Stimulation of Human Osteoblast Differentiation in Magneto-Mechanically Actuated Ferromagnetic Fiber Networks

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Substrates—Fibre Networks
2.2. Cell Culture and Seeding
2.3. Cell Adhesion and Cytoskeleton Organization
2.4. Cell Mineralization
2.4.1. OsteoImageTM Mineralization Assay
2.4.2. Alizarin Red Staining
2.5. Gene Expression Analysis
2.6. Protein Release
2.7. Magneto-Mechanical Actuation
2.7.1. Magneto-Mechanical Actuation of Cell-Seeded Fiber Networks
2.7.2. Magneto-Mechanical Network Deflection
2.8. Statistical Analysis
3. Results
3.1. Fibre Network Deformation
3.2. Cell Adhesion and Cytoskeleton Organization
3.3. Cell Mineralization
3.4. Gene Expression Analysis
3.5. Protein Release
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3, S131–S139. [Google Scholar] [CrossRef]
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: Current concepts and future directions. BMC Med. 2011, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Lanyon, L.E.; Goodship, A.E.; Pye, C.J.; MacFie, J.H. Mechanically adaptive bone remodelling. J. Biomech. 1982, 15, 141–154. [Google Scholar] [CrossRef]
- Rubin, C.T.; Lanyon, L.E. Regulation of bone mass by mechanical strain magnitude. Calcif. Tissue Int. 1985, 37, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Bone “mass” and the “mechanostat”: A proposal. Anat. Rec. 1987, 219, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.D.; Coon, B.G.; Yun, S.; Baeyens, N.; Tanaka, K.; Ouyang, M.; Schwartz, M.A. Integrins in mechanotransduction. Curr. Opin. Cell Biol. 2013, 25, 613–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, A.W.; Helmke, B.P.; Blackman, B.R.; Schwartz, M.A. Mechanisms of mechanotransduction. Dev. Cell 2006, 10, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Zaidi, M. Molecular regulation of mechanotransduction. Biochem. Biophys. Res. Commun. 2005, 328, 751–755. [Google Scholar] [CrossRef]
- Markaki, A.E.; Clyne, T.W. Magneto-mechanical actuation of bonded ferromagnetic fibre arrays. Acta Mater. 2005, 53, 877–889. [Google Scholar] [CrossRef]
- Markaki, A.E.; Clyne, T.W. Magneto-mechanical stimulation of bone growth in a bonded array of ferromagnetic fibres. Biomaterials 2004, 25, 4805–4815. [Google Scholar] [CrossRef]
