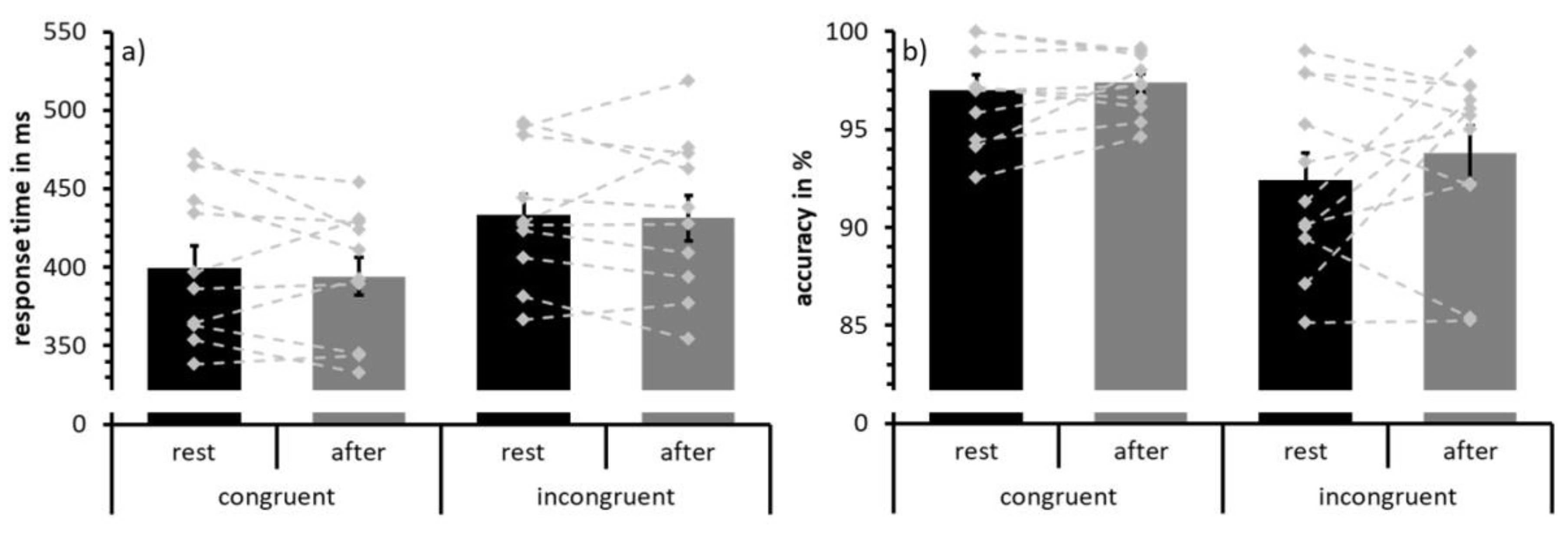
3.1. Behavioral Data
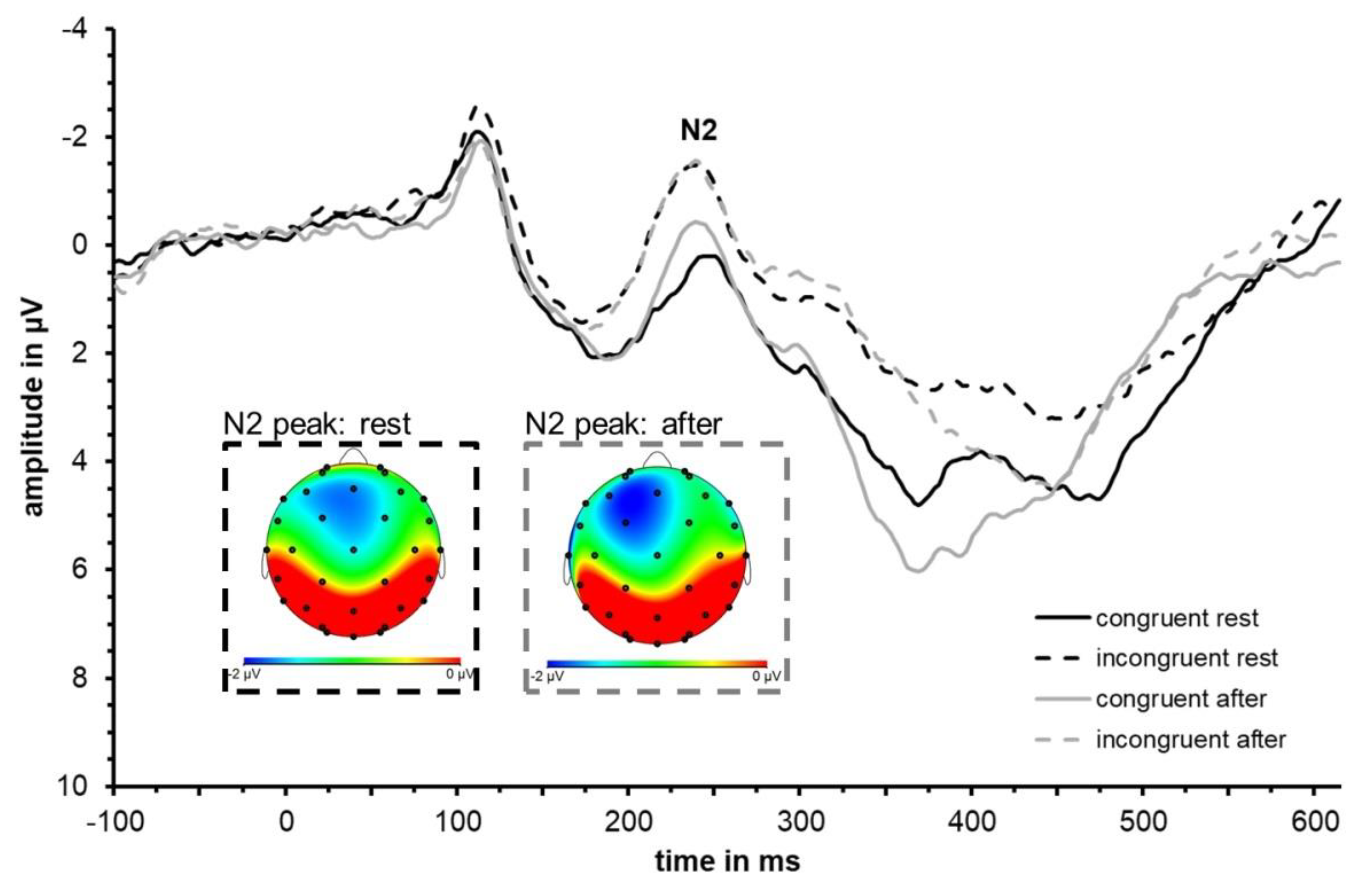
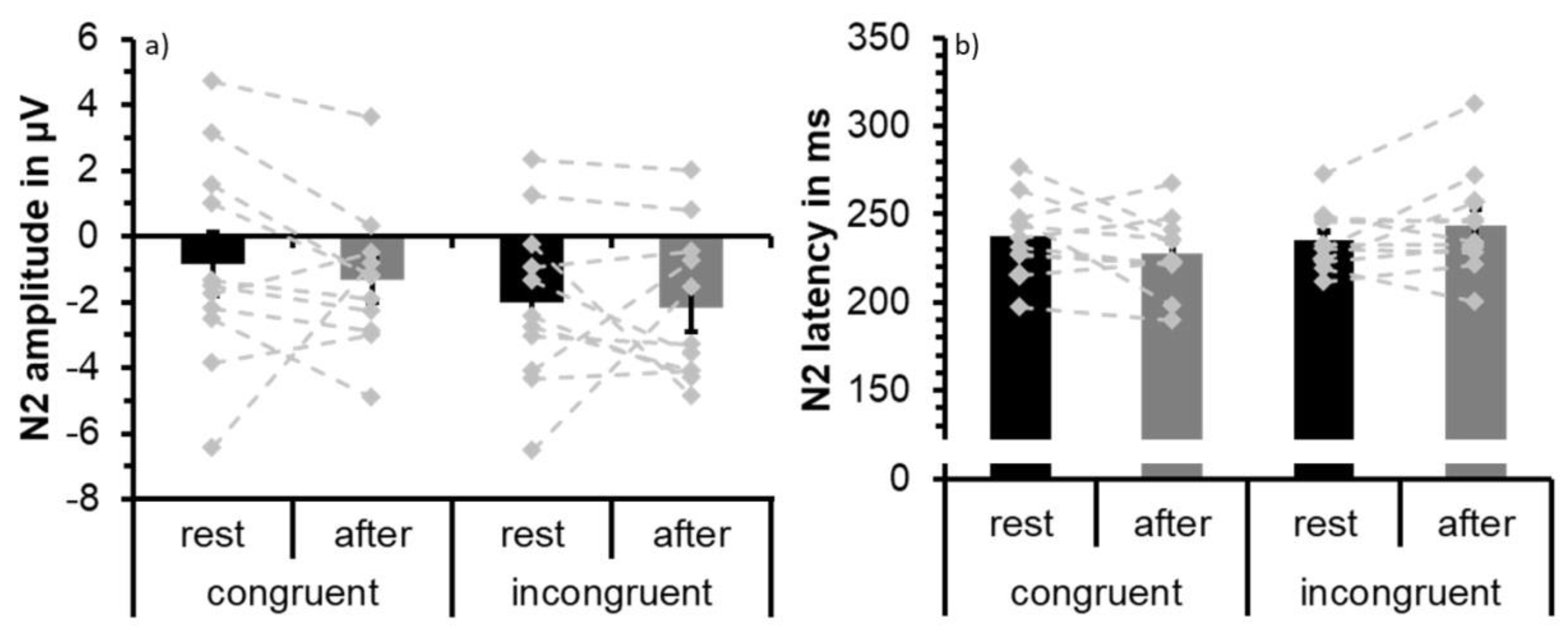
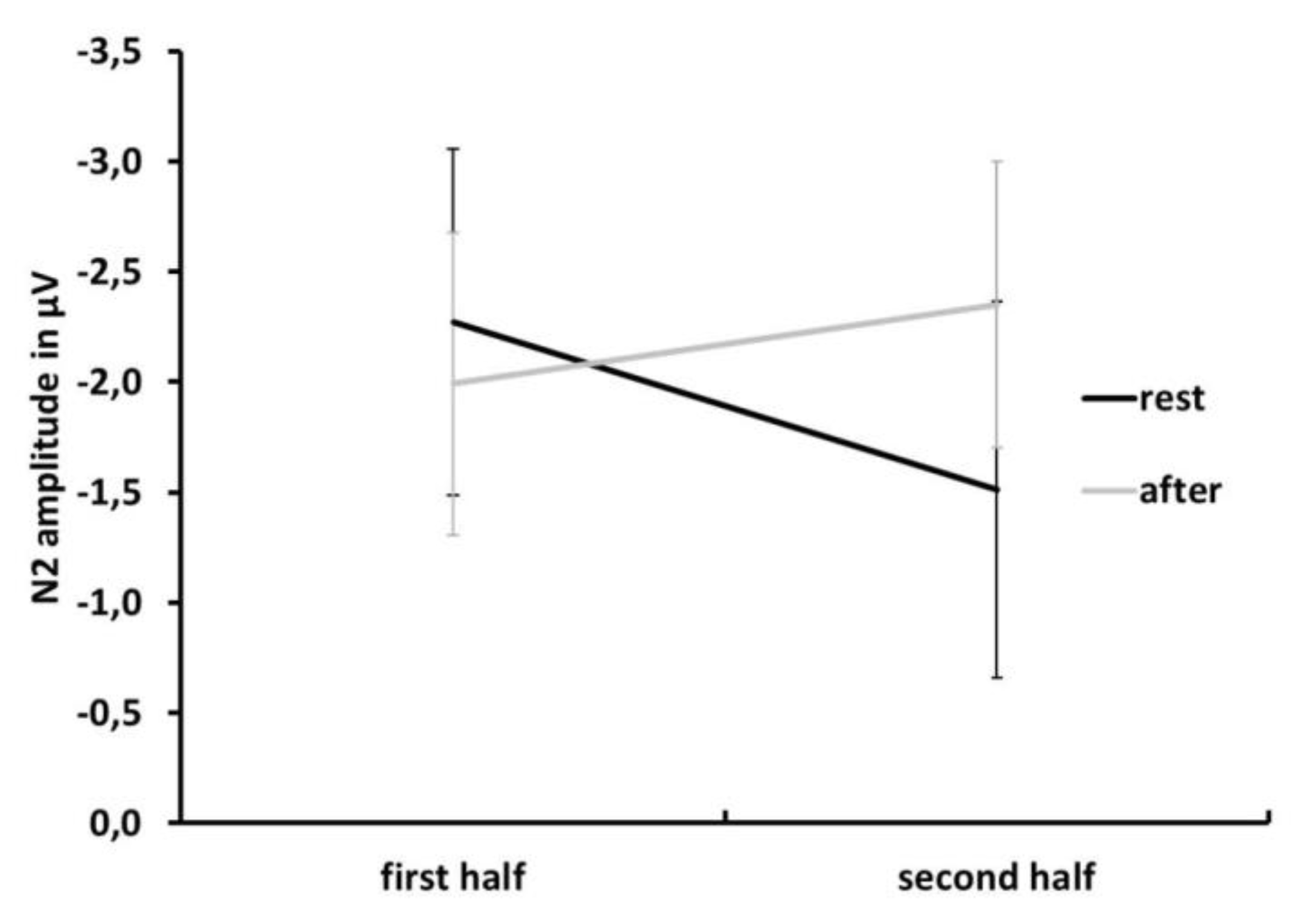
3.2. ERP Data: N2
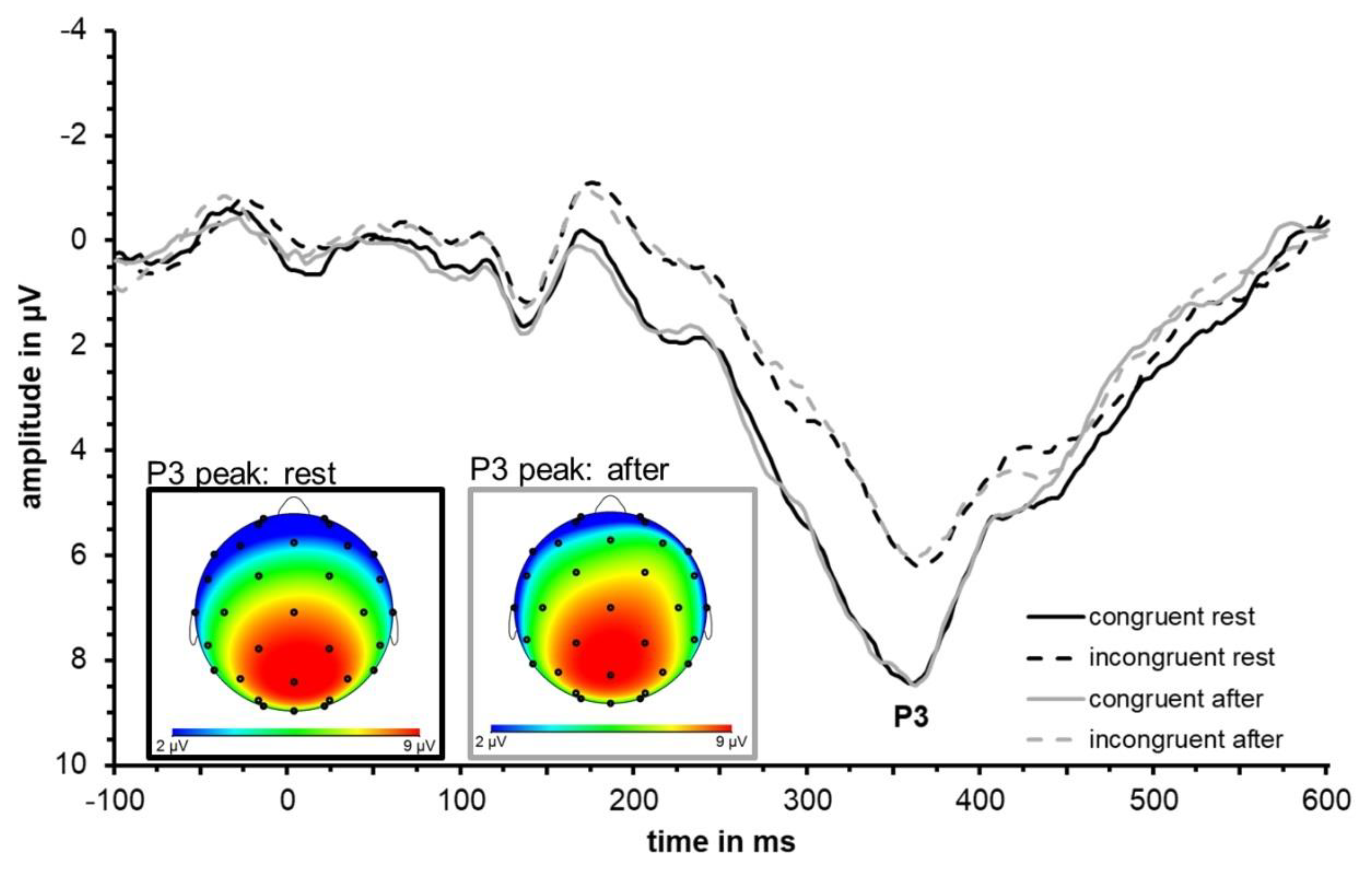
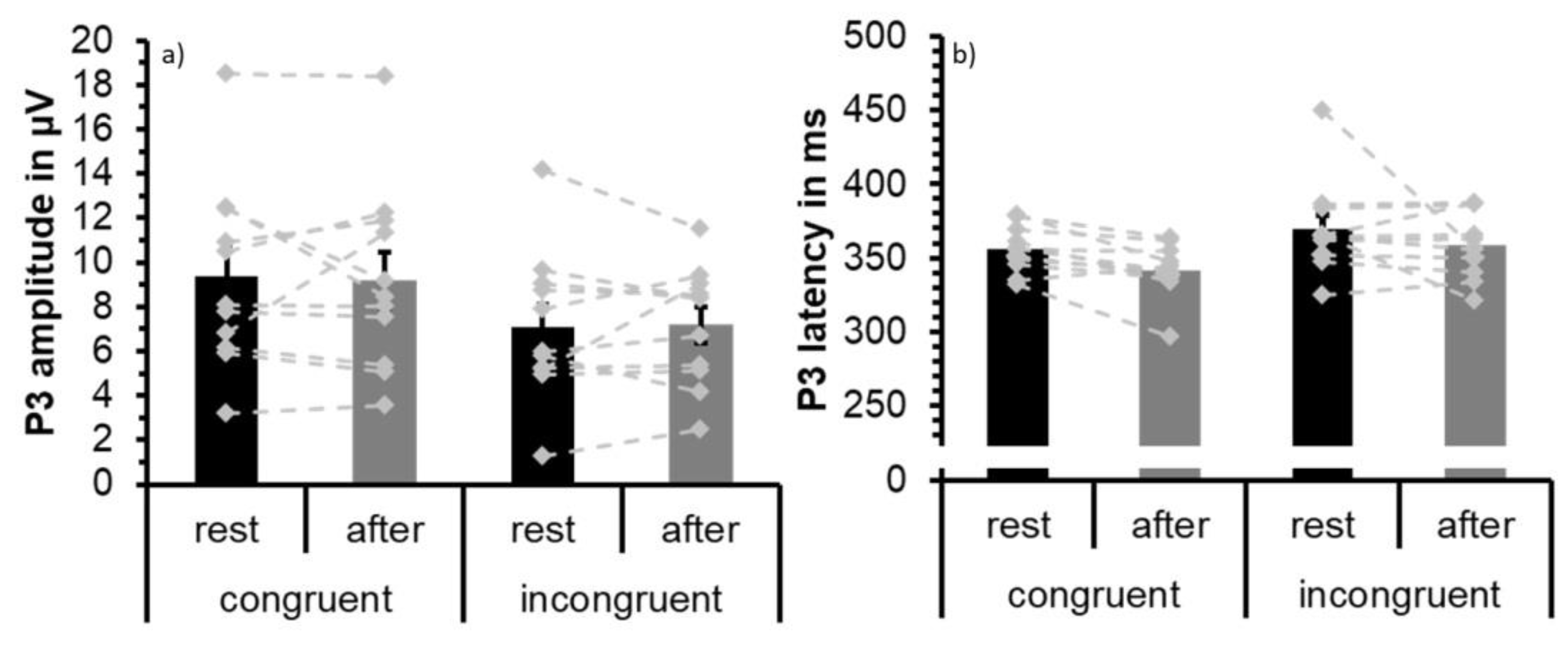
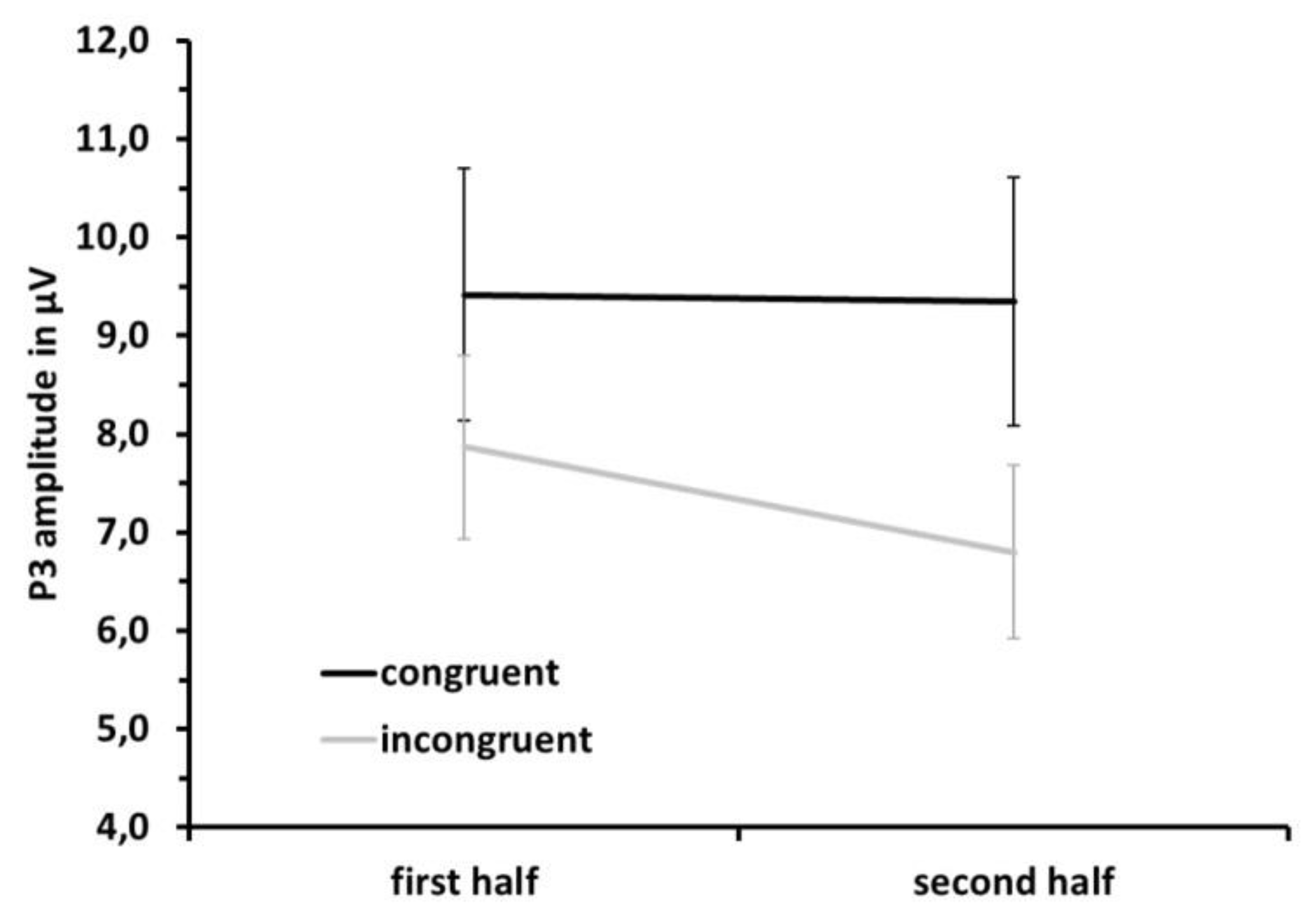
3.3. ERP Data: P3
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Audiffren, M. Exercise and cognitive function. In Acute Exercise and Psychological Functions: A Cognitive-Energetic Approach; McMorris, T., Tomporowski, P., Audiffren, M., McMorris, T., Tomporowski, P., Audiffren, M., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 3–39. [Google Scholar]
- Brisswalter, J.; Collardeau, M.; Rene, A. Effects of acute physical exercise characteristics on cognitive performance. Sports Med. 2002, 32, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Labban, J.D.; Gapin, J.I.; Etnier, J.L. The effects of acute exercise on cognitive performance: A meta-analysis. Brain Res. 2012, 1453, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Roig, M.; Nordbrandt, S.; Geertsen, S.S.; Nielsen, J.B. The effects of cardiovascular exercise on human memory: A review with meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 1645–1666. [Google Scholar] [CrossRef] [PubMed]
- Tomporowski, P.D. Effects of acute bouts of exercise on cognition. Acta Psychol. 2003, 112, 297–324. [Google Scholar] [CrossRef]
- Voelcker-Rehage, C.; Niemann, C.; Hübner, L.; Godde, B.; Winneke, A.H. Benefits of Physical Activity and Fitness for Lifelong Cognitive and Motor Development—Brain and Behavior, Sport and Exercise Psychology Research; Elsevier: Amsterdam, The Netherlands, 2016; pp. 43–73. [Google Scholar]
- Lambourne, K.; Tomporowski, P. The effect of exercise-induced arousal on cognitive task performance: A meta-regression analysis. Brain Res. 2010, 1341, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, K.; Nishihira, Y.; Higashiura, T.; Kuroiwa, K. The interactive effect of exercise intensity and task difficulty on human cognitive processing. Int. J. Psychophysiol. 2007, 65, 114–121. [Google Scholar] [CrossRef]
- Kamijo, K. Enhancing cognitive functioning and brain plasticity. In Effects of Acute Exercise on Event-Related Brain Potentials; Chodzko-Zajko, W., Kramer, A.F., Poon, L.W., Chodzko-Zajko, W., Kramer, A.F., Poon, L.W., Eds.; Human Kinetics: Champaign, IL, USA, 2009; pp. 111–132. [Google Scholar]
