
Membrane Heteroreceptor Complexes as Second-Order Protein Modulators: A Novel Integrative Mechanism through Allosteric Receptor–Receptor Interactions
 , ,
, ,  ,
,  , , and
, , and 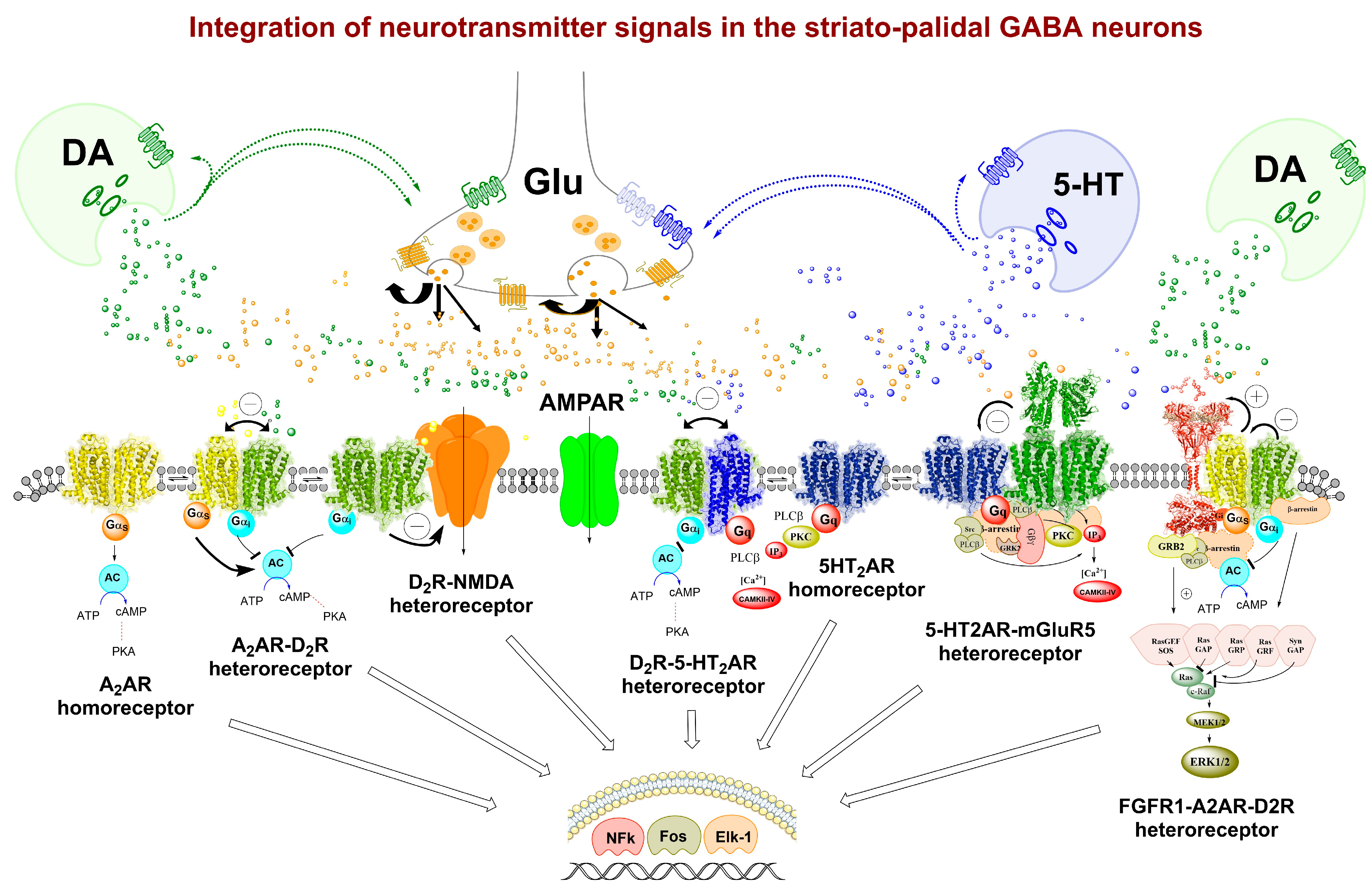{kind=link}
Abstract
:1. Introduction
2. On the Existence of GPCR Homo- and Heteroreceptor Complexes
2.1. GPCR-GPCR Heterocomplexes
2.2. GPCR-Ion Channels Heterocomplexes
2.3. GPCR–RTK Heterocomplexes
3. Molecular Integrations through Allosteric Receptor–Receptor Interactions in Heteroreceptor Complexes of Synaptic and Extra-Synaptic Regions
3.1. From Modulation of Receptor Protomer Recognition, Signaling and Trafficking to Functions in the CNS, including Behavioral Studies
3.2. Expanding the Concept of Meta-Modulation (Second-Order Modulation) and Protein Modulation Based on the Existence of Allosteric Receptor–Receptor Interactions in Heteroreceptor Complexes

4. Learning, Memory, and Synchrony at the Molecular Level
5. Allosteric Receptor–Receptor Interactions and Their Involvement in Brain Disease and Its Treatment
5.1. Pathophysiological Relevance for Major Depressive Disorder and Potential Drug Development
5.2. Pathophysiological Relevance for Cocaine Use Disorder and Potential Drug Development
5.3. Pathophysiological Relevance for Parkinson’s Disease and Drug Development
6. Conclusions and Future Aspects
Author Contributions
Funding
Conflicts of Interest
References
- Fuxe, K.; Agnati, L.F.; Benfenati, F.; Celani, M.; Zini, I.; Zoli, M.; Mutt, V. Evidence for the Existence of Receptor Receptor Interactions in the Central Nervous-System—Studies on the Regulation of Monoamine Receptors by Neuropeptides. J. Neural Transm. 1983, 18, 165–179. [Google Scholar]
- Zoli, M.; Agnati, L.F.; Hedlund, P.B.; Li, X.M.; Ferre, S.; Fuxe, K. Receptor-Receptor Interactions an Integrative Mechanism in Nerve-Cells. Mol. Neurobiol. 1993, 7, 293–334. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Agnati, L.F.; Borroto-Escuela, D.O. The impact of receptor-receptor interactions in heteroreceptor complexes on brain plasticity. Expert Rev. Neurother. 2014, 14, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Carlsson, J.; Ambrogini, P.; Narvaez, M.; Wydra, K.; Tarakanov, A.O.; Li, X.; Millon, C.; Ferraro, L.; Cuppini, R.; et al. Understanding the Role of GPCR Heteroreceptor Complexes in Modulating the Brain Networks in Health and Disease. Front. Cell Neurosci. 2017, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Frankowska, M.; Ferraro, L.; Guidolin, D.; Ciruela, F.; Agnati, L.F. The changing world of G protein-coupled receptors: From monomers to dimers and receptor mosaics with allosteric receptor-receptor interactions. J. Recept. Signal Transduct. Res. 2010, 30, 272–283. [Google Scholar] [CrossRef] [PubMed]
- White, J.H.; Wise, A.; Main, M.J.; Green, A.; Fraser, N.J.; Disney, G.H.; Barnes, A.A.; Emson, P.; Foord, S.M.; Marshall, F.H. Heterodimerization is required for the formation of a functional GABA(B) receptor. Nature 1998, 396, 679–682. [Google Scholar] [CrossRef]
- Marshall, F.H.; White, J.; Main, M.; Green, A.; Wise, A. GABA(B) receptors function as heterodimers. Biochem. Soc. Trans. 1999, 27, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.; Meyerhof, W. Oligomerization of sweet and bitter taste receptors. Methods Cell Biol. 2013, 117, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.S.; Park, E.A.; Sallee, F.R.; Parker, S.L. Two intracellular helices of G-protein coupling receptors could generally support oligomerization and coupling with transducers. Amino Acids 2011, 40, 261–268. [Google Scholar] [CrossRef]
- Lee, F.J.; Xue, S.; Pei, L.; Vukusic, B.; Chery, N.; Wang, Y.; Wang, Y.T.; Niznik, H.B.; Yu, X.M.; Liu, F. Dual regulation of NMDA receptor functions by direct protein-protein interactions with the dopamine D1 receptor. Cell 2002, 111, 219–230. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Tarakanov, A.O.; Fuxe, K. FGFR1-5-HT1A Heteroreceptor Complexes: Implications for Understanding and Treating Major Depression. Trends Neurosci. 2016, 39, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Kotliar, I.B.; Lorenzen, E.; Schwenk, J.M.; Hay, D.L.; Sakmar, T.P. Elucidating the Interactome of G Protein-Coupled Receptors and Receptor Activity-Modifying Proteins. Pharmacol. Rev. 2023, 75, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Arapulisamy, O.; Mannangatti, P.; Jayanthi, L.D. Regulated norepinephrine transporter interaction with the neurokinin-1 receptor establishes transporter subcellular localization. J. Biol. Chem. 2013, 288, 28599–28610. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wan, Q.; Pristupa, Z.B.; Yu, X.M.; Wang, Y.T.; Niznik, H.B. Direct protein-protein coupling enables cross-talk between dopamine D5 and gamma-aminobutyric acid A receptors. Nature 2000, 403, 274–280. [Google Scholar] [CrossRef]
- Perez de la Mora, M.; Ferre, S.; Fuxe, K. GABA-dopamine receptor-receptor interactions in neostriatal membranes of the rat. Neurochem. Res. 1997, 22, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Flajolet, M.; Wang, Z.; Futter, M.; Shen, W.; Nuangchamnong, N.; Bendor, J.; Wallach, I.; Nairn, A.C.; Surmeier, D.J.; Greengard, P. FGF acts as a co-transmitter through adenosine A(2A) receptor to regulate synaptic plasticity. Nat. Neurosci. 2008, 11, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Mudo, G.; Perez-Alea, M.; Ciruela, F.; Tarakanov, A.O.; Narvaez, M.; Di Liberto, V.; Agnati, L.F.; Belluardo, N.; et al. Fibroblast Growth Factor Receptor 1- 5-Hydroxytryptamine 1A Heteroreceptor Complexes and Their Enhancement of Hippocampal Plasticity. Biol. Psychiatry 2012, 71, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Borroto-Escuela, D.O. Heteroreceptor Complexes and their Allosteric Receptor-Receptor Interactions as a Novel Biological Principle for Integration of Communication in the CNS: Targets for Drug Development. Neuropsychopharmacology 2016, 41, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Borroto-Escuela, D.O.; Romero-Fernandez, W.; Palkovits, M.; Tarakanov, A.O.; Ciruela, F.; Agnati, L.F. Moonlighting proteins and protein-protein interactions as neurotherapeutic targets in the G protein-coupled receptor field. Neuropsychopharmacology 2014, 39, 131–155. [Google Scholar] [CrossRef]