- Daish, C.; Blanchard, R.; Fox, K.; Pivonka, P.; Pirogova, E. The Application of Pulsed Electromagnetic Fields (PEMFs) for Bone Fracture Repair: Past and Perspective Findings. Ann. Biomed. Eng. 2018, 46, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Kotani, H.; Kawaguchi, H.; Shimoaka, T.; Iwasaka, M.; Ueno, S.; Ozawa, H.; Nakamura, K.; Hoshi, K. Strong static magnetic field stimulates bone formation to a definite orientation in vitro and in vivo. J. Bone Miner. Res. 2002, 17, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.C.; Tomita, N.; Ikada, Y. Effects of static magnetic field on bone formation of rat femurs. Med. Eng. Phys. 1998, 20, 397–402. [Google Scholar] [CrossRef]
- Pankhurst, Q.; Thanh, N.; Jones, S.; Dobson, J. Progress in applications of magnetic nanoparticles in biomedicine. J. Phys. D Appl. Phys. 2009, 42, 224001. [Google Scholar] [CrossRef]
- Dobson, J. Remote control of cellular behaviour with magnetic nanoparticles. Nat. Nanotechnol. 2008, 3, 139. [Google Scholar] [CrossRef]
- Yuge, L.; Okubo, A.; Miyashita, T.; Kumagai, T.; Nikawa, T.; Takeda, S.; Kanno, M.; Urabe, Y.; Sugiyama, M.; Kataoka, K. Physical stress by magnetic force accelerates differentiation of human osteoblasts. Biochem. Biophys. Res. Commun. 2003, 311, 32–38. [Google Scholar] [CrossRef]
- Yuan, Z.; Memarzadeh, K.; Stephen, A.S.; Allaker, R.P.; Brown, R.A.; Huang, J. Development of a 3D Collagen Model for the In Vitro Evaluation of Magnetic-assisted Osteogenesis. Sci. Rep. 2018, 8, 16270. [Google Scholar] [CrossRef]
- Henstock, J.R.; Rotherham, M.; El Haj, A.J. Magnetic ion channel activation of TREK1 in human mesenchymal stem cells using nanoparticles promotes osteogenesis in surrounding cells. J. Tissue Eng. 2018, 9, 2041731418808695. [Google Scholar] [CrossRef]
- Rotherham, M.; Nahar, T.; Goodman, T.; Telling, N.; Gates, M.; El Haj, A. Magnetic Mechanoactivation of Wnt Signaling Augments Dopaminergic Differentiation of Neuronal Cells. Adv. Biosyst. 2019, 3, 1900091. [Google Scholar] [CrossRef]
- Markaki, A.; Justin, A. A magneto-active scaffold for stimulation of bone growth. Mater. Sci. Technol. 2014, 30, 1590–1598. [Google Scholar] [CrossRef]
- Dowling, N.; Kim, Y.-H.; Ahn, S.-K.; Lee, Y.-D. Effect of alloying elements and residuals on corrosion resistance of type 444 stainless steel. Corrosion 1999, 55, 187–199. [Google Scholar] [CrossRef]
- Asami, K.; Hashimoto, K. Importance of initial surface film in the degradation of stainless steels by atmospheric exposure. Corros. Sci. 2003, 45, 2263–2283. [Google Scholar] [CrossRef]
- Malheiro, V.N.; Spear, R.L.; Brooks, R.A.; Markaki, A.E. Osteoblast and monocyte responses to 444 ferritic stainless steel intended for a magneto-mechanically actuated fibrous scaffold. Biomaterials 2011, 32, 6883–6892. [Google Scholar] [CrossRef]
