Metabolic Fingerprint of Acromegaly and Its Potential Usefulness in Clinical Practice

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Ethics Statement
2.2. Design and Patients
2.3. Laboratory Measurements and Phenotypic Characterization
2.4. Preparation of Plasma Samples for Metabolomics Analysis
2.5. Metabolic Profiling by Nuclear Magnetic Resonance (NMR)
2.6. Statistical Analysis
2.6.1. General Comparisons among Groups
2.6.2. Metabolomic Data
3. Results
3.1. Patients
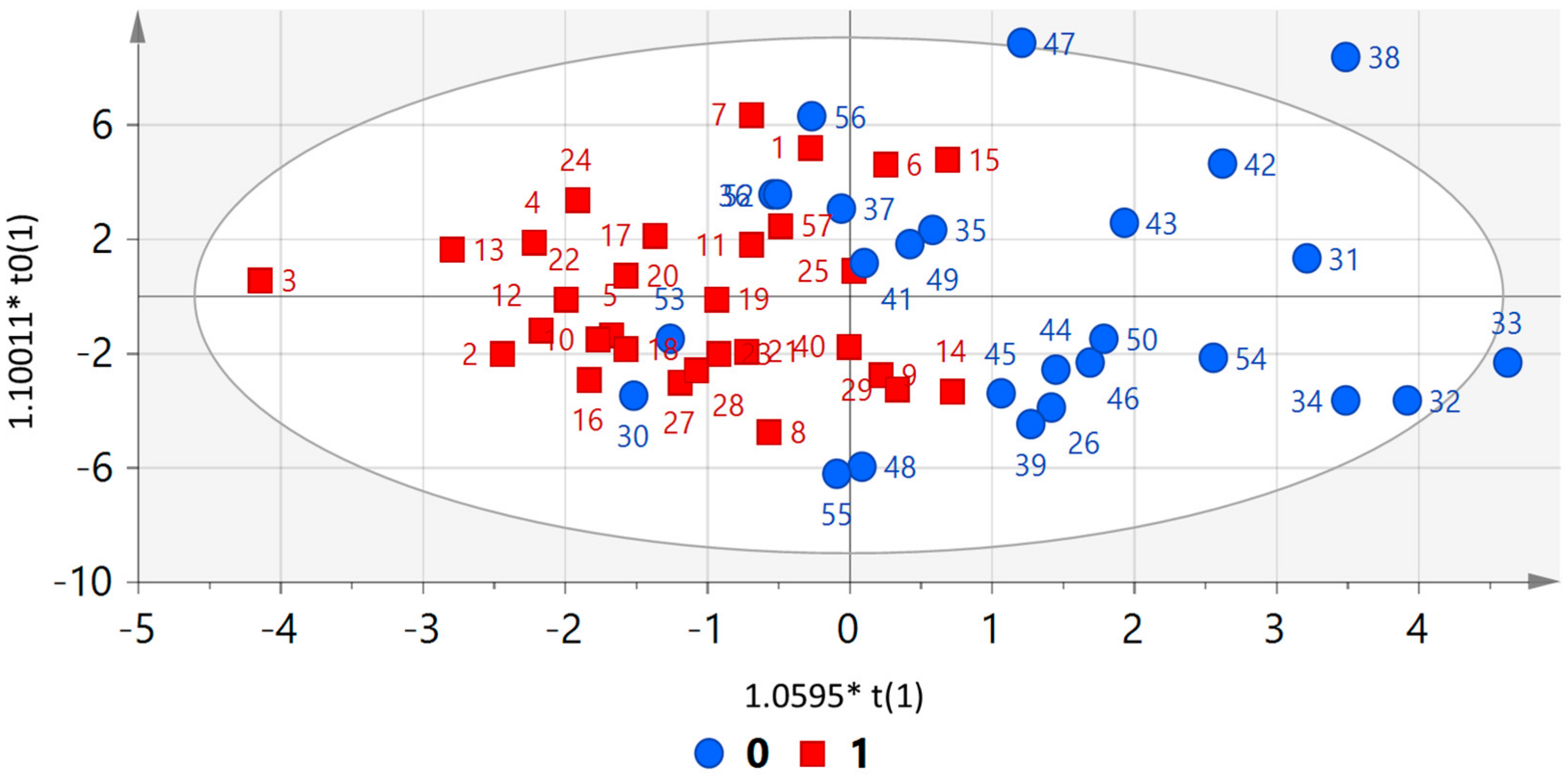
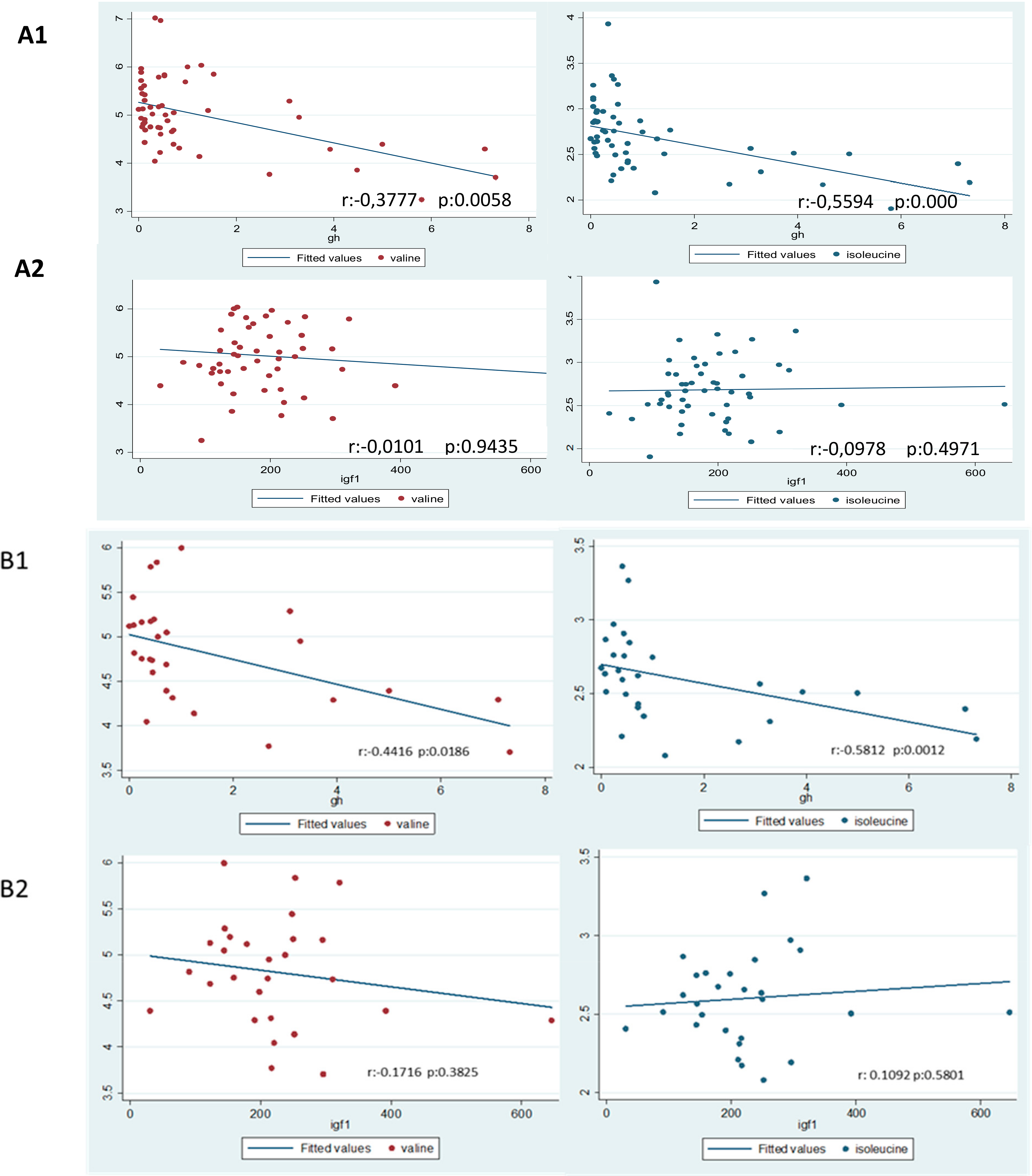
3.2. Metabolomic Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Melmed, S. Acromegaly pathogenesis and treatment. J. Clin. Invest. 2009, 119, 3189–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colao, A.; Ferone, D.; Marzullo, P.; Lombardi, G. Systemic complications of acromegaly: Epidemiology, pathogenesis, and management. Endocr. Rev. 2004, 25, 102–152. [Google Scholar] [CrossRef] [PubMed]
- Melmed, S. Medical progress: Acromegaly. N. Engl. J. Med. 2006, 355, 2558–2573. [Google Scholar] [CrossRef]
- Esposito, D.; Ragnarsson, O.; Granfeldt, D.; Marlow, T.; Johannsson, G.; Olsson, D.S. Decreasing mortality and changes in treatment patterns in patients with acromegaly from a nationwide study. Eur. J. Endocrinol. 2018, 178, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arosio, M.; Reimondo, G.; Malchiodi, E.; Berchialla, P.; Borraccino, A.; Marinis, L.D.; Pivonello, R.; Grottoli, S.; Losa, M.; Cannavò, S.; et al. Predictors of morbidity and mortality in acromegaly: An Italian survey. Eur. J. Endocrinol. 2012, 167, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Ritvonen, E.; Löyttyniemi, E.; Jaatinen, P.; Ebeling, T.; Moilanen, L.; Nuutila, P.; Kauppinen-Mäkelin, R.; Schalin-Jäntti, C. Mortality in acromegaly: A 20-year follow-up study. Endocr. Relat. Cancer 2016, 23, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Varadhan, L.; Reulen, R.C.; Brown, M.; Clayton, R.N. The role of cumulative growth hormone exposure in determining mortality and morbidity in acromegaly: A single centre study. Pituitary 2016, 19, 251–261. [Google Scholar] [CrossRef]
- Rajasoorya, C.; Holdaway, I.M.; Wrightson, P.; Scott, D.J.; Ibbertson, H.K. Determinants of clinical outcome and survival in acromegaly. Clin. Endocrinol. (Oxf.) 1994, 41, 95–102. [Google Scholar] [CrossRef]
- Orme, S.M.; McNally, R.J.; Cartwright, R.A.; Belchetz, P.E. United Kingdom Acromegaly Study Group. Mortality and cancer incidence in acromegaly: A retrospective cohort study. J. Clin. Endocrinol. Metab. 1998, 83, 2730–2734. [Google Scholar] [PubMed]
- Holdaway, I.M.; Rajasoorya, R.C.; Gamble, G.D. Factors influencing mortality in acromegaly. J. Clin. Endocrinol. Metab. 2004, 89, 667–674. [Google Scholar] [CrossRef]