- Kolb, P.; Kenakin, T.; Alexander, S.P.H.; Bermudez, M.; Bohn, L.M.; Breinholt, C.S.; Bouvier, M.; Hill, S.J.; Kostenis, E.; Martemyanov, K.A.; et al. Community guidelines for GPCR ligand bias: IUPHAR review 32. Br. J. Pharmacol. 2022, 179, 3651–3674. [Google Scholar] [CrossRef]
- Kenakin, T. Functional selectivity and biased receptor signaling. J. Pharmacol. Exp. Ther. 2011, 336, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Chini, B. Expanding neuropeptide signalling by multiplying receptor functional states and sub-cellular locations. Cell Tissue Res. 2019, 375, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suarez-Boomgaard, D.; Rivera, A.; et al. The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Perez de la Mora, M.; Borroto-Escuela, D.O.; Crespo-Ramirez, M.; Rejon-Orantes, J.D.C.; Palacios-Lagunas, D.A.; Martinez-Mata, M.K.; Sanchez-Luna, D.; Tesoro-Cruz, E.; Fuxe, K. Dysfunctional Heteroreceptor Complexes as Novel Targets for the Treatment of Major Depressive and Anxiety Disorders. Cells 2022, 11, 1826. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; DuPont, C.M.; Li, X.; Savelli, D.; Lattanzi, D.; Srivastava, I.; Narvaez, M.; Di Palma, M.; Barbieri, E.; Andrade-Talavera, Y.; et al. Disturbances in the FGFR1-5-HT1A Heteroreceptor Complexes in the Raphe-Hippocampal 5-HT System Develop in a Genetic Rat Model of Depression. Front. Cell. Neurosci. 2017, 11, 309. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. Seven transmembrane receptors: Something old, something new. Acta Physiol. 2007, 190, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, V.V.; Gurevich, E.V. GPCR monomers and oligomers: It takes all kinds. Trends Neurosci. 2008, 31, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Javitch, J.A. The ants go marching two by two: Oligomeric structure of G-protein-coupled receptors. Mol. Pharmacol. 2004, 66, 1077–1082. [Google Scholar] [CrossRef]
- Lee, S.P.; O’Dowd, B.F.; George, S.R. Homo- and hetero-oligomerization of G protein-coupled receptors. Life Sci. 2003, 74, 173–180. [Google Scholar] [CrossRef]
- Dale, N.C.; Johnstone, E.K.M.; Pfleger, K.D.G. GPCR heteromers: An overview of their classification, function and physiological relevance. Front. Endocrinol. 2022, 13, 931573. [Google Scholar] [CrossRef]
- Han, Y.; Moreira, I.S.; Urizar, E.; Weinstein, H.; Javitch, J.A. Allosteric communication between protomers of dopamine class A GPCR dimers modulates activation. Nat. Chem. Biol. 2009, 5, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Jordan, B.A.; Devi, L.A. G-protein-coupled receptor heterodimerization modulates receptor function. Nature 1999, 399, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Romero-Fernandez, W.; Borroto-Escuela, D.O.; Agnati, L.F.; Fuxe, K. Evidence for the existence of dopamine D2-oxytocin receptor heteromers in the ventral and dorsal striatum with facilitatory receptor-receptor interactions. Mol. Psychiatry 2013, 18, 849–850. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Li, X.; Tarakanov, A.O.; Savelli, D.; Narvaez, M.; Shumilov, K.; Andrade-Talavera, Y.; Jimenez-Beristain, A.; Pomierny, B.; Diaz-Cabiale, Z.; et al. Existence of Brain 5-HT1A-5-HT2A Isoreceptor Complexes with Antagonistic Allosteric Receptor-Receptor Interactions Regulating 5-HT1A Receptor Recognition. ACS Omega 2017, 2, 4779–4789. [Google Scholar] [CrossRef] [PubMed]
- Terrillon, S.; Durroux, T.; Mouillac, B.; Breit, A.; Ayoub, M.A.; Taulan, M.; Jockers, R.; Barberis, C.; Bouvier, M. Oxytocin and vasopressin V1a and V2 receptors form constitutive homo- and heterodimers during biosynthesis. Mol. Endocrinol. 2003, 17, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Kaupmann, K.; Malitschek, B.; Schuler, V.; Heid, J.; Froestl, W.; Beck, P.; Mosbacher, J.; Bischoff, S.; Kulik, A.; Shigemoto, R.; et al. GABA(B)-receptor subtypes assemble into functional heteromeric complexes. Nature 1998, 396, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Galvez, T.; Duthey, B.; Kniazeff, J.; Blahos, J.; Rovelli, G.; Bettler, B.; Prezeau, L.; Pin, J.P. Allosteric interactions between GB1 and GB2 subunits are required for optimal GABA(B) receptor function. EMBO J. 2001, 20, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Ramsay, D.; Pascal, G.; Carrillo, J.J. GPCR dimerisation. Life Sci. 2003, 74, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Muller, A.; Chou, Y.Y.; Ji, I.; Lajic, S.; Hanyaloglu, A.C.; Jonas, K.; Rahman, N.; Ji, T.H.; Huhtaniemi, I. Rescue of defective G protein-coupled receptor function in vivo by intermolecular cooperation. Proc. Natl. Acad. Sci. USA 2010, 107, 2319–2324. [Google Scholar] [CrossRef]
- Lee, C.; Ji, I.J.; Ji, T.H. Use of defined-function mutants to access receptor-receptor interactions. Methods 2002, 27, 318–323. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Flajolet, M.; Agnati, L.F.; Greengard, P.; Fuxe, K. Bioluminescence resonance energy transfer methods to study G protein-coupled receptor-receptor tyrosine kinase heteroreceptor complexes. Methods Cell Biol. 2013, 117, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, H.; De Francesco, P.N.; Kandil, D.; Theeuwes, W.F.; McCarthy, T.; van Oeffelen, W.E.; Perello, M.; Giblin, L.; Dinan, T.G.; Cryan, J.F. Ghrelin’s Orexigenic Effect Is Modulated via a Serotonin 2C Receptor Interaction. ACS Chem. Neurosci. 2015, 6, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gong, Z.; Lu, Y.B.; Xu, C.J.; Wei, T.F.; Yang, M.S.; Zhan, T.W.; Yang, Y.H.; Lin, L.; Liu, J.; et al. FLIM-FRET-Based Structural Characterization of a Class-A GPCR Dimer in the Cell Membrane. J. Mol. Biol. 2020, 432, 4596–4611. [Google Scholar] [CrossRef] [PubMed]
- Herrick-Davis, K.; Grinde, E.; Cowan, A.; Mazurkiewicz, J.E. Fluorescence correlation spectroscopy analysis of serotonin, adrenergic, muscarinic, and dopamine receptor dimerization: The oligomer number puzzle. Mol. Pharmacol. 2013, 84, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Herrick-Davis, K.; Grinde, E.; Lindsley, T.; Cowan, A.; Mazurkiewicz, J.E. Oligomer size of the serotonin 5-hydroxytryptamine 2C (5-HT2C) receptor revealed by fluorescence correlation spectroscopy with photon counting histogram analysis: Evidence for homodimers without monomers or tetramers. J. Biol. Chem. 2012, 287, 23604–23614. [Google Scholar] [CrossRef] [PubMed]