- Bhaskar, B.; Owen, R.; Bahmaee, H.; Rao, P.S.; Reilly, G.C. Design and assessment of a dynamic perfusion bioreactor for large bone tissue engineering scaffolds. Appl. Biochem. Biotechnol. 2018, 185, 555–563. [Google Scholar] [CrossRef]
- Ravichandran, A.; Wen, F.; Lim, J.; Chong, M.S.K.; Chan, J.K.; Teoh, S.H. Biomimetic fetal rotation bioreactor for engineering bone tissues—Effect of cyclic strains on upregulation of osteogenic gene expression. J. Tissue Eng. Regen. Med. 2018, 12, e2039–e2050. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, H.; Cui, Y.; Li, R.; Meng, F.; Ye, Z.; Zhang, X. Calcium Channel Opening Rather than the Release of ATP Causes the Apoptosis of Osteoblasts Induced by Overloaded Mechanical Stimulation. Cell. Physiol. Biochem. 2017, 42, 441–454. [Google Scholar] [CrossRef]
- Beşkardes, I.G.; Hayden, R.S.; Glettig, D.L.; Kaplan, D.L.; Gümüşderelioğlu, M. Bone tissue engineering with scaffold-supported perfusion co-cultures of human stem cell-derived osteoblasts and cell line-derived osteoclasts. Process Biochem. 2017, 59, 303–311. [Google Scholar] [CrossRef]
- Mitra, D.; Whitehead, J.; Yasui, O.W.; Leach, J.K. Bioreactor culture duration of engineered constructs influences bone formation by mesenchymal stem cells. Biomaterials 2017, 146, 29–39. [Google Scholar] [CrossRef]
- Tang, X.; Teng, S.; Liu, C.; Jagodzinski, M. Influence of hydrodynamic pressure on the proliferation and osteogenic differentiation of bone mesenchymal stromal cells seeded on polyurethane scaffolds. J. Biomed. Mater. Res. Part A 2017, 105, 3445–3455. [Google Scholar] [CrossRef]
- Filipowska, J.; Reilly, G.C.; Osyczka, A.M. A single short session of media perfusion induces osteogenesis in hBMSCs cultured in porous scaffolds, dependent on cell differentiation stage. Biotechnol. Bioeng. 2016, 113, 1814–1824. [Google Scholar] [CrossRef]
- Teng, S.; Liu, C.; Guenther, D.; Omar, M.; Neunaber, C.; Krettek, C.; Jagodzinski, M. Influence of biomechanical and biochemical stimulation on the proliferation and differentiation of bone marrow stromal cells seeded on polyurethane scaffolds. Exp. Ther. Med. 2016, 11, 2086–2094. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Anada, T.; Masuda, T.; Takano-Yamamoto, T.; Suzuki, O. Effect of mechanical stress on differentiation of mouse mesenchymal stem cells seeded into an octacalcium phosphate–gelatin scaffold. Sens. Actuators B Chem. 2015, 220, 125–130. [Google Scholar] [CrossRef]
