Emerging Influenza D Virus Threat: What We Know so Far!


Abstract
:1. Introduction
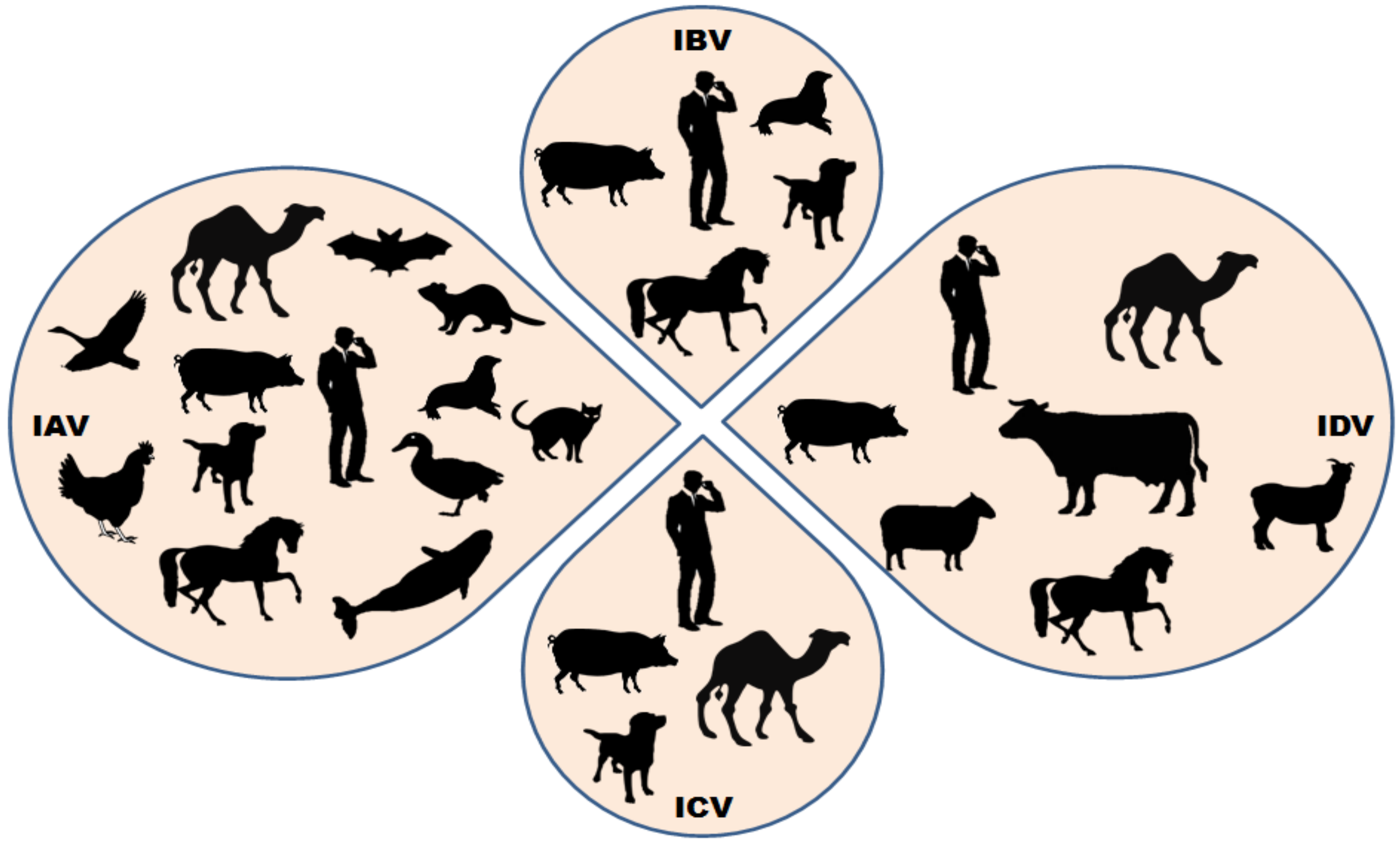
2. Origin, Epidemiology and Pathogenesis of Influenza D Virus
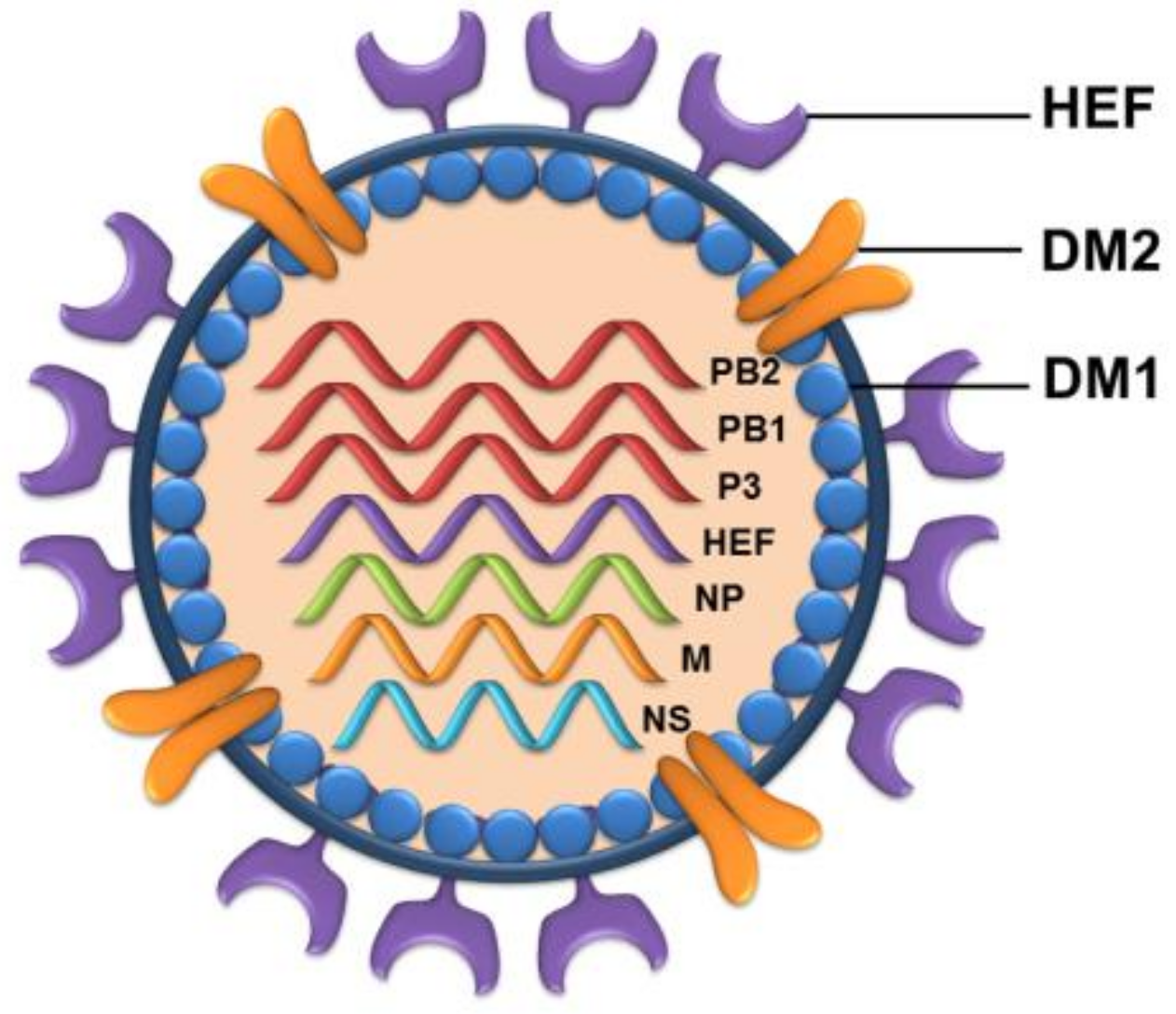
3. Evolution and Genome Structure of Influenza D Virus
4. Influenza D Virus and Public Health Concerns
5. Strategies to Control IDV Infection and Management of Future Threat
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Family—Orthomyxoviridae. In Virus Taxonomy; King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. (Eds.) Elsevier: San Diego, CA, USA, 2012; pp. 749–761. [Google Scholar]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and Swine: Proposal for a new genus in the Orthomyxoviridae family. MBio 2014, 5, e00031–00014. [Google Scholar] [CrossRef] [PubMed]
- Kilbourne, E.D. Taxonomy and Comparative Virology of the Influenza Viruses. In Influenza; Springer US: Boston, MA, USA, 1987; pp. 25–32. [Google Scholar]
- Kumar, B.; Asha, K.; Khanna, M.; Ronsard, L.; Meseko, C.A.; Sanicas, M. The emerging influenza virus threat: Status and new prospects for its therapy and control. Arch. Virol. 2018, 163, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Morens, D.M. 1918 Influenza: The mother of all pandemics. Emerg. Infect. Dis. 2006, 12, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Khanna, M.; Saxena, L.; Gupta, A.; Kumar, B.; Rajput, R. Influenza pandemics of 1918 and 2009: A comparative account. Future Virol. 2013, 8, 335–342. [Google Scholar] [CrossRef]
- Meseko, C.; Kumar, B.; Sanicas, M. Preventing Zoonotic Influenza. In Influenza-Therapeutics and Challenges; Saxena, S.K., Ed.; IntechOpen: London, UK, 2018; pp. 33–55. [Google Scholar]