- Jonas, K.C.; Huhtaniemi, I.; Hanyaloglu, A.C. Single-molecule resolution of G protein-coupled receptor (GPCR) complexes. Methods Cell Biol. 2016, 132, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Jonas, K.C.; Fanelli, F.; Huhtaniemi, I.T.; Hanyaloglu, A.C. Single molecule analysis of functionally asymmetric G protein-coupled receptor (GPCR) oligomers reveals diverse spatial and structural assemblies. J. Biol. Chem. 2015, 290, 3875–3892. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Xu, J.; Liang, Y.; Fang, X. Single-Molecule Imaging Reveals Differential AT1R Stoichiometry Change in Biased Signaling. Int. J. Mol. Sci. 2023, 25, 374. [Google Scholar] [CrossRef] [PubMed]
- Pediani, J.D.; Ward, R.J.; Marsango, S.; Milligan, G. Spatial Intensity Distribution Analysis: Studies of G Protein-Coupled Receptor Oligomerisation. Trends Pharmacol. Sci. 2018, 39, 175–186. [Google Scholar] [CrossRef]
- Perreault, M.L.; Hasbi, A.; Shen, M.Y.F.; Fan, T.; Navarro, G.; Fletcher, P.J.; Franco, R.; Lanciego, J.L.; George, S.R. Disruption of a dopamine receptor complex amplifies the actions of cocaine. Eur. Neuropsychopharm. 2016, 26, 1366–1377. [Google Scholar] [CrossRef]
- Nai, Q.; Li, S.; Wang, S.H.; Liu, J.; Lee, F.J.; Frankland, P.W.; Liu, F. Uncoupling the D1-N-methyl-D-aspartate (NMDA) receptor complex promotes NMDA-dependent long-term potentiation and working memory. Biol. Psychiatry 2009, 67, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Cottet, M.; Faklaris, O.; Falco, A.; Trinquet, E.; Pin, J.P.; Mouillac, B.; Durroux, T. Fluorescent ligands to investigate GPCR binding properties and oligomerization. Biochem. Soc. Trans. 2013, 41, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Cottet, M.; Albizu, L.; Comps-Agrar, L.; Trinquet, E.; Pin, J.P.; Mouillac, B.; Durroux, T. Time resolved FRET strategy with fluorescent ligands to analyze receptor interactions in native tissues: Application to GPCR oligomerization. Methods Mol. Biol. 2011, 746, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Albizu, L.; Cottet, M.; Kralikova, M.; Stoev, S.; Seyer, R.; Brabet, I.; Roux, T.; Bazin, H.; Bourrier, E.; Lamarque, L.; et al. Time-resolved FRET between GPCR ligands reveals oligomers in native tissues. Nat. Chem. Biol. 2010, 6, 587–594. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Craenenbroeck, K.V.; Romero-Fernandez, W.; Guidolin, D.; Woods, A.S.; Rivera, A.; Haegeman, G.; Agnati, L.F.; Tarakanov, A.O.; Fuxe, K. Dopamine D2 and D4 receptor heteromerization and its allosteric receptor-receptor interactions. Biochem. Biophys. Res. Commun. 2010, 404, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Garriga, P.; Ciruela, F.; Narvaez, M.; Tarakanov, A.O.; Palkovits, M.; Agnati, L.F.; Fuxe, K. G protein-coupled receptor heterodimerization in the brain. Methods Enzymol. 2013, 521, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Trifilieff, P.; Rives, M.L.; Urizar, E.; Piskorowski, R.A.; Vishwasrao, H.D.; Castrillon, J.; Schmauss, C.; Slattman, M.; Gullberg, M.; Javitch, J.A. Detection of antigen interactions ex vivo by proximity ligation assay: Endogenous dopamine D2-adenosine A2A receptor complexes in the striatum. Biotechniques 2011, 51, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Soderberg, O.; Gullberg, M.; Jarvius, M.; Ridderstrale, K.; Leuchowius, K.J.; Jarvius, J.; Wester, K.; Hydbring, P.; Bahram, F.; Larsson, L.G.; et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nat. Methods 2006, 3, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Raykova, D.; Kermpatsou, D.; Malmqvist, T.; Harrison, P.J.; Sander, M.R.; Stiller, C.; Heldin, J.; Leino, M.; Ricardo, S.; Klemm, A.; et al. A method for Boolean analysis of protein interactions at a molecular level. Nat. Commun. 2022, 13, 4755. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.O. Receptor-Receptor Interactions in the Central Nervous System; Fuxe, K., Borroto-Escuela, D.O., Eds.; Humana Press: New York, NY, USA, 2018; Volume 140, p. 346. [Google Scholar]
- Romero-Fernandez, W.; Carvajal-Tapia, C.; Prusky, A.; Katdare, K.A.; Wang, E.; Shostak, A.; Ventura-Antunes, L.; Harmsen, H.J.; Lippmann, E.S.; Fuxe, K.; et al. Detection, visualization and quantification of protein complexes in human Alzheimer’s disease brains using proximity ligation assay. Sci. Rep. 2023, 13, 11948. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Lopez-Salas, A.; Wydra, K.; Bartolini, M.; Zhou, Z.; Frankowska, M.; Suder, A.; Benitez-Porres, J.; Romero-Fernandez, W.; Filip, M.; et al. Combined treatment with Sigma1R and A2AR agonists fails to inhibit cocaine self-administration despite causing strong antagonistic accumbal A2AR-D2R complex interactions: The potential role of astrocytes. Front. Mol. Neurosci. 2023, 16, 1106765. [Google Scholar] [CrossRef] [PubMed]
- Romero-Fernandez, W.; Taura, J.J.; Crans, R.A.J.; Lopez-Cano, M.; Fores-Pons, R.; Narvaez, M.; Carlsson, J.; Ciruela, F.; Fuxe, K.; Borroto-Escuela, D.O. The mGlu5 Receptor Protomer-Mediated Dopamine D2 Receptor Trans-Inhibition Is Dependent on the Adenosine A2A Receptor Protomer: Implications for Parkinson’s Disease. Mol. Neurobiol. 2022, 59, 5955–5969. [Google Scholar] [CrossRef] [PubMed]
- Chruscicka, B.; Cowan, C.S.M.; Wallace Fitzsimons, S.E.; Borroto-Escuela, D.O.; Druelle, C.M.; Stamou, P.; Bergmann, C.A.; Dinan, T.G.; Slattery, D.A.; Fuxe, K.; et al. Molecular, biochemical and behavioural evidence for a novel oxytocin receptor and serotonin 2C receptor heterocomplex. Neuropharmacology 2021, 183, 108394. [Google Scholar] [CrossRef] [PubMed]
- Chruscicka, B.; Wallace Fitzsimons, S.E.; Borroto-Escuela, D.O.; Druelle, C.; Stamou, P.; Nally, K.; Dinan, T.G.; Cryan, J.F.; Fuxe, K.; Schellekens, H. Attenuation of Oxytocin and Serotonin 2A Receptor Signaling through Novel Heteroreceptor Formation. ACS Chem. Neurosci. 2019, 10, 3225–3240. [Google Scholar] [CrossRef] [PubMed]
- Stocco, E.; Sfriso, M.M.; Borile, G.; Contran, M.; Barbon, S.; Romanato, F.; Macchi, V.; Guidolin, D.; De Caro, R.; Porzionato, A. Experimental Evidence of A(2A)-D(2) Receptor-Receptor Interactions in the Rat and Human Carotid Body. Front. Physiol. 2021, 12, 645723. [Google Scholar] [CrossRef]