- Tanaka, S.M.; Tachibana, K. Frequency-dependence of mechanically stimulated osteoblastic calcification in tissue-engineered bone in vitro. Ann. Biomed. Eng. 2015, 43, 2083–2089. [Google Scholar] [CrossRef]
- Damaraju, S.; Matyas, J.R.; Rancourt, D.E.; Duncan, N.A. The role of gap junctions and mechanical loading on mineral formation in a collagen-I scaffold seeded with osteoprogenitor cells. Tissue Eng. Part A 2015, 21, 1720–1732. [Google Scholar] [CrossRef]
- Subramony, S.D.; Su, A.; Yeager, K.; Lu, H.H. Combined effects of chemical priming and mechanical stimulation on mesenchymal stem cell differentiation on nanofiber scaffolds. J. Biomech. 2014, 47, 2189–2196. [Google Scholar] [CrossRef]
- Bartnikowski, M.; Klein, T.J.; Melchels, F.P.; Woodruff, M.A. Effects of scaffold architecture on mechanical characteristics and osteoblast response to static and perfusion bioreactor cultures. Biotechnol. Bioeng. 2014, 111, 1440–1451. [Google Scholar] [CrossRef]
- Jia, Y.-Y.; Li, F.; Geng, N.; Gong, P.; Huang, S.-J.; Meng, L.-X.; Lan, J.; Ban, Y. Fluid flow modulates the expression of genes involved in the Wnt signaling pathway in osteoblasts in 3D culture conditions. Int. J. Mol. Med. 2014, 33, 1282–1288. [Google Scholar] [CrossRef]
- Henstock, J.R.; Rotherham, M.; Rashidi, H.; Shakesheff, K.M.; El Haj, A.J. Remotely activated mechanotransduction via magnetic nanoparticles promotes mineralization synergistically with bone morphogenetic protein 2: Applications for injectable cell therapy. Stem Cells Transl. Med. 2014, 3, 1363–1374. [Google Scholar] [CrossRef]
- Damaraju, S.; Matyas, J.R.; Rancourt, D.E.; Duncan, N.A. The effect of mechanical stimulation on mineralization in differentiating osteoblasts in collagen-I scaffolds. Tissue Eng. Part A 2014, 20, 3142–3153. [Google Scholar] [CrossRef]
- Malheiro, V.N.; Skepper, J.N.; Brooks, R.A.; Markaki, A.E. In vitro osteoblast response to ferritic stainless steel fiber networks for magneto-active layers on implants. J. Biomed. Mater. Res. Part A 2013, 101, 1588–1598. [Google Scholar] [CrossRef]
- Spear, R.L.; Brooks, R.A.; Markaki, A.E. Short-term in vitro responses of human peripheral blood monocytes to ferritic stainless steel fiber networks. J. Biomed. Mater. Res. A 2013, 101, 1456–1463. [Google Scholar] [CrossRef]