- McMorris, T.; Davranche, K.; Jones, G.; Hall, B.; Corbett, J.; Minter, C. Acute incremental exercise, performance of a central executive task, and sympathoadrenal system and hypothalamic-pituitary-adrenal axis activity. Int. J. Psychophysiol. 2009, 73, 334–340. [Google Scholar] [CrossRef]
- Themanson, J.R.; Hillman, C.H. Cardiorespiratory fitness and acute aerobic exercise effects on neuroelectric and behavioral measures of action monitoring. Neuroscience 2006, 141, 757–767. [Google Scholar] [CrossRef]
- Budde, H.; Voelcker-Rehage, C.; Pietrabyk-Kendziorra, S.; Ribeiro, P.; Tidow, G. Acute coordinative exercise improves attentional performance in adolescents. Neurosci. Lett. 2008, 441, 219–223. [Google Scholar] [CrossRef]
- Budde, H.; Pietrassyk-Kendziorra, S.; Bohm, S.; Voelcker-Rehage, C. Hormonal responses to physical and cognitive stress in a school setting. Neurosci. Lett. 2010, 474, 131–134. [Google Scholar] [CrossRef]
- Pontifex, M.B.; Saliba, B.J.; Raine, L.B.; Picchietti, D.L.; Hillman, C.H. Exercise improves behavioral, neurocognitive, and scholastic performance in children with attention-deficit/hyperactivity disorder. J. Pediatr. 2013, 162, 543–551. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.; Sproule, J.; Turner, A.; Hale, B.J. Acute, intermediate intensity exercise, and speed and accuracy in working memory tasks: A meta-analytical comparison of effects. Physiol. Behav. 2011, 102, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Hillman, C.H.; Snook, E.M.; Jerome, G.J. Acute cardiovascular exercise and executive control function. Int. J. Psychophysiol. 2003, 48, 307–314. [Google Scholar] [CrossRef]
- Scudder, M.R.; Drollette, E.S.; Pontifex, M.B.; Hillman, C.H. Neuroelectric indices of goal maintenance following a single bout of physical activity. Biol. Psychol. 2012, 89, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.C.; Westfall, D.R.; Soneson, J.; Gurd, B.; Hillman, C.H. Comparison of the acute effects of high-intensity interval training and continuous aerobic walking on inhibitory control. Psychophysiology 2017, 54, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Alderman, B.L.; Chu, C.H.; Wang, C.C.; Song, T.F.; Chen, F.T. Acute exercise has a general facilitative effect on cognitive function: A combined ERP temporal dynamics and BDNF study. Psychophysiology 2017, 54, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Pontifex, M.B.; Parks, A.C.; Henning, D.A.; Kamijo, K. Single bouts of exercise selectively sustain attentional processes. Psychophysiology 2015, 52, 618–625. [Google Scholar] [CrossRef]