- Khanna, M.; Kumar, P.; Choudhary, K.; Kumar, B.; Vijayan, V.K. Emerging influenza virus: A global threat. J. Biosci. 2008, 33, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Khanna, M.; Kumar, B.; Gupta, A.; Kumar, P. Pandemic Influenza A H1N1 (2009) Virus: Lessons from the Past and Implications for the Future. Indian J. Virol. 2012, 23, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, M.; Kumar, B.; Gupta, N.; Kumar, P.; Gupta, A.; Vijayan, V.K.; Kaur, H. Pandemic swine influenza virus (H1N1): A threatening evolution. Indian J. Microbiol. 2009, 49, 365–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treanor, J. Influenza Vaccine—Outmaneuvering Antigenic Shift and Drift. N. Engl. J. Med. 2004, 350, 218–220. [Google Scholar] [CrossRef]
- Lakdawala, S.S.; Jayaraman, A.; Halpin, R.A.; Lamirande, E.W.; Shih, A.R.; Stockwell, T.B.; Lin, X.; Simenauer, A.; Hanson, C.T.; Vogel, L.; et al. The soft palate is an important site of adaptation for transmissible influenza viruses. Nature 2015, 526, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Chothe, S.K.; Bhushan, G.; Nissly, R.H.; Yeh, Y.T.; Brown, J.; Turner, G.; Fisher, J.; Sewall, B.J.; Reeder, D.M.; Terrones, M.; et al. Avian and human influenza virus compatible sialic acid receptors in little brown bats. Sci. Rep. 2017, 7, 660. [Google Scholar] [CrossRef]
- Thontiravong, A.; Kitikoon, P.; Wannaratana, S.; Tantilertcharoen, R.; Tuanudom, R.; Pakpinyo, S.; Sasipreeyajan, J.; Oraveerakul, K.; Amonsin, A. Quail as a potential mixing vessel for the generation of new reassortant influenza A viruses. Vet. Microbiol. 2012, 160, 305–313. [Google Scholar] [CrossRef]