- Neelakantan, S.; Bosbach, W.; Woodhouse, J.; Markaki, A.E. Characterization and deformation response of orthotropic fibre networks with auxetic out-of-plane behaviour. Acta Mater. 2014, 66, 326–339. [Google Scholar] [CrossRef]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and β-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast differentiation and bone matrix formation in vivo and in vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef]
- Pettersson, L.F.; Kingham, P.J.; Wiberg, M.; Kelk, P. In vitro osteogenic differentiation of human mesenchymal stem cells from jawbone compared with dental tissue. Tissue Eng. Regen. Med. 2017, 14, 763–774. [Google Scholar] [CrossRef]
- Hiemer, B.; Ziebart, J.; Jonitz-Heincke, A.; Grunert, P.C.; Su, Y.; Hansmann, D.; Bader, R. Magnetically induced electrostimulation of human osteoblasts results in enhanced cell viability and osteogenic differentiation. Int. J. Mol. Med. 2016, 38, 57–64. [Google Scholar] [CrossRef]
- Chen, W.; Liu, X.; Chen, Q.; Bao, C.; Zhao, L.; Zhu, Z.; Xu, H.H. Angiogenic and osteogenic regeneration in rats via calcium phosphate scaffold and endothelial cell co-culture with human bone marrow mesenchymal stem cells (MSCs), human umbilical cord MSCs, human induced pluripotent stem cell-derived MSCs and human embryonic stem cell-derived MSCs. J. Tissue Eng. Regen. Med. 2018, 12, 191–203. [Google Scholar]
- Rutkovskiy, A.; Stensløkken, K.-O.; Vaage, I.J. Osteoblast differentiation at a glance. Med. Sci. Monit. Basic Res. 2016, 22, 95. [Google Scholar] [CrossRef]
- Dumas, V.; Perrier, A.; Malaval, L.; Laroche, N.; Guignandon, A.; Vico, L.; Rattner, A. The effect of dual frequency cyclic compression on matrix deposition by osteoblast-like cells grown in 3D scaffolds and on modulation of VEGF variant expression. Biomaterials 2009, 30, 3279–3288. [Google Scholar] [CrossRef]
- Sittichockechaiwut, A.; Scutt, A.M.; Ryan, A.J.; Bonewald, L.F.; Reilly, G.C. Use of rapidly mineralising osteoblasts and short periods of mechanical loading to accelerate matrix maturation in 3D scaffolds. Bone 2009, 44, 822–829. [Google Scholar] [CrossRef]
- Zong ming, W.; Jian yu, L.; Rui xin, L.; Hao, L.; Yong, G.; Lu, L.; Xin chang, Z.; Xi zheng, Z. Bone formation in rabbit cancellous bone explant culture model is enhanced by mechanical load. Biomed. Eng. Online 2013, 12, 35. [Google Scholar] [CrossRef]
- Matsubara, T.; Kida, K.; Yamaguchi, A.; Hata, K.; Ichida, F.; Meguro, H.; Aburatani, H.; Nishimura, R.; Yoneda, T. BMP2 regulates Osterix through Msx2 and Runx2 during osteoblast differentiation. J. Biol. Chem. 2008, 283, 29119–29125. [Google Scholar] [CrossRef]