- Kamijo, K.; Hayashi, Y.; Sakai, T.; Yahiro, T.; Tanaka, K.; Nishihira, Y. Acute effects of aerobic exercise on cognitive function in older adults. J. Gerontol. B Psychol. Sci. Soc. Sci. 2009, 64, 356–363. [Google Scholar] [CrossRef]
- Akatsuka, K.; Yamashiro, K.; Nakazawa, S.; Mitsuzono, R.; Maruyama, A. Acute aerobic exercise influences the inhibitory process in the go/no-go task in humans. Neurosci. Lett. 2015, 600, 80–84. [Google Scholar] [CrossRef]
- Pontifex, M.B.; McGowan, A.L.; Chandler, M.C.; Gwizdala, K.L.; Parks, A.C.; Fenn, K.; Kamijo, K. A primer on investigating the after effects of acute bouts of physical activity on cognition. Psychol. Sport Exerc. 2019, 40, 1–22. [Google Scholar] [CrossRef]
- Eriksen, B.A.; Eriksen, C.W. Effects of noise letters upon the identification of a target letter in a nonsearch task. Percept. Psychophys. 1974, 16, 143–149. [Google Scholar] [CrossRef]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A. The unity and diversity of executive functions and their contributions to complex ‘frontal lobe’ tasks: A latent variable analysis. Cognit. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Unsworth, N.; Spillers, G.J. Working memory capacity: Attention control, secondary memory, or both? A direct test of the dual-component model. J. Mem. Lang. 2010, 62, 392–406. [Google Scholar] [CrossRef]
- Yeung, N.; Botvinick, M.M.; Cohen, J.D. The Neural Basis of Error Detection: Conflict Monitoring and the Error-Related Negativity. Psychol. Rev. 2004, 111, 931–959. [Google Scholar] [CrossRef] [PubMed]
- Polich, J.; Lardon, M.T. P300 and long-term physical exercise. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 493–498. [Google Scholar] [CrossRef]
- Kok, A. On the utility of P3 amplitude as a measure of processing capacity. Psychophysiology 2001, 38, 557–577. [Google Scholar] [CrossRef] [PubMed]
- Frtusova, J.B.; Winneke, A.H.; Phillips, N.A. ERP evidence that auditory-visual speech facilitates working memory in younger and older adults. Psychol. Aging 2013, 28, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Segalowitz, S.J.; Wintink, A.J.; Cudmore, L.J. P3 topographical change with task familiarization and task complexity. Cogn. Brain Res. 2001, 12, 451–457. [Google Scholar] [CrossRef]
- Wild-Wall, N.; Falkenstein, M.; Gajewski, P.D. Age-related differences in working memory performance in a 2-back task. Front. Psychol. 2011, 2, 186. [Google Scholar] [CrossRef]