- Yoon, S.W.; Webby, R.J.; Webster, R.G. Evolution and ecology of influenza A viruses. Curr. Top. Microbiol. Immunol. 2014, 385, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenstrom, J.; Osterhaus, A.D.; Fouchier, R.A. Global patterns of influenza a virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Watanabe, S.; Maher, E.A.; Neumann, G.; Kawaoka, Y. Pandemic potential of avian influenza A (H7N9) viruses. Trends Microbiol. 2014, 22, 623–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Bi, Y.; Wong, G.; Gray, G.C.; Gao, G.F.; Li, S. Epidemiology, Evolution, and Recent Outbreaks of Avian Influenza Virus in China. J. Virol. 2015, 89, 8671–8676. [Google Scholar] [CrossRef] [Green Version]
- Francis, T., Jr. A New Type of Virus from Epidemic Influenza. Science 1940, 92, 405–408. [Google Scholar] [CrossRef]
- Osterhaus, A.D.; Rimmelzwaan, G.F.; Martina, B.E.; Bestebroer, T.M.; Fouchier, R.A. Influenza B virus in seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef]
- Kawano, J.; Onta, T.; Kida, H.; Yanagawa, R. Distribution of antibodies in animals against influenza B and C viruses. Jpn. J. Vet. Res. 1978, 26, 74–80. [Google Scholar]
- Chang, C.P.; New, A.E.; Taylor, J.F.; Chiang, H.S. Influenza virus isolations from dogs during a human epidemic in Taiwan. Int. J. Zoonoses 1976, 3, 61–64. [Google Scholar]
- Ran, Z.; Shen, H.; Lang, Y.; Kolb, E.A.; Turan, N.; Zhu, L.; Ma, J.; Bawa, B.; Liu, Q.; Liu, H.; et al. Domestic Pigs Are Susceptible to Infection with Influenza B Viruses. J. Virol. 2015, 89, 4818–4826. [Google Scholar] [CrossRef] [Green Version]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- Taylor, R.M. Studies on survival of influenza virus between epidemics and antigenic variants of the virus. Am. J. Public Health Nations Health 1949, 39, 171–178. [Google Scholar] [CrossRef]