- Lou, Y.; Javed, A.; Hussain, S.; Colby, J.; Frederick, D.; Pratap, J.; Xie, R.; Gaur, T.; Van Wijnen, A.J.; Jones, S.N. A Runx2 threshold for the cleidocranial dysplasia phenotype. Hum. Mol. Genet. 2008, 18, 556–568. [Google Scholar] [CrossRef] [Green Version]
- Ignatius, A.; Blessing, H.; Liedert, A.; Schmidt, C.; Neidlinger-Wilke, C.; Kaspar, D.; Friemert, B.; Claes, L. Tissue engineering of bone: Effects of mechanical strain on osteoblastic cells in type I collagen matrices. Biomaterials 2005, 26, 311–318. [Google Scholar] [CrossRef]
- Rath, B.; Nam, J.; Knobloch, T.J.; Lannutti, J.J.; Agarwal, S. Compressive forces induce osteogenic gene expression in calvarial osteoblasts. J. Biomech. 2008, 41, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, Y.; Suzawa, M.; Kikuchi, T.; Matsumoto, T. Interaction of matrix collagen with osteoblastic cells enhances stimulatory effects of bone morphogenetic protein (BMP)-2 on the differentiation of osteoblasts. Bone 1995, 6, 568. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Z. Influence of mesenchymal stem cells with endothelial progenitor cells in co-culture on osteogenesis and angiogenesis: An in vitro study. Arch. Med. Res. 2013, 44, 504–513. [Google Scholar] [CrossRef]
- Samee, M.; Kasugai, S.; Kondo, H.; Ohya, K.; Shimokawa, H.; Kuroda, S. Bone morphogenetic protein-2 (BMP-2) and vascular endothelial growth factor (VEGF) transfection to human periosteal cells enhances osteoblast differentiation and bone formation. J. Pharmacol. Sci. 2008, 18, 0809050139. [Google Scholar] [CrossRef]
- Rumpler, M.; Woesz, A.; Dunlop, J.W.; van Dongen, J.T.; Fratzl, P. The effect of geometry on three-dimensional tissue growth. J. R. Soc. Interface 2008, 5, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammoto, T.; Ingber, D.E. Mechanical control of tissue and organ development. Development 2010, 137, 1407–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]








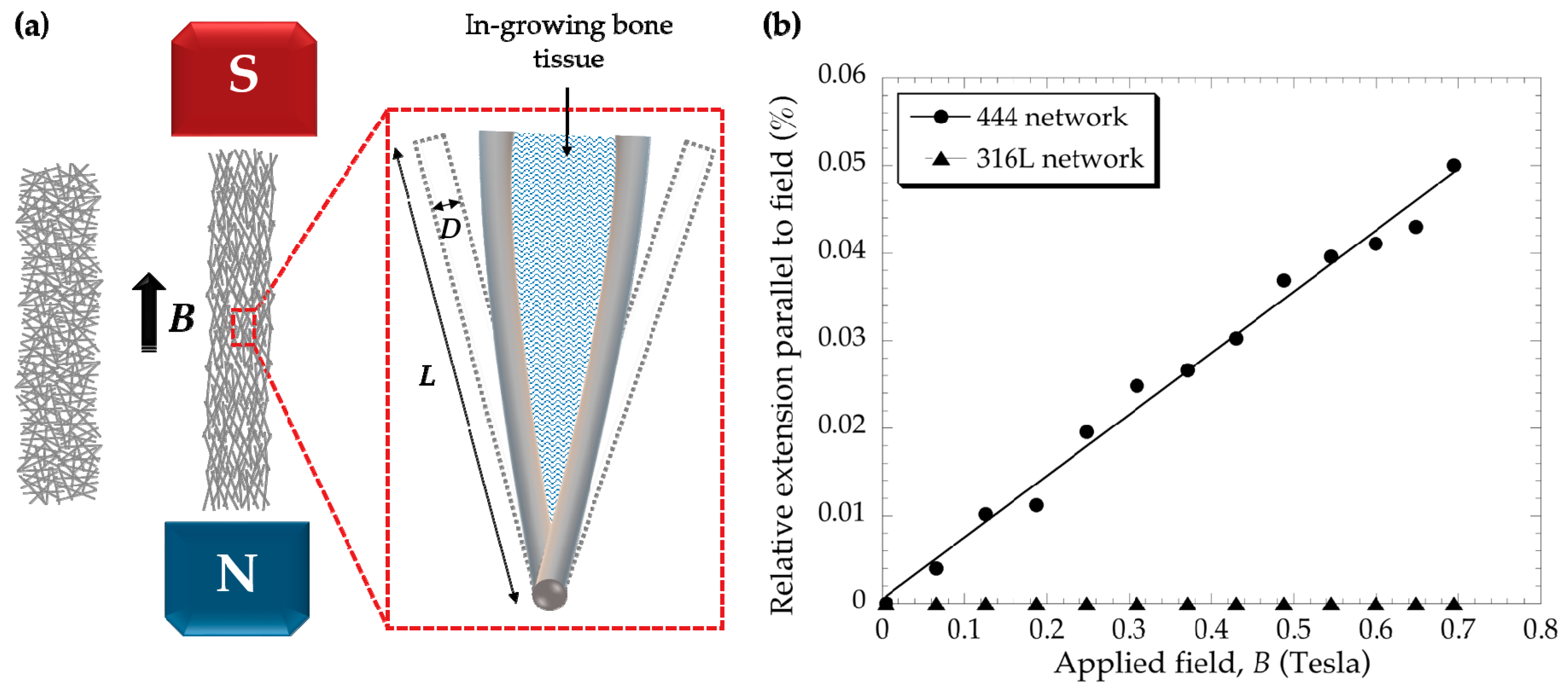{kind=link}
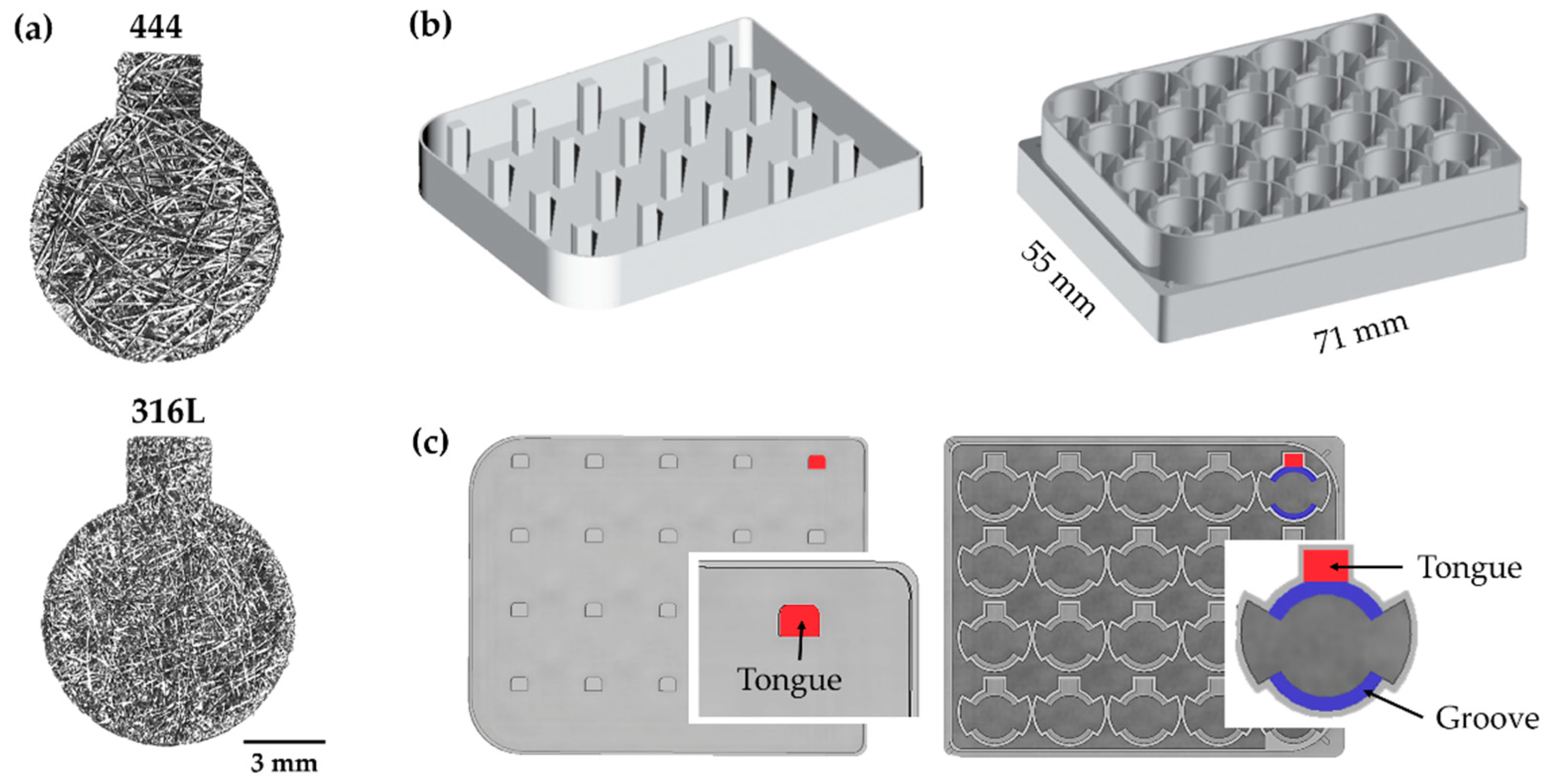{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scaffold | Cell Type | Stimulation Device | Mechanical Stimulation | Effect of Mechanical Stimulation on Bone-Related Transcriptions, Growth Factors, and Proteins | Ref |
|---|---|---|---|---|---|
| PU foam | hES-MP | Perfusion bioreactor | Flow rate: 3.47 mL·min−1 | ALP and DNA significantly increased. | [24] |
| PCL-TCP | hBMSC | Perfusion bioreactor | Cyclic compression: 0.22%, 1 Hz, 4 h per day. Biaxial rotation: 5 rpm, 90° angle, flow rate of 3.8 mL·min−1. | Combination of cyclic compression and biaxial rotation showed upregulation of ALP by 3.2–fold, OPN by 2.4–fold, OCN by 10–fold, COL1A1– by 2–fold, Runx2 by 3.5–fold | [25] |