- Mercado, M.; Gonzalez, B.; Vargas, G.; Ramirez, C.; de los Monteros, A.L.E.; Sosa, E.; Jervis, P.; Roldan, P.; Mendoza, V.; López-Félix, B.; et al. Successful mortality reduction and control of comorbidities in patients with acromegaly followed at a highly specialized multidisciplinary clinic. J. Clin. Endocrinol. Metab. 2014, 99, 4438–4446. [Google Scholar] [CrossRef] [PubMed]
- Sardella, C.; Cappellani, D.; Urbani, C.; Manetti, L.; Marconcini, G.; Tomisti, L.; Lupi, I.; Rossi, G.; Scattina, I.; Lombardi, M.; et al. Disease activity and lifestyle influence comorbidities and cardiovascular events in patients with acromegaly. Eur. J. Endocrinol. 2016, 175, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Schwartzbauer, G.; Sperling, M.A.; Devaskar, S.U.; Thamotharan, S.; Robbins, P.D.; McTiernan, C.F.; Liu, J.L.; Jiang, J.; Frank, S.J.; et al. Demonstration of direct effects of growth hormone on neonatal cardiomyocytes. J. Biol. Chem. 2001, 276, 22892–22900. [Google Scholar] [CrossRef] [PubMed]
- Isgaard, J.; Arcopinto, M.; Karason, K.; Cittadini, A. GH and the cardiovascular system: An update on a topic at heart. Endocrine 2015, 48, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.M. Quality of life in acromegaly. Neuroendocrinology 2006, 83, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Bevan, J.S.; Bronstein, M.D.; Casanueva, F.F.; Chanson, P.; Petersenn, S.; Thanh, X.-M.T.; Sert, C.; Houchard, A.; Guillemin, I.; et al. SAGIT®: Clinician-reported outcome instrument for managing acromegaly in clinical practice—development and results from a pilot study. Pituitary 2016, 19, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Van der Lely, A.J.; Gomez, R.; Pleil, A.; Badia, X.; Brue, T.; Buchfelder, M.; Burman, P.; Clemmons, D.; Ghigo, E.; Jørgensen, J.O.L.; et al. Development of ACRODAT®, a new software medical device to assess disease activity in patients with acromegaly. Pituitary 2017, 20, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef]
- Cuschieri, S. The STROBE guidelines. Saudi J. Anaesth. 2019, 13, S31–S34. [Google Scholar] [CrossRef]
- Katznelson, L.; Laws, E.R.; Melmed, S.; Molitch, M.E.; Murad, M.H.; Utz, A.; Wass, J.A.H. Acromegaly: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2014, 99, 3933–3951. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, E.; Solis-Herrera, C.; Trautmann, M.E.; Malloy, J.; Triplitt, C.L. Assessment of Pancreatic β-Cell Function: Review of Methods and Clinical Applications. Curr. Diabetes Rev. 2014, 10, 2–42. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of HOMA modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Ulrich, E.L.; Akutsu, H.; Doreleijers, J.F.; Harano, Y.; Ioannidis, Y.E.; Lin, J.; Livny, M.; Mading, S.; Maziuk, D.; Miller, Z.; et al. BioMagResBank. Nucleic Acids Res. 2008, 36, D402–D408. [Google Scholar] [CrossRef] [PubMed]