- Crescenzo-Chaigne, B.; Barbezange, C.; van der Werf, S. Non coding extremities of the seven influenza virus type C vRNA segments: Effect on transcription and replication by the type C and type A polymerase complexes. Virol. J. 2008, 5, 132. [Google Scholar] [CrossRef]
- Homma, M.; Ohyama, S.; Katagiri, S. Age Distribution of the Antibody to Type C Influenza Virus. Microbiol. Immunol. 1982, 26, 639–642. [Google Scholar] [CrossRef] [Green Version]
- Manuguerra, J.C.; Hannoun, C.; Saenz Mdel, C.; Villar, E.; Cabezas, J.A. Sero-epidemiological survey of influenza C virus infection in Spain. Eur. J. Epidemiol. 1994, 10, 91–94. [Google Scholar] [CrossRef]
- Yuanji, G.; Fengen, J.; Ping, W.; Min, W.; Jiming, Z. Isolation of Influenza C Virus from Pigs and Experimental Infection of Pigs with Influenza C Virus. J. Gen. Virol. 1983, 64, 177–182. [Google Scholar] [CrossRef]
- Ohwada, K.; Kitame, F.; Sugawara, K.; Nishimura, H.; Homma, M.; Nakamura, K. Distribution of the Antibody to Influenza C Virus in Dogs and Pigs in Yamagata Prefecture, Japan. Microbiol. Immunol. 1987, 31, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Elias, S.; Elizabeth, A.J.C.; Hicham Ait, L.; Justine, O.; Félix, A.; Gilbert, L.A.; Emmanuel Couacy, H.; Dishon, M.; Sharon, L.D.; Said, A.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. J. 2017, 23, 1556. [Google Scholar] [CrossRef]
- Bailey, E.S.; Choi, J.Y.; Fieldhouse, J.K.; Borkenhagen, L.K.; Zemke, J.; Zhang, D.; Gray, G.C. The continual threat of influenza virus infections at the human–animal interfaceWhat is new from a one health perspective? Evol. Med. Public Health 2018, 2018, 192–198. [Google Scholar] [CrossRef]
- Vaccines against influenza WHO position paper—November 2012. Wkly. Epidemiol. Rec. 2012, 87, 461–476.