- Andrianarivelo, A.; Saint-Jour, E.; Pousinha, P.; Fernandez, S.P.; Petitbon, A.; De Smedt-Peyrusse, V.; Heck, N.; Ortiz, V.; Allichon, M.C.; Kappes, V.; et al. Disrupting D1-NMDA or D2-NMDA receptor heteromerization prevents cocaine’s rewarding effects but preserves natural reward processing. Sci. Adv. 2021, 7, eabg5970. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dwork, A.J.; Trifilieff, P.; Javitch, J.A. Detection of G Protein-Coupled Receptor Complexes in Postmortem Human Brain by Proximity Ligation Assay. Curr. Protoc. Neurosci. 2020, 91, e86. [Google Scholar] [CrossRef] [PubMed]
- Biezonski, D.K.; Trifilieff, P.; Meszaros, J.; Javitch, J.A.; Kellendonk, C. Evidence for limited D1 and D2 receptor coexpression and colocalization within the dorsal striatum of the neonatal mouse. J. Comp. Neurol. 2015, 523, 1175–1189. [Google Scholar] [CrossRef]
- Martinez-Pinilla, E.; Rico, A.J.; Rivas-Santisteban, R.; Lillo, J.; Roda, E.; Navarro, G.; Lanciego, J.L.; Franco, R. Expression of GPR55 and either cannabinoid CB(1) or CB(2) heteroreceptor complexes in the caudate, putamen, and accumbens nuclei of control, parkinsonian, and dyskinetic non-human primates. Brain Struct. Funct. 2020, 225, 2153–2164. [Google Scholar] [CrossRef]
- Martinez-Pinilla, E.; Reyes-Resina, I.; Onatibia-Astibia, A.; Zamarbide, M.; Ricobaraza, A.; Navarro, G.; Moreno, E.; Dopeso-Reyes, I.G.; Sierra, S.; Rico, A.J.; et al. CB1 and GPR55 receptors are co-expressed and form heteromers in rat and monkey striatum. Exp. Neurol. 2014, 261, 44–52. [Google Scholar] [CrossRef]
- Walsh, S.M.; Mathiasen, S.; Christensen, S.M.; Fay, J.F.; King, C.; Provasi, D.; Borrero, E.; Rasmussen, S.G.F.; Fung, J.J.; Filizola, M.; et al. Single Proteoliposome High-Content Analysis Reveals Differences in the Homo-Oligomerization of GPCRs. Biophys. J. 2018, 115, 300–312. [Google Scholar] [CrossRef]
- Song, W.; Duncan, A.L.; Sansom, M.S.P. Modulation of adenosine A2a receptor oligomerization by receptor activation and PIP(2) interactions. Structure 2021, 29, 1312–1325.e3. [Google Scholar] [CrossRef]
- Kasai, R.S.; Ito, S.V.; Awane, R.M.; Fujiwara, T.K.; Kusumi, A. The Class-A GPCR Dopamine D2 Receptor Forms Transient Dimers Stabilized by Agonists: Detection by Single-Molecule Tracking. Cell Biochem. Biophys. 2018, 76, 29–37. [Google Scholar] [CrossRef]
- Kasai, R.S.; Kusumi, A. Single-molecule imaging revealed dynamic GPCR dimerization. Curr. Opin. Cell Biol. 2014, 27, 78–86. [Google Scholar] [CrossRef]
- Kasai, R.S.; Suzuki, K.G.; Prossnitz, E.R.; Koyama-Honda, I.; Nakada, C.; Fujiwara, T.K.; Kusumi, A. Full characterization of GPCR monomer-dimer dynamic equilibrium by single molecule imaging. J. Cell Biol. 2011, 192, 463–480. [Google Scholar] [CrossRef]
- Chakraborty, H.; Jafurulla, M.; Clayton, A.H.A.; Chattopadhyay, A. Exploring oligomeric state of the serotonin(1A) receptor utilizing photobleaching image correlation spectroscopy: Implications for receptor function. Faraday Discuss. 2018, 207, 409–421. [Google Scholar] [CrossRef]
- Teichmann, A.; Gibert, A.; Lampe, A.; Grzesik, P.; Rutz, C.; Furkert, J.; Schmoranzer, J.; Krause, G.; Wiesner, B.; Schulein, R. The specific monomer/dimer equilibrium of the corticotropin-releasing factor receptor type 1 is established in the endoplasmic reticulum. J. Biol. Chem. 2014, 289, 24250–24262. [Google Scholar] [CrossRef]
- Marsango, S.; Jenkins, L.; Pediani, J.D.; Bradley, S.J.; Ward, R.J.; Hesse, S.; Biener, G.; Stoneman, M.R.; Tobin, A.B.; Raicu, V.; et al. The M(1) muscarinic receptor is present in situ as a ligand-regulated mixture of monomers and oligomeric complexes. Proc. Natl. Acad. Sci. USA 2022, 119, e2201103119. [Google Scholar] [CrossRef]
- Pediani, J.D.; Ward, R.J.; Godin, A.G.; Marsango, S.; Milligan, G. Dynamic Regulation of Quaternary Organization of the M1 Muscarinic Receptor by Subtype-selective Antagonist Drugs. J. Biol. Chem. 2016, 291, 13132–13146. [Google Scholar] [CrossRef]
- Park, P.S.; Wells, J.W. Monomers and oligomers of the M2 muscarinic cholinergic receptor purified from Sf9 cells. Biochemistry 2003, 42, 12960–12971. [Google Scholar] [CrossRef]
- Petersen, J.; Wright, S.C.; Rodriguez, D.; Matricon, P.; Lahav, N.; Vromen, A.; Friedler, A.; Stromqvist, J.; Wennmalm, S.; Carlsson, J.; et al. Agonist-induced dimer dissociation as a macromolecular step in G protein-coupled receptor signaling. Nat. Commun. 2017, 8, 226. [Google Scholar] [CrossRef]
- Nguyen, K.D.Q.; Vigers, M.; Sefah, E.; Seppala, S.; Hoover, J.P.; Schonenbach, N.S.; Mertz, B.; O’Malley, M.A.; Han, S. Homo-oligomerization of the human adenosine A(2A) receptor is driven by the intrinsically disordered C-terminus. eLife 2021, 10, e66662. [Google Scholar] [CrossRef]
- Kharche, S.A.; Sengupta, D. Dynamic protein interfaces and conformational landscapes of membrane protein complexes. Curr. Opin. Struct. Biol. 2020, 61, 191–197. [Google Scholar] [CrossRef]
- Bockaert, J.; Marin, P.; Dumuis, A.; Fagni, L. The ‘magic tail’ of G protein-coupled receptors: An anchorage for functional protein networks. FEBS Lett. 2003, 546, 65–72. [Google Scholar] [CrossRef]
- Petit-Pedrol, M.; Groc, L. Regulation of membrane NMDA receptors by dynamics and protein interactions. J. Cell Biol. 2021, 220, 6101. [Google Scholar] [CrossRef]
- Calebiro, D.; Koszegi, Z. The subcellular dynamics of GPCR signaling. Mol. Cell. Endocrinol. 2019, 483, 24–30. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Fuxe, K. Oligomeric Receptor Complexes and Their Allosteric Receptor-Receptor Interactions in the Plasma Membrane Represent a New Biological Principle for Integration of Signals in the CNS. Front. Mol. Neurosci. 2019, 12, 230. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. Historical review: A brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol. Sci. 2004, 25, 413–422. [Google Scholar] [CrossRef]
- Franco, R.; Martinez-Pinilla, E.; Lanciego, J.L.; Navarro, G. Basic Pharmacological and Structural Evidence for Class A G-Protein-Coupled Receptor Heteromerization. Front. Pharmacol. 2016, 7, 181687. [Google Scholar] [CrossRef]
- Hasbi, A.; O’Dowd, B.F.; George, S.R. Dopamine D1-D2 receptor heteromer signaling pathway in the brain: Emerging physiological relevance. Mol. Brain 2011, 4, 26. [Google Scholar] [CrossRef]
- Maroteaux, L.; Bechade, C.; Roumier, A. Dimers of serotonin receptors: Impact on ligand affinity and signaling. Biochimie 2019, 161, 23–33. [Google Scholar] [CrossRef]