- Knacke, H.; Pietzner, M.; Do, K.T.; Römisch-Margl, W.; Kastenmüller, G.; Völker, U.; Völzke, H.; Krumsiek, J.; Artati, A.; Wallaschofski, H.; et al. Metabolic Fingerprints of Circulating IGF-1 and the IGF-1/IGFBP-3 Ratio: A Multifluid Metabolomics Study. J. Clin. Endocrinol. Metab. 2016, 101, 4730–4742. [Google Scholar] [CrossRef]
- Fuchs, S.; Bundy, J.G.; Davies, S.K.; Viney, J.M.; Swire, J.S.; Leroi, A.M. A metabolic signature of long life in Caenorhabditis elegans. BMC Biol. 2010, 8, 14. [Google Scholar] [CrossRef]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2018. [Google Scholar] [CrossRef]
- Mastrangelo, A.; Martos-Moreno, G.Á.; Rupérez, F.J.; Chowen, J.A.; Barbas, C.; Argente, J. Metabolomics changes in patients with PAPP-A2 deficiency in response to rhIGF1 treatment. Growth Horm. IGF Res. 2018, 42, 28–31. [Google Scholar] [CrossRef]
- Berger, M.; Zimmermann-Telschow, H.; Berchtold, P.; Drost, H.; Müller, W.A.; Gries, F.A.; Zimmermann, H. Blood amino acid levels in patients with insulin excess (functioning insulinoma) and insulin deficiency (diabetic ketosis). Metabolism 1978, 27, 793–799. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Koulman, A.; Griffin, J.L. Towards metabolic biomarkers of insulin resistance and type 2 diabetes: Progress from the metabolome. Lancet Diabetes Endocrinol. 2014, 2, 65–75. [Google Scholar] [CrossRef]
- Ruiz-Canela, M.; Guasch-Ferré, M.; Toledo, E.; Clish, C.B.; Razquin, C.; Liang, L.; Wang, D.D.; Corella, D.; Estruch, R.; Hernáez, Á.; et al. Plasma branched chain/aromatic amino acids, enriched Mediterranean diet and risk of type 2 diabetes: Case-cohort study within the PREDIMED Trial. Diabetologia 2018, 61, 1560–1571. [Google Scholar] [CrossRef] [PubMed]
- Kahl, S.; Roden, M. Amino acids—lifesaver or killer in patients with diabetes? Nat. Rev. Endocrinol. 2018, 14, 449–451. [Google Scholar] [CrossRef]
- Gancheva, S.; Jelenik, T.; Álvarez-Hernández, E.; Roden, M. Interorgan Metabolic Crosstalk in Human Insulin Resistance. Physiol. Rev. 2018, 98, 1371–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colao, A.; Auriemma, R.S.; Savastano, S.; Galdiero, M.; Grasso, L.F.S.; Lombardi, G.; Pivonello, R. Glucose Tolerance and Somatostatin Analog Treatment in Acromegaly: A 12-Month Study. J. Clin. Endocrinol. Metab. 2009, 94, 2907–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coculescu, M.; Niculescu, D.; Lichiardopol, R.; Purice, M. Insulin resistance and insulin secretion in non-diabetic acromegalic patients. Exp. Clin. Endocrinol. Diabetes 2007, 115, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, C.L.; Orsi, E.; Giavoli, C.; Cappiello, V.; Epaminonda, P.; Beck-Peccoz, P.; Arosio, M. Evaluation of insulin resistance in acromegalic patients before and after treatment with somatostatin analogues. J. Endocrinol. Invest. 