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLOS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef]
- Yuanji, G.; Desselberger, U. Genome Analysis of Influenza C Viruses Isolated in 1981/82 from Pigs in China. J. Gen. Virol. 1984, 65, 1857–1872. [Google Scholar] [CrossRef] [Green Version]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic characterization of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza D virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.-P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.-F. Influenza D virus infection in Mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza D virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Ferguson, L.; Smith, D.R.; Woolums, A.R.; Epperson, W.B.; Wan, X.-F. Serological evidence for high prevalence of Influenza D Viruses in Cattle, Nebraska, United States, 2003–2004. Virology 2017, 501, 88–91. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.-M.; Wang, S.-C.; Peng, C.; Yu, J.-M.; Zhuang, Q.-Y.; Hou, G.-Y.; Liu, S.; Li, J.-P.; Chen, J.-M. Identification of a potential novel type of influenza virus in Bovine in China. Virus Genes 2014, 49, 493–496. [Google Scholar] [CrossRef]
- Shao-Lun, Z.; He, Z.; Sheng-Nan, C.; Xia, Z.; Tao, L.; Runxia, L.; Dian-Hong, L.; Xiao-Hui, W.; Wen-Kang, W.; Dan, W.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. J. 2017, 23, 1392. [Google Scholar] [CrossRef]
- Shin, M.; Maiko, E.; Tomoya, K.; Akiko, T.-U.; James, K.C.; Kazuyuki, U.; Masugi, N.; Benjamin, H.; Taisuke, H. Influenza D Virus Infection in Herd of Cattle, Japan. Emerg. Infect. Dis. J. 2016, 22, 1517. [Google Scholar] [CrossRef]
- Mariette, D.; Claire, P.; Gilles, M. Influenza D Virus in Cattle, France, 2011–2014. Emerg. Infect. Dis. J. 2015, 21, 368. [Google Scholar] [CrossRef]
- Chiara, C.; Silvia, F.; Aurora De, M.; Laura, B.; Ilaria, B.; Carlo, R.; Arrigo, N.; Emanuela, F. Detection of Influenza D Virus among Swine and Cattle, Italy. Emerg. Infect. Dis. J. 2016, 22, 352. [Google Scholar] [CrossRef]
- Foni, E.; Chiapponi, C.; Baioni, L.; Zanni, I.; Merenda, M.; Rosignoli, C.; Kyriakis, C.S.; Luini, M.V.; Mandola, M.L.; Bolzoni, L.; et al. Influenza D in Italy: Towards a better understanding of an emerging viral infection in swine. Sci. Rep. 2017, 7, 11660. [Google Scholar] [CrossRef] [PubMed]
- Orla, F.; Clare, G.; Jean, M.; Claire, I.; Mariette, D.; Ben, H.; Guy, M.; Eoin, R. Influenza D Virus in Cattle, Ireland. Emerg. Infect. Dis. J. 2018, 24, 389. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological evidence for the co-circulation of two lineages of influenza D viruses in equine populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef]
- Horimoto, T.; Hiono, T.; Mekata, H.; Odagiri, T.; Lei, Z.; Kobayashi, T.; Norimine, J.; Inoshima, Y.; Hikono, H.; Murakami, K.; et al. Nationwide Distribution of Bovine Influenza D Virus Infection in Japan. PLoS ONE 2016, 11, e0163828. [Google Scholar] [CrossRef]
- Chantal, J.S.; Justine, O.; Maude, P.; Serge, L.; Félix, W.; Claude, P.M.; Judith, M.H.; Mariette, F.D. Influenza D Virus Circulation in Cattle and Swine, Luxembourg, 2012–2016. Emerg. Infect. Dis. J. 2018, 24, 1388. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, R.; Zhang, L.; Shen, B.; Wang, N.; Xu, Q.; He, W.; He, W.; Li, G.; Su, S. Evolutionary changes of the novel Influenza D virus hemagglutinin-esterase fusion gene revealed by the codon usage pattern. Virulence 2018. [Google Scholar] [CrossRef]