- Ciruela, F.; Burgueno, J.; Casado, V.; Canals, M.; Marcellino, D.; Goldberg, S.R.; Bader, M.; Fuxe, K.; Agnati, L.F.; Lluis, C.; et al. Combining mass spectrometry and pull-down techniques for the study of receptor heteromerization. Direct epitope-epitope electrostatic interactions between adenosine A2A and dopamine D2 receptors. Anal. Chem. 2004, 76, 5354–5363. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Marcellino, D.; Narvaez, M.; Flajolet, M.; Heintz, N.; Agnati, L.; Ciruela, F.; Fuxe, K. A serine point mutation in the adenosine A2AR C-terminal tail reduces receptor heteromerization and allosteric modulation of the dopamine D2R. Biochem. Biophys. Res. Commun. 2010, 394, 222–227. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Rodriguez, D.; Romero-Fernandez, W.; Kapla, J.; Jaiteh, M.; Ranganathan, A.; Lazarova, T.; Fuxe, K.; Carlsson, J. Mapping the Interface of a GPCR Dimer: A Structural Model of the A2A Adenosine and D2 Dopamine Receptor Heteromer. Front. Pharmacol. 2018, 9, 829. [Google Scholar] [CrossRef]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Jockers, R. Do orphan G-protein-coupled receptors have ligand-independent functions? New insights from receptor heterodimers. EMBO Rep. 2006, 7, 1094–1098. [Google Scholar] [CrossRef]
- Bueschbell, B.; Manga, P.; Penner, E.; Schiedel, A.C. Evidence for Protein-Protein Interaction between Dopamine Receptors and the G Protein-Coupled Receptor 143. Int. J. Mol. Sci. 2021, 22, 8328. [Google Scholar] [CrossRef]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Couturier, C.; Delagrange, P.; Jockers, R. The orphan GPR50 receptor specifically inhibits MT1 melatonin receptor function through heterodimerization. EMBO J. 2006, 25, 3012–3023. [Google Scholar] [CrossRef]
- Balenga, N.A.; Martinez-Pinilla, E.; Kargl, J.; Schroder, R.; Peinhaupt, M.; Platzer, W.; Balint, Z.; Zamarbide, M.; Dopeso-Reyes, I.G.; Ricobaraza, A.; et al. Heteromerization of GPR55 and cannabinoid CB2 receptors modulates signalling. Br. J. Pharmacol. 2014, 171, 5387–5406. [Google Scholar] [CrossRef]
- Reyes-Resina, I.; Navarro, G.; Aguinaga, D.; Canela, E.I.; Schoeder, C.T.; Zaluski, M.; Kiec-Kononowicz, K.; Saura, C.A.; Muller, C.E.; Franco, R. Molecular and functional interaction between GPR18 and cannabinoid CB(2) G-protein-coupled receptors. Relevance in neurodegenerative diseases. Biochem. Pharmacol. 2018, 157, 169–179. [Google Scholar] [CrossRef]
- Szymanska, K.; Kalafut, J.; Przybyszewska, A.; Paziewska, B.; Adamczuk, G.; Kielbus, M.; Rivero-Muller, A. FSHR Trans-Activation and Oligomerization. Front. Endocrinol. 2018, 9, 760. [Google Scholar] [CrossRef]
- Quitterer, U.; AbdAlla, S. Discovery of Pathologic GPCR Aggregation. Front. Med. 2019, 6, 9. [Google Scholar] [CrossRef]
- AbdAlla, S.; Lother, H.; Quitterer, U. AT1-receptor heterodimers show enhanced G-protein activation and altered receptor sequestration. Nature 2000, 407, 94–98. [Google Scholar] [CrossRef]
- Gill, K.S.; Mehta, K.; Heredia, J.D.; Krishnamurthy, V.V.; Zhang, K.; Procko, E. Multiple mechanisms of self-association of chemokine receptors CXCR4 and CCR5 demonstrated by deep mutagenesis. J. Biol. Chem. 2023, 299, 105229. [Google Scholar] [CrossRef]
- Contento, R.L.; Molon, B.; Boularan, C.; Pozzan, T.; Manes, S.; Marullo, S.; Viola, A. CXCR4-CCR5: A couple modulating T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 10101–10106. [Google Scholar] [CrossRef]
- Andrianarivelo, A.; Saint-Jour, E.; Walle, R.; Trifilieff, P.; Vanhoutte, P. Modulation and functions of dopamine receptor heteromers in drugs of abuse-induced adaptations. Neuropharmacology 2019, 152, 42–50. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chu, X.P.; Mao, L.M.; Wang, M.; Lan, H.X.; Li, M.H.; Zhang, G.C.; Parelkar, N.K.; Fibuch, E.E.; Haines, M.; et al. Modulation of D2R-NR2B interactions in response to cocaine. Neuron 2006, 52, 897–909. [Google Scholar] [CrossRef]
- Tanganelli, S.; Antonelli, T.; Tomasini, M.C.; Beggiato, S.; Fuxe, K.; Ferraro, L. Relevance of dopamine D(2)/neurotensin NTS1 and NMDA/neurotensin NTS1 receptor interaction in psychiatric and neurodegenerative disorders. Curr. Med. Chem. 2012, 19, 304–316. [Google Scholar] [CrossRef]
- Marino, M.J.; Rouse, S.T.; Levey, A.I.; Potter, L.T.; Conn, P.J. Activation of the genetically defined m1 muscarinic receptor potentiates N-methyl-D-aspartate (NMDA) receptor currents in hippocampal pyramidal cells. Proc. Natl. Acad. Sci. USA 1998, 95, 11465–11470. [Google Scholar] [CrossRef]
- Altier, C.; Zamponi, G.W. Analysis of GPCR/ion channel interactions. Methods Mol. Biol. 2011, 756, 215–225. [Google Scholar] [CrossRef]
- Hermosilla, T.; Moreno, C.; Itfinca, M.; Altier, C.; Armisen, R.; Stutzin, A.; Zamponi, G.W.; Varela, D. L-type calcium channel beta subunit modulates angiotensin II responses in cardiomyocytes. Channels 2011, 5, 280–286. [Google Scholar] [CrossRef]
- Ambrogini, P.; Lattanzi, D.; Pagliarini, M.; Di Palma, M.; Sartini, S.; Cuppini, R.; Fuxe, K.; Borroto-Escuela, D.O. 5HT1AR-FGFR1 Heteroreceptor Complexes Differently Modulate GIRK Currents in the Dorsal Hippocampus and the Dorsal Raphe Serotonin Nucleus of Control Rats and of a Genetic Rat Model of Depression. Int. J. Mol. Sci. 2023, 24, 7467. [Google Scholar] [CrossRef] [PubMed]
- Doupnik, C.A. GPCR-Kir channel signaling complexes: Defining rules of engagement. J. Recept. Signal Transduct. Res. 2008, 28, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Sahlholm, K.; Nilsson, J.; Marcellino, D.; Fuxe, K.; Arhem, P. The human histamine H3 receptor couples to GIRK channels in Xenopus oocytes. Eur. J. Pharmacol. 2007, 567, 206–210. [Google Scholar] [CrossRef]
- Chen, R.; Ferris, M.J.; Wang, S. Dopamine D2 autoreceptor interactome: Targeting the receptor complex as a strategy for treatment of substance use disorder. Pharmacol. Ther. 2020, 213, 107583. [Google Scholar] [CrossRef]
- Quarta, D.; Ciruela, F.; Patkar, K.; Borycz, J.; Solinas, M.; Lluis, C.; Franco, R.; Wise, R.A.; Goldberg, S.R.; Hope, B.T.; et al. Heteromeric nicotinic acetylcholine-dopamine autoreceptor complexes modulate striatal dopamine release. Neuropsychopharmacology 2007, 32, 35–42. [Google Scholar] [CrossRef]