2003, 26, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Kasayama, S.; Otsuki, M.; Takagi, M.; Saito, H.; Sumitani, S.; Kouhara, H.; Koga, M.; Saitoh, Y.; Ohnishi, T.; Arita, N. Impaired beta-cell function in the presence of reduced insulin sensitivity determines glucose tolerance status in acromegalic patients. Clin. Endocrinol. (Oxf.) 2000, 52, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Møller, N.; Schmitz, O.; Jøorgensen, J.O.; Astrup, J.; Bak, J.F.; Christensen, S.E.; Alberti, K.G.; Weeke, J. Basal- and insulin-stimulated substrate metabolism in patients with active acromegaly before and after adenomectomy. J. Clin. Endocrinol. Metab. 1992, 74, 1012–1019. [Google Scholar] [PubMed]
- Cozzolino, A.; Feola, T.; Simonelli, I.; Puliani, G.; Pozza, C.; Giannetta, E.; Gianfrilli, D.; Pasqualetti, P.; Lenzi, A.; Isidori, A.M. Somatostatin Analogs and Glucose Metabolism in Acromegaly: A Meta-analysis of Prospective Interventional Studies. J. Clin. Endocrinol. Metab. 2018, 103, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Tzanela, M.; Vassiliadi, D.A.; Gavalas, N.; Szabo, A.; Margelou, E.; Valatsou, A.; Vassilopoulos, C. Glucose homeostasis in patients with acromegaly treated with surgery or somatostatin analogues. Clin. Endocrinol. 2011, 75, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, G.; Floriani, I.; Bonadonna, S.; Torri, V.; Chanson, P.; Giustina, A. Effects of somatostatin analogs on glucose homeostasis: A metaanalysis of acromegaly studies. J. Clin. Endocrinol. Metab. 2009, 94, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.K.; Jenkins, A.B.; Furler, S.M.; Borkman, M.; Chisholm, D.J. Impact of octreotide, a long-acting somatostatin analogue, on glucose tolerance and insulin sensitivity in acromegaly. Clin. Endocrinol. (Oxf.) 1992, 36, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Giustina, G.; Peracchi, M.; Reschini, E.; Panerai, E.; Pinto, M. Dose-response study of the inhibiting effect of somatostatin on growth hormone and insulin secretion in normal subjects and acromegalic patients. Metab. Clin. Exp. 1975, 24, 807–815. [Google Scholar] [CrossRef]
- Feola, T.; Cozzolino, A.; Simonelli, I.; Sbardella, E.; Pozza, C.; Giannetta, E.; Gianfrilli, D.; Pasqualetti, P.; Lenzi, A.; Isidori, A.M. Pegvisomant Improves Glucose Metabolism in Acromegaly: A Meta-Analysis of Prospective Interventional Studies. J. Clin. Endocrinol. Metab. 2019, 104, 2892–2902. [Google Scholar] [CrossRef]
- Olarescu, N.C.; Heck, A.; Godang, K.; Ueland, T.; Bollerslev, J. The Metabolic Risk in Patients Newly Diagnosed with Acromegaly Is Related to Fat Distribution and Circulating Adipokines and Improves after Treatment. Neuroendocrinology 2016, 103, 197–206. [Google Scholar] [CrossRef]