- Lucas, F.; Kaijian, L.; Alicia, K.O.; Fred, L.C.; Sherry, B.; Katie, H.-D.; Hailiang, S.; John, B.; Mark, W.L.; Bianca, Q.; et al. Influenza D Virus Infection in Feral Swine Populations, United States. Emerg. Infect. Dis. J. 2018, 24, 1020. [Google Scholar] [CrossRef]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.-F. Pathogenesis of Influenza D Virus in Cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of Two Distinct Genetic and Antigenic Lineages of Proposed Influenza D Virus in Cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Mekata, H.; Yamamoto, M.; Hamabe, S.; Tanaka, H.; Omatsu, T.; Mizutani, T.; Hause, B.M.; Okabayashi, T. Molecular epidemiological survey and phylogenetic analysis of bovine influenza D virus in Japan. Transbound. Emerg. Dis. 2018, 65, e355–e360. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Velthuis, A.J.; Fodor, E. Influenza virus RNA polymerase: Insights into the mechanisms of viral RNA synthesis. Nat. Rev. Microbiol. 2016, 14, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, S.; Murakami, S.; Shindo, K.; Horimoto, T.; Sagara, H.; Noda, T.; Kawaoka, Y. Influenza C and D Viruses Package Eight Organized Ribonucleoprotein Complexes. J. Virol. 2018, 92, e02084–02017. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. An Open Receptor-Binding Cavity of Hemagglutinin-Esterase-Fusion Glycoprotein from Newly-Identified Influenza D Virus: Basis for Its Broad Cell Tropism. PLOS Pathog. 2016, 12, e1005411. [Google Scholar] [CrossRef]
- Yu, J.; Hika, B.; Liu, R.; Sheng, Z.; Hause, B.M.; Li, F.; Wang, D. The Hemagglutinin-Esterase Fusion Glycoprotein Is a Primary Determinant of the Exceptional Thermal and Acid Stability of Influenza D Virus. mSphere 2017, 2, e00254–00217. [Google Scholar] [CrossRef]
- Noda, T.; Kawaoka, Y. Structure of influenza virus ribonucleoprotein complexes and their packaging into virions. Rev. Med. Virol. 2010, 20, 380–391. [Google Scholar] [CrossRef]
- Kumar, B.; Khanna, M.; Kumar, P.; Sood, V.; Vyas, R.; Banerjea, A.C. Nucleic acid-mediated cleavage of M1 gene of influenza A virus is significantly augmented by antisense molecules targeted to hybridize close to the cleavage site. Mol. Biotechnol. 2012, 51, 27–36. [Google Scholar] [CrossRef]
- Kumar, B.; Kumar, P.; Rajput, R.; Saxena, L.; Daga, M.; Khanna, M. Sequence-specific cleavage of BM2 gene transcript of influenza B virus by 10-23 catalytic motif containing DNA enzymes significantly inhibits viral RNA translation and replication. Nucleic Acid Ther. 2013, 23, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Rajput, R.; Pati, D.R.; Khanna, M. Potent Intracellular Knock-Down of Influenza A Virus M2 Gene Transcript by DNAzymes Considerably Reduces Viral Replication in Host Cells. Mol. Biotechnol. 2015, 57, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Kesinger, E.; Liu, J.; Jensen, A.; Chia, C.P.; Demers, A.; Moriyama, H. Influenza D virus M2 protein exhibits ion channel activity in Xenopus laevis oocytes. PLoS ONE 2018, 13, e0199227. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.; Fodor, E. Emerging Roles for the Influenza A Virus Nuclear Export Protein (NEP). PLoS Pathog. 2012, 8, e1003019. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, D.; Kelvin, D.J.; Li, L.; Zheng, Z.; Yoon, S.-W.; Wong, S.-S.; Farooqui, A.; Wang, J.; Banner, D.; et al. Infectivity, Transmission, and Pathology of Human-Isolated H7N9 Influenza Virus in Ferrets and Pigs. Science 2013, 341, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-L.; Lipatov, A.S.; Ilyushina, N.A.; Govorkova, E.A.; Franks, J.; Yilmaz, N.; Douglas, A.; Hay, A.; Krauss, S.; Rehg, J.E.; et al. Inefficient Transmission of H5N1 Influenza Viruses in a Ferret Contact Model. J. Virol. 2007, 81, 6890–6898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maines, T.R.; Chen, L.-M.; Matsuoka, Y.; Chen, H.; Rowe, T.; Ortin, J.; Falcón, A.; Hien, N.T.; Mai, L.Q.; Sedyaningsih, E.R.; et al. Lack of transmission of H5N1 avian–human reassortant influenza viruses in a ferret model. Proc. Natl. Acad. Sci. USA 2006, 103, 12121–12126. [Google Scholar] [CrossRef]