- Daffner, K.R.; Chong, H.; Sun, X.; Tarbi, E.C.; Riis, J.L.; McGinnis, S.M.; Holcomb, P.J. Mechanisms underlying age- and performance-related differences in working memory. J. Cogn. Neurosci. 2011, 23, 1298–1314. [Google Scholar] [CrossRef]
- Coles, M.G.; Gratton, G.; Bashore, T.R.; Eriksen, C.W.; Donchin, E. A psychophysiological investigation of the continuous flow model of human information processing. J. Exp. Psychol. Hum. Percept. Perform. 1985, 11, 529–553. [Google Scholar] [CrossRef] [PubMed]
- Donchin, E.; Coles, M.G. Is the P300 component a manifestation of context updating? Behav. Brain Sci. 1988, 11, 357–374. [Google Scholar] [CrossRef]
- Ridderinkhof, K.R.; van der Molen, M.W. A psychophysiological analysis of developmental differences in the ability to resist interference. Child Dev. 1995, 66, 1040–1056. [Google Scholar] [CrossRef]
- Pontifex, M.B.; Hillman, C.H. Neuroelectric and behavioral indices of interference control during acute cycling. Clin. Neurophysiol. 2007, 118, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Drollette, E.S.; Scudder, M.R.; Raine, L.B.; Moore, R.D.; Saliba, B.J.; Pontifex, M.B.; Hillman, C.H. Acute exercise facilitates brain function and cognition in children who need it most: An ERP study of individual differences in inhibitory control capacity. Dev. Cogn. Neurosci. 2014, 7, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Magnie, M.N.; Bermon, S.; Martin, F.; Madany-Lounis, M.; Suisse, G.; Muhammad, W.; Dolisi, C. P300, N400, aerobic fitness, and maximal aerobic exercise. Psychophysiology 2000, 37, 369–377. [Google Scholar] [CrossRef]
- Nakamura, Y.; Nishimoto, K.; Akamatu, M.; Takahashi, M.; Maruyama, A. The effect of jogging on P300 event related potentials. Electromyogr. Clin. Neurophysiol. 1999, 39, 71–74. [Google Scholar]
- Chang, Y.K.; Pesce, C.; Chiang, Y.T.; Kuo, C.Y.; Fong, D.Y. Antecedent acute cycling exercise affects attention control: An ERP study using attention network test. Front. Hum. Neurosci. 2015, 9, 156. [Google Scholar] [CrossRef]
- Chu, C.H.; Kramer, A.F.; Song, T.F.; Wu, C.H.; Hung, T.M.; Chang, Y.K. Acute Exercise and Neurocognitive Development in Preadolescents and Young Adults: An ERP Study. Neural Plast. 2017, 2017, 2631909. [Google Scholar] [CrossRef]
- Kamijo, K.; Nishihira, Y.; Hatta, A.; Kaneda, T.; Wasaka, T.; Kida, T.; Kuroiwa, K. Differential influences of exercise intensity on information processing in the central nervous system. Eur. J. Appl. Physiol. 2004, 92, 305–311. [Google Scholar] [CrossRef]