- Olarescu, N.C.; Bollerslev, J. The Impact of Adipose Tissue on Insulin Resistance in Acromegaly. Trends Endocrinol. Metab. 2016, 27, 226–237. [Google Scholar] [CrossRef]
- Briet, C.; Ilie, M.D.; Kuhn, E.; Maione, L.; Brailly-Tabard, S.; Salenave, S.; Cariou, B.; Chanson, P. Changes in metabolic parameters and cardiovascular risk factors after therapeutic control of acromegaly vary with the treatment modality. Data from the Bicêtre cohort, and review of the literature. Endocrine 2019, 63, 348–360. [Google Scholar] [CrossRef]
- Reid, T.J.; Jin, Z.; Shen, W.; Reyes-Vidal, C.M.; Fernandez, J.C.; Bruce, J.N.; Kostadinov, J.; Post, K.D.; Freda, P.U. IGF-1 levels across the spectrum of normal to elevated in acromegaly: Relationship to insulin sensitivity, markers of cardiovascular risk and body composition. Pituitary 2015, 18, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Urbani, C.; Sardella, C.; Calevro, A.; Rossi, G.; Scattina, I.; Lombardi, M.; Lupi, I.; Manetti, L.; Martino, E.; Bogazzi, F. Effects of medical therapies for acromegaly on glucose metabolism. Eur. J. Endocrinol. 2013, 169, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fryburg, D.A.; Gelfand, R.A.; Barrett, E.J. Growth hormone acutely stimulates forearm muscle protein synthesis in normal humans. Am. J. Physiol. 1991, 260, E499–E504. [Google Scholar] [CrossRef] [PubMed]
- Garibotto, G. Muscle amino acid metabolism and the control of muscle protein turnover in patients with chronic renal failure. Nutrition 1999, 15, 145–155. [Google Scholar] [CrossRef]
- Fryburg, D.A.; Louard, R.J.; Gerow, K.E.; Gelfand, R.A.; Barrett, E.J. Growth hormone stimulates skeletal muscle protein synthesis and antagonizes insulin’s antiproteolytic action in humans. Diabetes 1992, 41, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Horber, F.F.; Haymond, M.W. Human growth hormone prevents the protein catabolic side effects of prednisone in humans. J. Clin. Invest. 1990, 86, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Nørrelund, H.; Nair, K.S.; Jørgensen, J.O.L.; Christiansen, J.S.; Møller, N. The Protein-Retaining Effects of Growth Hormone During Fasting Involve Inhibition of Muscle-Protein Breakdown. Diabetes 2001, 50, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, R.; Nishida, H.; Yamamoto, Y.; Wake, I.; Kai, N.; Kikkawa, U.; Okimura, Y. Growth hormone and Insulin-like growth factor-I (IGF-I) modulate the expression of L-type amino acid transporters in the muscles of spontaneous dwarf rats and L6 and C2C12 myocytes. Growth Horm. IGF Res. 2018, 42, 66–73. [Google Scholar] [CrossRef]