- Bontempi, L.; Savoia, P.; Bono, F.; Fiorentini, C.; Missale, C. Dopamine D3 and acetylcholine nicotinic receptor heteromerization in midbrain dopamine neurons: Relevance for neuroplasticity. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2017, 27, 313–324. [Google Scholar] [CrossRef]
- Di Palma, M.; Sartini, S.; Lattanzi, D.; Cuppini, R.; Pita-Rodriguez, M.; Diaz-Carmenate, Y.; Narvaez, M.; Fuxe, K.; Borroto-Escuela, D.O.; Ambrogini, P. Evidence for the existence of A2AR-TrkB heteroreceptor complexes in the dorsal hippocampus of the rat brain: Potential implications of A2AR and TrkB interplay upon ageing. Mech. Ageing Dev. 2020, 190, 111289. [Google Scholar] [CrossRef]
- Narvaez, M.; Andrade-Talavera, Y.; Valladolid-Acebes, I.; Fredriksson, M.; Siegele, P.; Hernandez-Sosa, A.; Fisahn, A.; Fuxe, K.; Borroto-Escuela, D.O. Existence of FGFR1-5-HT1AR heteroreceptor complexes in hippocampal astrocytes. Putative link to 5-HT and FGF2 modulation of hippocampal gamma oscillations. Neuropharmacology 2020, 170, 108070. [Google Scholar] [CrossRef]
- Di Liberto, V.; Borroto-Escuela, D.O.; Frinchi, M.; Verdi, V.; Fuxe, K.; Belluardo, N.; Mudo, G. Existence of muscarinic acetylcholine receptor (mAChR) and fibroblast growth factor receptor (FGFR) heteroreceptor complexes and their enhancement of neurite outgrowth in neural hippocampal cultures. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 235–245. [Google Scholar] [CrossRef]
- Fuxe, K.; Ferre, S.; Zoli, M.; Agnati, L.F. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A(2A) dopamine D-2 and adenosine A(1) dopamine D-1 receptor interactions in the basal ganglia. Brain Res. Rev. 1998, 26, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Torvinen, M.; Marcellino, D.; Canals, M.; Agnati, L.F.; Lluis, C.; Franco, R.; Fuxe, K. Adenosine A2A receptor and dopamine D3 receptor interactions: Evidence of functional A2A/D3 heteromeric complexes. Mol. Pharmacol. 2005, 67, 400–407. [Google Scholar] [CrossRef]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Guescini, M.; Fernandez-Duenas, V.; Tanganelli, S.; Rivera, A.; Ciruela, F.; Agnati, L.F. Adenosine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, e18–e42. [Google Scholar] [CrossRef] [PubMed]
- Franco, R. Neurotransmitter receptor heteromers in neurodegenerative diseases and neural plasticity. J. Neural Transm. 2009, 116, 983–987. [Google Scholar] [CrossRef] [PubMed]
- George, S.R.; Kern, A.; Smith, R.G.; Franco, R. Dopamine receptor heteromeric complexes and their emerging functions. Prog. Brain Res. 2014, 211, 183–200. [Google Scholar] [CrossRef]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Marcellino, D.; Ciruela, F.; Agnati, L.F.; Fuxe, K. Dopamine D2 and 5-hydroxytryptamine 5-HT((2)A) receptors assemble into functionally interacting heteromers. Biochem. Biophys. Res. Commun. 2010, 401, 605–610. [Google Scholar] [CrossRef]
- Kolasa, M.; Solich, J.; Faron-Gorecka, A.; Zurawek, D.; Pabian, P.; Lukasiewicz, S.; Kusmider, M.; Szafran-Pilch, K.; Szlachta, M.; Dziedzicka-Wasylewska, M. Paroxetine and Low-dose Risperidone Induce Serotonin 5-HT(1A) and Dopamine D2 Receptor Heteromerization in the Mouse Prefrontal Cortex. Neuroscience 2018, 377, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Narvaez, M.; Oflijan, J.; Agnati, L.F.; Fuxe, K. Hallucinogenic 5-HT2AR agonists LSD and DOI enhance dopamine D2R protomer recognition and signaling of D2-5-HT2A heteroreceptor complexes. Biochem. Biophys. Res. Commun. 2014, 443, 278–284. [Google Scholar] [CrossRef]
- Lukasiewicz, S.; Blasiak, E.; Szafran-Pilch, K.; Dziedzicka-Wasylewska, M. Dopamine D2 and serotonin 5-HT1A receptor interaction in the context of the effects of antipsychotics—In vitro studies. J. Neurochem. 2016, 137, 549–560. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Ambrogini, P.; Narvaez, M.; Di Liberto, V.; Beggiato, S.; Ferraro, L.; Fores-Pons, R.; Alvarez-Contino, J.E.; Lopez-Salas, A.; Mudo, G.; et al. Serotonin Heteroreceptor Complexes and Their Integration of Signals in Neurons and Astroglia-Relevance for Mental Diseases. Cells 2021, 10, 1902. [Google Scholar] [CrossRef]
- Fuxe, K.; Agnati, L.F.; Cintra, A.; Andersson, K.; Eneroth, P.; Harfstrand, A.; Zoli, M.; Goldstein, M. Studies on central D1 receptors role in volume transmission, neuroendrocrine regulation and release of noradrenaline. Adv. Exp. Med. Biol. 1988, 235, 83–119. [Google Scholar] [CrossRef]
- Bjelke, B.; Stromberg, I.; O’Connor, W.T.; Andbjer, B.; Agnati, L.F.; Fuxe, K. Evidence for volume transmission in the dopamine denervated neostriatum of the rat after a unilateral nigral 6-OHDA microinjection. Studies with systemic D-amphetamine treatment. Brain Res. 1994, 662, 11–24. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Agnati, L.F.; Bechter, K.; Jansson, A.; Tarakanov, A.O.; Fuxe, K. The role of transmitter diffusion and flow versus extracellular vesicles in volume transmission in the brain neural-glial networks. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140183. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.O. Volume transmission and receptor-receptor interactions in heteroreceptor complexes: Understanding the role of new concepts for brain communication. Neural Regen. Res. 2016, 11, 1220–1223. [Google Scholar] [CrossRef]
- Agnati, L.F.; Cortelli, P.; Biagini, G.; Bjelke, B.; Fuxe, K. Different classes of volume transmission signals exist in the central nervous system and are affected by metabolic signals, temperature gradients and pressure waves. Neuroreport 1994, 6, 9–12. [Google Scholar] [CrossRef]
- Katz, P.S.; Edwards, D.H. Metamodulation: The control and modulation of neuromodulationGet accessArrow. In Beyond Neurotransmission: Neuromodulation and Its Importance for Information Processing; Katz, P.S., Ed.; Oxford University Press: Oxford, UK, 1999; pp. 349–382. [Google Scholar]
- Philpot, B.D.; Bear, M.F.; Abraham, W.C. Metaplasticity: The plasticity of synaptic plasticity. In Beyond Neurotransmission: Neuromodulation and Its Importance for Information Processing; Katz, P.S., Ed.; Oxford University Press: Oxford, UK, 1999; pp. 160–197. [Google Scholar]
- Fuxe, K.; Agnati, L.F. Receptor-receptor interactions in the central nervous system. A new integrative mechanism in synapses. Med. Res. Rev. 1985, 5, 441–482. [Google Scholar] [CrossRef]