| PCL | MC3T3-E1 | Loading device | Compression at 10 millistrain, 1 Hz, 0.5 h and 2 h for 4/8/12 days | ATP release was induced by overloading and further increased the intracellular calcium concentration | [26] |
| Chitosan-HA super-porous hydrogel | hBMSC & THP-1 | Perfusion bioreactor | Bidirectional perfusion, flow rate of 6 mL·h−1, 28 days | ALP, BSP, COL1A1, ON, TRAP, and IL-6 upregulated, Runx2, OPN, and OCN not affected | [27] |
| HA-PLG | hBMSC | Perfusion bioreactor and XYZ shaker | Continuous perfusion at 3 mL·min−1 or XYZ shaker, 30 rpm, max tilt angle of 4.5° | SP7 and COL1A1 upregulated after 7 days in the continuous perfusion. No differences at 14 and 21 days. Ca deposited increased in the continuous perfusion at 7, 14, and 21 days compared to the shaker. | [28] |
| PU-based meniscus | rBMSC | Perfusion bioreactor | Flow rate of 10 mL·min−1 and hydrodynamic pressure (2 s per cycle) at 10 mL·min−1, 0.5 Hz, at hydraulic pressure: 0–60 or 0–120 mmHg. | ALP upregulated by 4, 6.5, 6.2-fold and OC upregulated by 1, 5.5, 5.7-fold for 0–60 mmHg group after 1,2,3 weeks, respectively. No noticeable effect for 0–120 mmHg group. | [29] |
| Gelatin-coated porous polyurethane | hBMPC | Perfusion bioreactor | Single perfusion session for 2 h, flow rate of 2.5 mL·min−1 on day 5 or 7. | ALP and Runx2 activity increased on day 7 compared to day 5 and static controls. BMP-2 increased on day 5 compared to day 7 and static controls. | [30] |
| PU-based | hBMSC | Perfusion bioreactor | Perfusion at 10 mL·min−1 with 10% cyclic compression 0.5 or 5 Hz. | Runx2 inhibited in 5 Hz group on day 7. COL1A1 upregulated on day 7 and OCN enhanced on day 14 for 0.5 Hz. | [31] |
| Octacalcium phosphate and gelatin | mBMSC | Loading device | Cyclic compressive strains of 20, 40, and 60%, 0.75 Hz, 4 h for 3 or 7 days. | ALP activity decreased with increasing strains. COL1 was not significantly affected. OPN and OCN upregulated for the 20% cyclic strain group compared to the control and other strain groups. | [32] |
| Collagen I | rBMSC | A piezo-type mechanical stimulator | Sinusoidal compressive deformation, 0.2% at 0.2, 2, 10, 20, 40, 60 Hz, 3 min per day. | Ca deposition (optically monitored) increased on days 5–7 and reached the highest value for 2 Hz (1.5-fold) on day 14. | [33] |