- Chang, Y.K.; Chu, C.H.; Wang, C.C.; Song, T.F.; Wei, G.X. Effect of acute exercise and cardiovascular fitness on cognitive function: An event-related cortical desynchronization study. Psychophysiology 2015, 52, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol in psychoneuroendocrine research: Recent developments and applications. Psychoneuroendocrinology 1994, 19, 313–333. [Google Scholar] [CrossRef]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Winneke, A.H.; Godde, B.; Reuter, E.; Vieluf, S.; Voelcker-Rehage, C. The association between physical activity and attentional control in younger and older middle-aged adults: An ERP study. GeroPsych J. Gerontopsychol. Geriatr. Psychiatry 2012, 25, 207–221. [Google Scholar] [CrossRef]
- Voelcker-Rehage, C.; Godde, B.; Staudinger, U.M. Physical and motor fitness are both related to cognition in old age. Eur. J. Neurosci. 2010, 31, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Mezzani, A.; Agostoni, P.; Cohen-Solal, A.; Corra, U.; Jegier, A.; Kouidi, E.; Mazic, S.; Meurin, P.; Piepoli, M.; Simon, A. Standards for the use of cardiopulmonary exercise testing for the functional evaluation of cardiac patients: A report from the Exercise Physiology Section of the European Association for Cardiovascular Prevention and Rehabilitation. Eur. J. Cardiovasc. Prev. Rehabil. 2009, 16, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Beaver, W.L.; Wasserman, K.; Whipp, B.J. A new method for detecting anaerobic threshold by gas exchange. J. Appl. Physiol. 1986, 60, 2020–2027. [Google Scholar] [CrossRef]
- Voelcker-Rehage, C.; Godde, B.; Staudinger, U.M. Cardiovascular and coordination training differentially improve cognitive performance and neural processing in older adults. Front. Hum. Neurosci. 2011, 5, 26. [Google Scholar] [CrossRef]
- Meyer, T.; Lucia, A.; Earnest, C.P.; Kindermann, W. A conceptual framework for performance diagnosis and training prescription from submaximal gas exchange parameters—Theory and application. Int. J. Sports Med. 2005, 26, S38–S48. [Google Scholar] [CrossRef]
- Prioux, J.; Ramonatxo, M.; Hayot, M.; Mucci, P.; Prefaut, C. Effect of ageing on the ventilatory response and lactate kinetics during incremental exercise in man. Eur. J. Appl. Physiol. 2000, 81, 100–107. [Google Scholar] [CrossRef]
- Jasper, H.H. The ten-twenty electrode system of the international federation of electroencephalography and clinical neurophysiology. Electroencephalogr. Clin. Neurophysiol. 1958, 183, 321–375. [Google Scholar]
- Greenhouse, S.W.; Geisser, S. On methods in the analysis of profile data. Psychometrika 1959, 24, 95–112. [Google Scholar] [CrossRef]