- Sciascia, Q.L.; Pacheco, D.; McCoard, S.A. Administration of Exogenous Growth Hormone Is Associated with Changes in Plasma and Intracellular Mammary Amino Acid Profiles and Abundance of the Mammary Gland Amino Acid Transporter SLC3A2 in Mid-Lactation Dairy Cows. PLoS ONE 2015, 10, e0134323. [Google Scholar] [CrossRef]
- Møller, N.; Jørgensen, J.O.L. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocr. Rev. 2009, 30, 152–177. [Google Scholar] [CrossRef]
- Füchtbauer, L.; Olsson, D.S.; Bengtsson, B.-Å.; Norrman, L.-L.; Sunnerhagen, K.S.; Johannsson, G. Muscle strength in patients with acromegaly at diagnosis and during long-term follow-up. Eur. J. Endocrinol. 2017, 177, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Doll, H.; David, J.; Wass, J. Impact of musculoskeletal disease on quality of life in long-standing acromegaly. Eur. J. Endocrinol. 2008, 158, 587–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; He, H.; Fang, S.; Hua, Y.; Yang, X.; Yuan, Y.; Liang, S.; Liu, P.; Tian, Y.; Xu, F.; et al. Time Series Characteristics of Serum Branched-Chain Amino Acids for Early Diagnosis of Chronic Heart Failure. J. Proteome Res. 2019, 18, 2121–2128. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Z.; Kolwicz, S.C.; Abell, L.; Roe, N.D.; Kim, M.; Zhou, B.; Cao, Y.; Ritterhoff, J.; Gu, H.; et al. Defective Branched-Chain Amino Acid (BCAA) Catabolism Disrupts Glucose Metabolism and Sensitizes the Heart to Ischemia-reperfusion Injury. Cell Metab. 2017, 25, 374–385. [Google Scholar] [CrossRef]
- Akutsu, H.; Kreutzer, J.; Wasmeier, G.; Ropers, D.; Rost, C.; Möhlig, M.; Wallaschofski, H.; Buchfelder, M.; Schöfl, C. Acromegaly per se does not increase the risk for coronary artery disease. Eur. J. Endocrinol. 2010, 162, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Aulinas, A.; Crespo, I.; Viladés, D.; Leta, R.; Urgell, E.; Biagetti, B.; Webb, S.M.; Valassi, E. Cystatin-C and epicardial adipose tissue as noninvasive predictors of cardiovascular risk in acromegaly. Clin. Endocrinol. (Oxf.) 2017, 86, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Bogazzi, F.; Battolla, L.; Spinelli, C.; Rossi, G.; Gavioli, S.; Bello, V.D.; Cosci, C.; Sardella, C.; Volterrani, D.; Talini, E.; et al. Risk Factors for Development of Coronary Heart Disease in Patients with Acromegaly: A Five-Year Prospective Study. J. Clin. Endocrinol. Metab. 2007, 92, 4271–4277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannavo, S.; Almoto, B.; Cavalli, G.; Squadrito, S.; Romanello, G.; Vigo, M.T.; Fiumara, F.; Benvenga, S.; Trimarchi, F. Acromegaly and Coronary Disease: An Integrated Evaluation of Conventional Coronary Risk Factors and Coronary Calcifications Detected by Computed Tomography. J. Clin. Endocrinol. Metab. 2006, 91, 3766–3772. [Google Scholar] [CrossRef]