- Munster, V.J.; de Wit, E.; van den Brand, J.M.A.; Herfst, S.; Schrauwen, E.J.A.; Bestebroer, T.M.; van de Vijver, D.; Boucher, C.A.; Koopmans, M.; Rimmelzwaan, G.F.; et al. Pathogenesis and Transmission of Swine-Origin 2009 A(H1N1) Influenza Virus in Ferrets. Science 2009, 325, 481–483. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.B.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and Transmission of the Novel Bovine Influenza D Virus in a Guinea Pig Model. J. Virol. 2015, 89, 11990–12001. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Gaunt, E.R.; Digard, P.; Templeton, K.; Simmonds, P. Detection of influenza C virus but not influenza D virus in Scottish respiratory samples. J. Clin. Virol. 2016, 74, 50–53. [Google Scholar] [CrossRef] [Green Version]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza D virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Faccini, S.; De Mattia, A.; Chiapponi, C.; Barbieri, I.; Boniotti, M.B.; Rosignoli, C.; Franzini, G.; Moreno, A.; Foni, E.; Nigrelli, A.D. Development and evaluation of a new Real-Time RT-PCR assay for detection of proposed influenza D virus. J. Virol. Methods 2017, 243, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, M.; Tsuchiaka, S.; Rahpaya, S.S.; Hasebe, A.; Otsu, K.; Sugimura, S.; Kobayashi, S.; Komatsu, N.; Nagai, M.; Omatsu, T.; et al. Development of a one-run real-time PCR detection system for pathogens associated with bovine respiratory disease complex. J. Vet. Med. Sci. 2017, 79, 517–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henritzi, D.; Hoffmann, B.; Wacheck, S.; Pesch, S.; Herrler, G.; Beer, M.; Harder, T.C. A newly developed tetraplex real-time RT-PCR for simultaneous screening of influenza virus types A, B, C and D. Influ. Other Respir. Viruses 2019, 13, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Lelli, D.; Lavazza, A.; Sozzi, E.; Zanni, I.; Chiapponi, C.; Foni, E.; Capucci, L.; Brocchi, E. MAb-based competitive ELISA for the detection of antibodies against influenza D virus. Transbound. Emerg. Dis. 2019, 66, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.D.; Ma, M.; Xia, Y.; Wang, T.; Shu, B.; Lednicky, J.A.; Ma, M.-J.; Lu, J.; Gray, G.C. Bioaerosol Sampling in Modern Agriculture: A Novel Approach for Emerging Pathogen Surveillance? J. Infect. Dis. 2016, 214, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, E.S.; Choi, J.Y.; Zemke, J.; Yondon, M.; Gray, G.C. Molecular surveillance of respiratory viruses with bioaerosol sampling in an airport. Trop. Dis. Travel Med. Vaccines 2018, 4, 11. [Google Scholar] [CrossRef]
- Kumar, P.; Khanna, M.; Kumar, B.; Rajput, R.; Banerjea, A.C. A conserved matrix epitope based DNA vaccine protects mice against influenza A virus challenge. Antiviral Res. 2012, 93, 78–85. [Google Scholar] [CrossRef]
- Wan, Y.; Kang, G.; Sreenivasan, C.; Daharsh, L.; Zhang, J.; Fan, W.; Wang, D.; Moriyama, H.; Li, F.; Li, Q. A DNA Vaccine Expressing Consensus Hemagglutinin-Esterase Fusion Protein Protected Guinea Pigs from Infection by Two Lineages of Influenza D Virus. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Hause, B.M.; Huntimer, L.; Falkenberg, S.; Henningson, J.; Lechtenberg, K.; Halbur, T. An inactivated influenza D virus vaccine partially protects cattle from respiratory disease caused by homologous challenge. Vet. Microbiol. 2017, 199, 47–53. [Google Scholar] [CrossRef]
- Easterday, B.C.; Trainer, D.O.; TŮMovÁ, B.; Pereira, H.G. Evidence of Infection with Influenza Viruses in Migratory Waterfowl. Nature 1968, 219, 523–524. [Google Scholar] [CrossRef]
- Webby, R.J.; Webster, R.G.; Richt, J.A. Influenza viruses in animal wildlife populations. Curr. Top. Microbiol. Immunol. 2007, 315, 67–83. [Google Scholar] [PubMed]