- Luck, S.J. An Introduction to the Event-Related Potential Technique (Cognitive Neuroscience); MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Kutas, M.; McCarthy, G.; Donchin, E. Augmenting mental chronometry: The P300 as a measure of stimulus evaluation time. Science 1977, 197, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Grego, F.; Vallier, J.M.; Collardeau, M.; Bermon, S.; Ferrari, P.; Candito, M.; Bayer, P.; Magnie, M.N.; Brisswalter, J. Effects of long duration exercise on cognitive function, blood glucose, and counterregulatory hormones in male cyclists. Neurosci. Lett. 2004, 364, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Yagi, Y.; Coburn, K.L.; Estes, K.M.; Arruda, J.E. Effects of aerobic erxercise and gender on visual and auditory P300, reaction time, and accuracy. Eur. J. Appl. Physiol. 1999, 80, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Audiffren, M. The reticular-activating hypofrontality (RAH) model of acute exercise. Neurosci. Biobehav. Rev. 2011, 35, 1305–1325. [Google Scholar] [CrossRef] [PubMed]
- Pagliari, R.; Peyrin, L. Norepinephrine release in the rat frontal cortex under treadmill exercise: A study with microdialysis. J. Appl. Physiol. 1995, 78, 2121–2130. [Google Scholar] [CrossRef]
- Meeusen, R.; Smolders, I.; Sarre, S.; de Meirleir, K.; Keizer, H.; Serneels, M.; Ebinger, G.; Michotte, Y. Endurance training effects on neurotransmitter release in rat striatum: An in vivo microdialysis study. Acta Physiol. Scand. 1997, 159, 335–341. [Google Scholar] [CrossRef]
- McMorris, T.; Collard, K.; Corbett, J.; Dicks, M.; Swain, J.P. A test of the catecholamines hypothesis for an acute exercise cognition interaction. Pharmacol. Biochem. Behav. 2008, 89, 106–115. [Google Scholar] [CrossRef]
- Winter, B.; Breitenstein, C.; Mooren, F.C.; Voelker, K.; Fobker, M.; Lechtermann, A.; Krueger, K.; Fromme, A.; Korsukewitz, C.; Floel, A.; et al. High impact running improves learning. Neurobiol. Learn. Mem. 2007, 87, 597–609. [Google Scholar] [CrossRef]
- Bäckman, L.; Ginovart, N.; Dixon, R.A.; Wahlin, T.B.R.; Wahlin, A.; Halldin, C.; Farde, L. Age-related cognitive deficits mediated by changes in the striatal dopamine system. Am. J. Psychiatry 2000, 157, 635–637. [Google Scholar] [CrossRef]
- Rieckmann, A.; Karlsson, S.; Karlsson, P.; Brehmer, Y.; Fischer, H.; Farde, L.; Nyberg, L.; Bäckman, L. Dopamine D1 receptor associations within and between Dopaminergic pathways in younger and elderly adults: Links to cognitive performance. Cereb. Cortex 2011, 21, 2023–2032. [Google Scholar] [CrossRef]