- Dong, W.; Zhou, M.; Dong, M.; Pan, B.; Liu, Y.; Shao, J.; Gu, X.; Huang, Y.; Li, G.; Wang, Y.; et al. Keto acid metabolites of branched-chain amino acids inhibit oxidative stress-induced necrosis and attenuate myocardial ischemia-reperfusion injury. J. Mol. Cell. Cardiol. 2016, 101, 90–98. [Google Scholar] [CrossRef]
- Ra, S.-G.; Miyazaki, T.; Kojima, R.; Komine, S.; Ishikura, K.; Kawanaka, K.; Honda, A.; Matsuzaki, Y.; Ohmori, H. Effect of BCAA supplement timing on exercise-induced muscle soreness and damage: A pilot placebo-controlled double-blind study. J. Sports Med. Phys. Fit. 2018, 58, 1582–1591. [Google Scholar] [CrossRef]
- VanDusseldorp, T.A.; Escobar, K.A.; Johnson, K.E.; Stratton, M.T.; Moriarty, T.; Cole, N.; McCormick, J.J.; Kerksick, C.M.; Vaughan, R.A.; Dokladny, K.; et al. Effect of Branched-Chain Amino Acid Supplementation on Recovery Following Acute Eccentric Exercise. Nutrients 2018, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, A.; Michitaka, K.; Kiguchi, D.; Izumoto, H.; Ueki, H.; Kaneto, M.; Kitahata, S.; Aibiki, T.; Okudaira, T.; Tomida, H.; et al. Efficacy of branched-chain amino acid supplementation and walking exercise for preventing sarcopenia in patients with liver cirrhosis. Eur. J. Gastroenterol. Hepatol. 2017, 29, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, Y.; Takahashi, H.; Akiyama, T.; Murayama, K.; Iwane, S.; Kuwashiro, T.; Tanaka, K.; Kawazoe, S.; Ono, N.; Eguchi, T.; et al. Supplementation with branched-chain amino acids ameliorates hypoalbuminemia, prevents sarcopenia, and reduces fat accumulation in the skeletal muscles of patients with liver cirrhosis. J. Gastroenterol. 2018, 53, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Kitaura, Y.; Kadota, Y.; Morishita, Y.; Ota, M.; Yamanaka, F.; Xu, M.; Ikawa, M.; Inoue, N.; Kawano, F.; et al. Muscle-specific deletion of BDK amplifies loss of myofibrillar protein during protein undernutrition. Sci. Rep. 2017, 7, 39825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Kitaura, Y.; Ishikawa, T.; Kadota, Y.; Terai, C.; Shindo, D.; Morioka, T.; Ota, M.; Morishita, Y.; Ishihara, K.; et al. Endurance performance and energy metabolism during exercise in mice with a muscle-specific defect in the control of branched-chain amino acid catabolism. PLoS ONE 2017, 12, e0180989. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cases (n = 30) | Controls (n = 30) | p-Value | |
|---|---|---|---|
| Gender male (n, %) | 16 (53.3%) | 16 (53.3%) | 0.60 |
| Age years (mean +/- SD) | 59.5 ± 11.3 | 56.3 ± 11. | 0.22 |
| BMI kg/m2 (mean ± SD) | 29.57 ± 5.7 | 27.3 ± 5.1 | 0.08 |
| Smoking (n, %) | 5 (16.7%) | 5 (16.7%) | 0.95 |
| Diabetes (n, %) | 7 (23.3%) | 6 (20.0%) | 0.09 |
| Hypertension (n, %) | 8 (26.7%) | 11 (36.7%) | 0.58 |