- Krauss, S.; Walker, D.; Pryor, S.P.; Niles, L.; Chenghong, L.; Hinshaw, V.S.; Webster, D.R.G. Influenza A Viruses of Migrating Wild Aquatic Birds in North America. Vector-Borne Zoonotic Dis. 2004, 4, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Stallknecht, D.E.; Slemons, R.D.; Winker, K.; Suarez, D.L.; Scott, M.; Swayne, D.E. Phylogenetic analyses of type A influenza genes in natural reservoir species in North America reveals genetic variation. Virus Res. 2005, 114, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Munster, V.J.; Baas, C.; Lexmond, P.; Waldenstrom, J.; Wallensten, A.; Fransson, T.; Rimmelzwaan, G.F.; Beyer, W.E.; Schutten, M.; Olsen, B.; et al. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog. 2007, 3, e61. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D.E.; Shane, S.M. Host range of avian influenza virus in free-living birds. Vet. Res. Commun. 1988, 12, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Feare, C.J.; Yasue, M. Asymptomatic infection with highly pathogenic avian influenza H5N1 in wild birds: How sound is the evidence? Virol. J. 2006, 3, 96. [Google Scholar] [CrossRef] [PubMed]
- Causey, D.; Edwards, S.V. Ecology of Avian Influenza Virus in Birds. J. Infect. Dis. 2008, 197, S29–S33. [Google Scholar] [CrossRef]
- Normile, D. Evidence Points to Migratory Birds in H5N1 Spread. Science 2006, 311, 1225–1225. [Google Scholar] [CrossRef]
- Zhou, N.N.; Shortridge, K.F.; Claas, E.C.J.; Krauss, S.L.; Webster, R.G. Rapid Evolution of H5N1 Influenza Viruses in Chickens in Hong Kong. J. Virol. 1999, 73, 3366–3374. [Google Scholar]
- Asha, K.; Kumar, P.; Sanicas, M.; Meseko, C.; Khanna, M.; Kumar, B. Advancements in Nucleic Acid Based Therapeutics against Respiratory Viral Infections. J. Clin. Med. 2018, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Khanna, M.; Saxena, L.; Rajput, R.; Kumar, B.; Prasad, R. Gene silencing: A therapeutic approach to combat influenza virus infections. Future Microbiol. 2015, 10, 131–140. [Google Scholar] [CrossRef]
- Kumar, B.; Asha, K.; Chauhan, S. DNAzyme mediated post-transcriptional gene silencing: A novel therapeutic approach. WebmedCentral Mol. Biol. 2013, 4, WMC004415. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, B.; Rajput, R.; Saxena, L.; Banerjea, A.C.; Khanna, M. Cross-protective effect of antisense oligonucleotide developed against the common 3′ NCR of influenza A virus genome. Mol. Biotechnol. 2013, 55, 203–211. [Google Scholar] [CrossRef]
- Rajput, R.; Khanna, M.; Kumar, P.; Kumar, B.; Sharma, S.; Gupta, N.; Saxena, L. Small interfering RNA targeting the nonstructural gene 1 transcript inhibits influenza A virus replication in experimental mice. Nucleic Acid Ther. 2012, 22, 414–422. [Google Scholar] [CrossRef]
- Motard, J.; Rouxel, R.; Paun, A.; von Messling, V.; Bisaillon, M.; Perreault, J.P. A novel ribozyme-based prophylaxis inhibits influenza A virus replication and protects from severe disease. PLoS ONE 2011, 6, e27327. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, S.; Wang, S. DNAzymes Dz13 target the c-jun possess antiviral activity against influenza A viruses. Microb. Pathog. 2017, 103, 155–161. [Google Scholar] [CrossRef]
- Tang, X.B.; Hobom, G.; Luo, D. Ribozyme mediated destruction of influenza A virus in vitro and in vivo. J. Med. Virol. 1994, 42, 385–395. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genus | Species | Number of Genomic Segments |
|---|---|---|
| Alphainfluenzavirus | Influenza A virus | 8 |
| Betainfluenzavirus | Influenza B virus | 8 |
| Gammainfluenzavirus | Influenza C virus | 7 |
| Deltainfluenzavirus | Influenza D virus | 7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asha, K.; Kumar, B. Emerging Influenza D Virus Threat: What We Know so Far! J. Clin. Med. 2019, 8, 192. https://doi.org/10.3390/jcm8020192
Asha K, Kumar B. Emerging Influenza D Virus Threat: What We Know so Far! Journal of Clinical Medicine. 2019; 8(2):192. https://doi.org/10.3390/jcm8020192
Chicago/Turabian StyleAsha, Kumari, and Binod Kumar. 2019. "Emerging Influenza D Virus Threat: What We Know so Far!" Journal of Clinical Medicine 8, no. 2: 192. https://doi.org/10.3390/jcm8020192