- Basso, J.C.; Suzuki, W.A. The Effects of Acute Exercise on Mood, Cognition, Neurophysiology, and Neurochemical Pathways: A Review. Brain Plast. 2017, 2, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Pogarell, O.; Padberg, F.; Karch, S.; Segmiller, F.; Juckel, G.; Mulert, C.; Hegerl, U.; Tatsch, K.; Koch, W. Dopaminergic mechanisms of target detection—P300 event related potential and striatal dopamine. Psychiatry Res. Neuroimaging 2011, 194, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Finkenzeller, T.; Doppelmayr, M.; Würth, S.; Amesberger, G. Impact of maximal physical exertion on interference control and electrocortical activity in well-trained persons. Eur. J. Appl. Physiol. 2018, 118, 2509–2521. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPM | Target | At Rest | Avg. during Exercise | After 20 min. Exercise | Flanker Start | Flanker End |
|---|---|---|---|---|---|---|
| Avg. | 113.00 | 74.73 | 114.65 | 119.55 | 82.36 | 78.43 |
| SD | 13.21 | 12.92 | 12.98 | 14.25 | 11.50 | 9.13 |
| Min | 96.00 | 60.00 | 92.40 | 95.00 | 68.00 | 68.00 |
| Max | 140.00 | 100.00 | 139.20 | 149.00 | 100.00 | 97.00 |
| Response time | F(df) | MSE | partial η2 | p |
| Condition | F(1,10) = 52.42 | 274.84 | 0.840 | <0.001 * |
| Time | F(1,10) = 0.03 | 520.72 | 0.003 | 0.862 |
| Condition × Time | F(1,10) = 0.003 | 217.06 | 0.000 | 0.960 |
| Response accuracy | F(df) | MSE | partial η2 | p |
| Condition | F(1,10) = 16.76 | 11.09 | 0.626 | 0.002 * |
| Time | F(1,10) = 1.72 | 4.81 | 0.147 | 0.219 |
| Condition × Time | F(1,10) = 0.42 | 6.49 | 0.041 | 0.530 |
| N2 | Rest | After Exercise | ||
|---|---|---|---|---|
| Latency in ms | Amplitude in µV | Latency in ms | Amplitude in µV | |
| Congruent | 238.81 (28.56) | −1.07 (3.75) | 231.00 (27.57) | −1.40 (2.58) |
| Incongruent | 243.79 (30.76) | −2.36 (3.03) | 245.03 (30.39) | −2.55 (2.66) |
| N2 Amplitude | F(DF) | MSE | Partial H2 | P |
|---|---|---|---|---|
| Condition | F(1,10) = 7.25 | 9.41 | 0.420 | 0.023 * |
| Time | F(1,10) = 0.25 | 30.71 | 0.024 | 0.628 |
| Electrode | F(5,50) = 1.14 | 21.75 | 0.102 | 0.328 |
| Cond × Time | F(1,10) = 0.24 | 7.46 | 0.024 | 0.634 |
| Cond × Elec | F(5,50) = 3.33 | 1.00 | 0.25 | 0.011 * |
| Time × Elec | F(5,50) = 0.50 | 2.42 | 0.047 | 0.604 |
| Cond × Time × Elec | F(5,50) = 0.93 | 0.68 | 0.085 | 0.422 |
| N2 Latency | ||||
| Condition | F(1,10) = 0.85 | 3733.30 | 0.078 | 0.379 |
| Time | F(1,10) = 0.02 | 1372.17 | 0.002 | 0.884 |