- Ribeiro, J.A.; Sebastiao, A.M. Modulation and metamodulation of synapses by adenosine. Acta Physiol. 2010, 199, 161–169. [Google Scholar] [CrossRef]
- Sebastiao, A.M.; Ribeiro, J.A. Tuning and fine-tuning of synapses with adenosine. Curr. Neuropharmacol. 2009, 7, 180–194. [Google Scholar] [CrossRef]
- Sebastiao, A.M.; Ribeiro, J.A. Neuromodulation and metamodulation by adenosine: Impact and subtleties upon synaptic plasticity regulation. Brain Res. 2015, 1621, 102–113. [Google Scholar] [CrossRef]
- Fields, R.D.; Burnstock, G. Purinergic signalling in neuron-glia interactions. Nat. Reviews. Neurosci. 2006, 7, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Ciruela, F.; Casado, V.; Rodrigues, R.J.; Lujan, R.; Burgueno, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef] [PubMed]
- Cristovao-Ferreira, S.; Navarro, G.; Brugarolas, M.; Perez-Capote, K.; Vaz, S.H.; Fattorini, G.; Conti, F.; Lluis, C.; Ribeiro, J.A.; McCormick, P.J.; et al. A1R-A2AR heteromers coupled to Gs and G i/0 proteins modulate GABA transport into astrocytes. Purinergic Signal. 2013, 9, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Fores-Pons, R.; Vasudevan, L.; Romero-Fernandez, W.; Frankowska, M.; Ferraro, L.; Beggiato, S.; Crespo-Ramirez, M.; Rivera, A.; et al. The Balance of MU-Opioid, Dopamine D2 and Adenosine A2A Heteroreceptor Complexes in the Ventral Striatal-Pallidal GABA Antireward Neurons May Have a Significant Role in Morphine and Cocaine Use Disorders. Front. Pharmacol. 2021, 12, 627032. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Fuxe, K. Adenosine heteroreceptor complexes in the basal ganglia are implicated in Parkinson’s disease and its treatment. J. Neural Transm. 2019, 126, 455–471. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, A.; Cimadevila, M.; Cadavid, M.I.; Loza, M.I.; Brea, J. Serotonin-2A homodimers are needed for signalling via both phospholipase A2 and phospholipase C in transfected CHO cells. Eur. J. Pharmacol. 2017, 800, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Romero-Fernandez, W.; Wydra, K.; Borroto-Escuela, D.O.; Jastrzebska, J.; Zhou, Z.; Frankowska, M.; Filip, M.; Fuxe, K. Increased density and antagonistic allosteric interactions in A2AR-D2R heterocomplexes in extinction from cocaine use, lost in cue induced reinstatement of cocaine seeking. Pharmacol. Biochem. Behav. 2022, 215, 173375. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Wydra, K.; Filip, M.; Fuxe, K. A2AR-D2R Heteroreceptor Complexes in Cocaine Reward and Addiction. Trends Pharmacol. Sci. 2018, 39, 1008–1020. [Google Scholar] [CrossRef]
- Halberstadt, A.L.; Lehmann-Masten, V.D.; Geyer, M.A.; Powell, S.B. Interactive effects of mGlu5 and 5-HT2A receptors on locomotor activity in mice. Psychopharmacology 2011, 215, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Agnati, L.F.; Marcoli, M.; Borroto-Escuela, D.O. Volume Transmission in Central Dopamine and Noradrenaline Neurons and Its Astroglial Targets. Neurochem. Res. 2015, 40, 2600–2614. [Google Scholar] [CrossRef]
- Taddeucci, A.; Olivero, G.; Roggeri, A.; Milanese, C.; Giorgio, F.P.D.; Grilli, M.; Marchi, M.; Garrone, B.; Pittaluga, A. Presynaptic 5-HT(2A)-mGlu2/3 Receptor-Receptor Crosstalk in the Prefrontal Cortex: Metamodulation of Glutamate Exocytosis. Cells 2022, 11. [Google Scholar] [CrossRef]
- Pittaluga, A.; Marchi, M. Synaptosomes and Metamodulation of Receptors. Methods Mol. Biol. 2022, 2417, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Borroto-Escuela, D.O.; Ciruela, F.; Guidolin, D.; Agnati, L.F. Receptor-receptor interactions in heteroreceptor complexes: A new principle in biology. Focus on their role in learning and memory. Neurosci. Discov. 2014, 2. [Google Scholar] [CrossRef]
- Pittaluga, A.; Roggeri, A.; Vallarino, G.; Olivero, G. Somatostatin, a Presynaptic Modulator of Glutamatergic Signal in the Central Nervous System. Int. J. Mol. Sci. 2021, 22, 5864. [Google Scholar] [CrossRef] [PubMed]
- Agnati, L.F.; Fuxe, K.; Zoli, M.; Rondanini, C.; Ogren, S.O. New vistas on synaptic plasticity: The receptor mosaic hypothesis of the engram. Med. Biol. 1982, 60, 183–190. [Google Scholar] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Pintsuk, J.; Narvaez, M.; Corrales, F.; Zaniewska, M.; Agnati, L.F.; Franco, R.; Tanganelli, S.; Ferraro, L.; et al. Understanding the Functional Plasticity in Neural Networks of the Basal Ganglia in Cocaine Use Disorder: A Role for Allosteric Receptor-Receptor Interactions in A2A-D2 Heteroreceptor Complexes. Neural Plast. 2016, 2016, 4827268. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Neural synchrony in brain disorders: Relevance for cognitive dysfunctions and pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Ambrogini, P.; Chruscicka, B.; Lindskog, M.; Crespo-Ramirez, M.; Hernandez-Mondragon, J.C.; Perez de la Mora, M.; Schellekens, H.; Fuxe, K. The Role of Central Serotonin Neurons and 5-HT Heteroreceptor Complexes in the Pathophysiology of Depression: A Historical Perspective and Future Prospects. Int. J. Mol. Sci. 2021, 22, 1927. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R.; Young, L.J. The neurobiology of attachment. Nat. Rev. Neurosci. 2001, 2, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Young, L.J.; Lim, M.M.; Gingrich, B.; Insel, T.R. Cellular mechanisms of social attachment. Horm. Behav. 2001, 40, 133–138. [Google Scholar] [CrossRef]
- Schellekens, H.; Dinan, T.G.; Cryan, J.F. Taking two to tango: A role for ghrelin receptor heterodimerization in stress and reward. Front. Neurosci. 2013, 7, 148. [Google Scholar] [CrossRef]
- De la Mora, M.P.; Perez-Carrera, D.; Crespo-Ramirez, M.; Tarakanov, A.; Fuxe, K.; Borroto-Escuela, D.O. Signaling in dopamine D2 receptor-oxytocin receptor heterocomplexes and its relevance for the anxiolytic effects of dopamine and oxytocin interactions in the amygdala of the rat. Biochim. Biophys. Acta 2016, 1862, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Albizu, L.; Moreno, J.L.; Gonzalez-Maeso, J.; Sealfon, S.C. Heteromerization of G protein-coupled receptors: Relevance to neurological disorders and neurotherapeutics. CNS Neurol. Disord. Drug Targets 2010, 9, 636–650. [Google Scholar] [CrossRef]