| Dyslipidemia (n, %) | 11 (36.7%) | 14 (46.7%) | 0.60 |
| HOMA-IR p50 (IQR) | 1.9 (2.8) | 3.0 (2.6) | 0.06 |
| HOMA-B p50 (IQR) | 131.3 (137.1) | 164.3 (92.0) | 0.25 |
| GH p50 (IQR) | 0.63 (2.69) | 0.31 (0.71) | 0.03 |
| IGF-1 p50 (IQR) | 216.1 ± 192 | 143.0 ± 58 | 0.00 |
| ACRO_Active (n = 5) | ACRO_Medical (n = 14) | ACRO_Remission (n = 11) | p | |
|---|---|---|---|---|
| Gender Male (n, %) | 2 (40.0%) | 9 (64.0%) | 5 (45.0%) | 0.55 |
| Age years (mean +/- SD) | 55.4 ± 9.2 | 64.3 ± 11.1 | 55.4 ± 10.9 | 0.10 |
| BMI kg/m2 (mean ± SD) | 29.7 ± 7.4 | 27.6 ± 4.1 | 32.1 ± 6.1 | 0.19 |
| Smoking (n, %) | 2 (40.0%) | 2 (14.3%) | 1 (9.1%) | 0.28 |
| Diabetes (n, %) | 2 (40.0%) | 5 (37.7%) | 0 (0.0%) | 0.04 |
| Hypertension (n, %) | 2 (40.0%) | 5 (37.7%) | 1 (9.1%) | 0.30 |
| Dyslipidemia (n, %) | 1 (20.0%) | 6 (20.0%) | 4 (42.9%) | 0.88 |
| HOMA-IR p50 (IQR) | 2.9 (1.88) | 1.8 (3.03) | 1.7 (2.57) | 0.93 |
| HOMA-B p50 (IQR) | 132.1 (114.9) | 110.2 (155.8) | 135.1 (100.2) | 0.60 |
| GH (ng/dL) p50 (IQR) | 5.0 (7.9) | 0.8 (2.2) a | 0.4 (0.5) a | 0.00 |
| IGF-1 (ng/dL) p50 (IQR) | 392.0 (121) | 216.5 (100) b | 179.0 (115) a | 0.00 |
| Lysine | Lactate | Valine | Isoleucine | Dimethylamine | |
|---|---|---|---|---|---|
| ACRO p50 (IQR) | 0.72 (0.05) | 12.26 (2.78) | 4.75 (0.87) | 2.54 (0.41) | 1.35 (0.18) |
| Control p50 (IQR) | 0.78 (0.08) | 14.85 (5.20) | 5.20 (1.06) | 2.80 (0.51) | 1.20 (0.18) |
| p-value Mann–Whitney | 0.002 | 0.001 | 0.019 | 0.023 | 0.016 |
| 1—Active (n = 5) | 2—ACRO_Medical (n = 14) | 3—ACRO_Remission (n = 11) | Jonckheere–Terpstra | ||
|---|---|---|---|---|---|
| Tau_b | p | ||||
| Isoleucine (AUs) | 2.50 (0.18) | 2.46 (0.44) | 2.67 (0.33) | 0.26 | 0.08 |
| Lysine (AUs) | 0.69 (0.02) | 0.71 (0.08) | 0.73 (0.03) | 0.28 | 0.06 |
| Valine (AUs) | 4.30 (0.24) | 4.71 (1.02) | 5.00 (0.54) | 0.30 | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biagetti, B.; Herance, J.R.; Ferrer, R.; Aulinas, A.; Palomino-Schätzlein, M.; Mesa, J.; Castaño, J.P.; Luque, R.M.; Simó, R. Metabolic Fingerprint of Acromegaly and Its Potential Usefulness in Clinical Practice. J. Clin. Med. 2019, 8, 1549. https://doi.org/10.3390/jcm8101549
Biagetti B, Herance JR, Ferrer R, Aulinas A, Palomino-Schätzlein M, Mesa J, Castaño JP, Luque RM, Simó R. Metabolic Fingerprint of Acromegaly and Its Potential Usefulness in Clinical Practice. Journal of Clinical Medicine. 2019; 8(10):1549. https://doi.org/10.3390/jcm8101549
Chicago/Turabian StyleBiagetti, Betina, J.R. Herance, Roser Ferrer, Anna Aulinas, Martina Palomino-Schätzlein, Jordi Mesa, J.P. Castaño, Raul M. Luque, and Rafael Simó. 2019. "Metabolic Fingerprint of Acromegaly and Its Potential Usefulness in Clinical Practice" Journal of Clinical Medicine 8, no. 10: 1549. https://doi.org/10.3390/jcm8101549