| Electrode | F(5,50) = 3.26 | 328.41 | 0.246 | 0.044 * |
| Cond × Time | F(1,10) = 4.89 | 1179.93 | 0.328 | 0.051 |
| Cond × Elec | F(5,50) = 1.25 | 265.30 | 0.11 | 0.310 |
| Time × Elec | F(5,50) = 0.36 | 398.45 | 0.035 | 0.765 |
| Cond × Time × Elec | F(5,50) = 1.42 | 193.25 | 0.125 | 0.259 |
| P3 Amplitude | ||||
| Condition | F(1,10) = 25.98 | 11.59 | 0.722 | <0.001 * |
| Time | F(1,10) = 0.002 | 21.21 | 0.000 | 0.968 |
| Electrode | F(5,50) = 0.99 | 17.98 | 0.090 | 0.384 |
| Cond × Time | F(1,10) = 0.37 | 3.75 | 0.036 | 0.554 |
| Cond × Elec | F(5,50) = 0.91 | 2.06 | 0.084 | 0.427 |
| Time × Elec | F(5,50) = 0.40 | 3.86 | 0.038 | 0.635 |
| Cond × Time × Elec | F(5,50) = 1.75 | 0.82 | 0.149 | 0.185 |
| P3 Latency | ||||
| Condition | F(1,10) = 6.34 | 2481.69 | 0.388 | 0.030 * |
| Time | F(1,10) = 5.99 | 1670.05 | 0.375 | 0.034 * |
| Electrode | F(5,50) = 4.46 | 4965.29 | 0.309 | 0.049 * |
| Cond × Time | F(1,10) =0.09 | 1864.47 | 0.009 | 0.774 |
| Cond × Elec | F(5,50) = 0.72 | 210.94 | 0.067 | 0.561 |
| Time × Elec | F(5,50) = 0.93 | 718.05 | 0.085 | 0.416 |
| Cond × Time × Elec | F(5,50) = 0.48 | 653.62 | 0.046 | 0.646 |
| P3. | Rest | After Exercise | ||
|---|---|---|---|---|
| Latency in ms | Amplitude in µV | Latency in ms | Amplitude in µV | |
| Congruent | 346.59 (20.49) | 10.11 (4.36) | 336.47 (16.87) | 9.76 (4.06) |
| Incongruent | 364.17 (32.41) | 7.91 (3.54) | 350.32 (18.96) | 7.49 (2.62) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winneke, A.H.; Hübner, L.; Godde, B.; Voelcker-Rehage, C. Moderate Cardiovascular Exercise Speeds Up Neural Markers of Stimulus Evaluation During Attentional Control Processes. J. Clin. Med. 2019, 8, 1348. https://doi.org/10.3390/jcm8091348
Winneke AH, Hübner L, Godde B, Voelcker-Rehage C. Moderate Cardiovascular Exercise Speeds Up Neural Markers of Stimulus Evaluation During Attentional Control Processes. Journal of Clinical Medicine. 2019; 8(9):1348. https://doi.org/10.3390/jcm8091348
Chicago/Turabian StyleWinneke, Axel H., Lena Hübner, Ben Godde, and Claudia Voelcker-Rehage. 2019. "Moderate Cardiovascular Exercise Speeds Up Neural Markers of Stimulus Evaluation During Attentional Control Processes" Journal of Clinical Medicine 8, no. 9: 1348. https://doi.org/10.3390/jcm8091348
APA StyleWinneke, A. H., Hübner, L., Godde, B., & Voelcker-Rehage, C. (2019). Moderate Cardiovascular Exercise Speeds Up Neural Markers of Stimulus Evaluation During Attentional Control Processes. Journal of Clinical Medicine, 8(9), 1348. https://doi.org/10.3390/jcm8091348