| Collagen I | mESC | Loading device (six-well loading plate) | 5% cyclic compressive strain, 1 Hz, 2 loading cycles, 4 h followed by 16-h rest (total: 40 h). | Runx2 upregulated in loaded constructs on days 15, 20, and 30. | [34] |
| PLGA | hMSC | Bioreactor | Dynamic tensile stimulation: 1% strain at 1 Hz for 90 min twice daily for 28 days. | COL1, COL2, fibronectin and Tenascin-C were upregulated. | [35] |
| PCL | MC3T3-E1 | Perfusion bioreactor | Computational fluid dynamic (CFD) analysis: flow rate of 1 mL·h−1, wall shear stress 3 Pa. | ALP per DNA values in perfusion culture increased from 14 to 28 days. | [36] |
| Partially deproteinized bone | Osteoblast-like rat osteosarcoma cells (ROS17/2.8) | 3D fluid flow cell culture system | Shear stress of 0.8 Pa, loading for 1 h/2 h/4 h followed by statically incubated for 23 h/22 h/20 h. | ALP, LRP5, β-catenin and Wnt3A, upregulated in all loading conditions. | [37] |
| Collagen hydrogel | hMSCs | Magnetic Force Bioreactor | Magnetic field of 25 mT and 1 Hz for 1 h a day for 28 days | 2.4-fold increase in mineralization and matrix density | [38] |
| Collagen I | mESCs | Loading device (six-well loading plate) | 5% cyclic compressive strain, 1 Hz, 2 loading cycles, 4 h followed by 16-h rest (total: 40 h). | Connexin-43 was 12-fold on day 5 and 5-fold on day 30. Oct-4 increased on day 5. Runx2 increased on day 7. | [39] |
| Fiber Network | Fiber Volume Fraction (%) | Fiber Cross-Sectional Shape | Mean Fiber Inclination Angle to the Vertical (°) |
|---|---|---|---|
| 444 | 15.7 ± 1.0 | Rectangular (60 × 100 µm2) | 81.87 ± 0.21 |
| 316L | 15.4 ± 0.9 | Hexagonal (side length 20 µm, diagonal length 40 µm) | 83.49 ± 0.09 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katarivas Levy, G.; Birch, M.A.; Brooks, R.A.; Neelakantan, S.; Markaki, A.E. Stimulation of Human Osteoblast Differentiation in Magneto-Mechanically Actuated Ferromagnetic Fiber Networks. J. Clin. Med. 2019, 8, 1522. https://doi.org/10.3390/jcm8101522
Katarivas Levy G, Birch MA, Brooks RA, Neelakantan S, Markaki AE. Stimulation of Human Osteoblast Differentiation in Magneto-Mechanically Actuated Ferromagnetic Fiber Networks. Journal of Clinical Medicine. 2019; 8(10):1522. https://doi.org/10.3390/jcm8101522
Chicago/Turabian StyleKatarivas Levy, Galit, Mark A. Birch, Roger A. Brooks, Suresh Neelakantan, and Athina E. Markaki. 2019. "Stimulation of Human Osteoblast Differentiation in Magneto-Mechanically Actuated Ferromagnetic Fiber Networks" Journal of Clinical Medicine 8, no. 10: 1522. https://doi.org/10.3390/jcm8101522