- Gonzalez-Maeso, J.; Ang, R.L.; Yuen, T.; Chan, P.; Weisstaub, N.V.; Lopez-Gimenez, J.F.; Zhou, M.; Okawa, Y.; Callado, L.F.; Milligan, G.; et al. Identification of a serotonin/glutamate receptor complex implicated in psychosis. Nature 2008, 452, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Baki, L.; Fribourg, M.; Younkin, J.; Eltit, J.M.; Moreno, J.L.; Park, G.; Vysotskaya, Z.; Narahari, A.; Sealfon, S.C.; Gonzalez-Maeso, J.; et al. Cross-signaling in metabotropic glutamate 2 and serotonin 2A receptor heteromers in mammalian cells. Pflug. Arch. Eur. J. Physiol. 2016, 468, 775–793. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.L.; Miranda-Azpiazu, P.; Garcia-Bea, A.; Younkin, J.; Cui, M.; Kozlenkov, A.; Ben-Ezra, A.; Voloudakis, G.; Fakira, A.K.; Baki, L.; et al. Allosteric signaling through an mGlu2 and 5-HT2A heteromeric receptor complex and its potential contribution to schizophrenia. Sci. Signal. 2016, 9, ra5. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.L.; Muguruza, C.; Umali, A.; Mortillo, S.; Holloway, T.; Pilar-Cuellar, F.; Mocci, G.; Seto, J.; Callado, L.F.; Neve, R.L.; et al. Identification of three residues essential for 5-hydroxytryptamine 2A-metabotropic glutamate 2 (5-HT2A.mGlu2) receptor heteromerization and its psychoactive behavioral function. J. Biol. Chem. 2012, 287, 44301–44319. [Google Scholar] [CrossRef] [PubMed]
- Burnat, G.; Branski, P.; Solich, J.; Kolasa, M.; Chruscicka, B.; Dziedzicka-Wasylewska, M.; Pilc, A. The functional cooperation of 5-HT(1A) and mGlu4R in HEK-293 cell line. Pharmacol. Rep. 2020, 72, 1358–1369. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Romero-Fernandez, W.; Zhou, Z.; Frankowska, M.; Filip, M.; Fuxe, K. A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration. Int. J. Mol. Sci. 2019, 20, 6100. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Li, X.; Rodriguez, D.; Carlsson, J.; Jastrzebska, J.; Filip, M.; Fuxe, K. Disruption of A2AR-D2R Heteroreceptor Complexes After A2AR Transmembrane 5 Peptide Administration Enhances Cocaine Self-Administration in Rats. Mol. Neurobiol. 2018, 55, 7038–7048. [Google Scholar] [CrossRef]
- Kourrich, S.; Su, T.P.; Fujimoto, M.; Bonci, A. The sigma-1 receptor: Roles in neuronal plasticity and disease. Trends Neurosci. 2012, 35, 762–771. [Google Scholar] [CrossRef]
- Beggiato, S.; Borelli, A.C.; Borroto-Escuela, D.; Corbucci, I.; Tomasini, M.C.; Marti, M.; Antonelli, T.; Tanganelli, S.; Fuxe, K.; Ferraro, L. Cocaine modulates allosteric D2-sigma1 receptor-receptor interactions on dopamine and glutamate nerve terminals from rat striatum. Cell. Signal. 2017, 40, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Narvaez, M.; Wydra, K.; Pintsuk, J.; Pinton, L.; Jimenez-Beristain, A.; Di Palma, M.; Jastrzebska, J.; Filip, M.; Fuxe, K. Cocaine self-administration specifically increases A2AR-D2R and D2R-sigma1R heteroreceptor complexes in the rat nucleus accumbens shell. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2017, 155, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Wydra, K.; Zhou, Z.; Suder, A.; Filip, M.; Fuxe, K. OSU-6162, a Sigma1R Ligand in Low Doses, Can Further Increase the Effects of Cocaine Self-Administration on Accumbal D2R Heteroreceptor Complexes. Neurotox. Res. 2020, 37, 433–444. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Fuxe, K. The integrative role of G protein-coupled receptor heterocomplexes in Parkinson’s disease. Neural Regen. Res. 2022, 17, 2211–2212. [Google Scholar] [CrossRef]
- Aguirre, J.A.; Andbjer, B.; Gonzalez-Baron, S.; Hansson, A.; Stromberg, I.; Agnati, L.F.; Fuxe, K. Group I mGluR antagonist AIDA protects nigral DA cells from MPTP-induced injury. Neuroreport 2001, 12, 2615–2617. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T.; Fujinaga, M.; Kawamura, K.; Furutsuka, K.; Nengaki, N.; Shimoda, Y.; Shiomi, S.; Takei, M.; Hashimoto, H.; Yui, J.; et al. Dynamic Changes in Striatal mGluR1 But Not mGluR5 during Pathological Progression of Parkinson’s Disease in Human Alpha-Synuclein A53T Transgenic Rats: A Multi-PET Imaging Study. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 375–384. [Google Scholar] [CrossRef]
- Ferreira, D.G.; Batalha, V.L.; Vicente Miranda, H.; Coelho, J.E.; Gomes, R.; Goncalves, F.Q.; Real, J.I.; Rino, J.; Albino-Teixeira, A.; Cunha, R.A.; et al. Adenosine A2A Receptors Modulate alpha-Synuclein Aggregation and Toxicity. Cereb. Cortex 2017, 27, 718–730. [Google Scholar] [CrossRef]
- Sulzer, D.; Surmeier, D.J. Neuronal vulnerability, pathogenesis, and Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2013, 28, 41–50. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef]
- Perez de la Mora, M.; Hernandez-Mondragon, C.; Crespo-Ramirez, M.; Rejon-Orantes, J.; Borroto-Escuela, D.O.; Fuxe, K. Conventional and Novel Pharmacological Approaches to Treat Dopamine-Related Disorders: Focus on Parkinson’s Disease and Schizophrenia. Neuroscience 2020, 439, 301–318. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Perez De La Mora, M.; Manger, P.; Narvaez, M.; Beggiato, S.; Crespo-Ramirez, M.; Navarro, G.; Wydra, K.; Diaz-Cabiale, Z.; Rivera, A.; et al. Brain Dopamine Transmission in Health and Parkinson’s Disease: Modulation of Synaptic Transmission and Plasticity Through Volume Transmission and Dopamine Heteroreceptors. Front. Synaptic Neurosci. 2018, 10, 20. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirchandani-Duque, M.; Choucri, M.; Hernández-Mondragón, J.C.; Crespo-Ramírez, M.; Pérez-Olives, C.; Ferraro, L.; Franco, R.; Pérez de la Mora, M.; Fuxe, K.; Borroto-Escuela, D.O. Membrane Heteroreceptor Complexes as Second-Order Protein Modulators: A Novel Integrative Mechanism through Allosteric Receptor–Receptor Interactions. Membranes 2024, 14, 96. https://doi.org/10.3390/membranes14050096
Mirchandani-Duque M, Choucri M, Hernández-Mondragón JC, Crespo-Ramírez M, Pérez-Olives C, Ferraro L, Franco R, Pérez de la Mora M, Fuxe K, Borroto-Escuela DO. Membrane Heteroreceptor Complexes as Second-Order Protein Modulators: A Novel Integrative Mechanism through Allosteric Receptor–Receptor Interactions. Membranes. 2024; 14(5):96. https://doi.org/10.3390/membranes14050096
Chicago/Turabian StyleMirchandani-Duque, Marina, Malak Choucri, Juan C. Hernández-Mondragón, Minerva Crespo-Ramírez, Catalina Pérez-Olives, Luca Ferraro, Rafael Franco, Miguel Pérez de la Mora, Kjell Fuxe, and Dasiel O. Borroto-Escuela. 2024. "Membrane Heteroreceptor Complexes as Second-Order Protein Modulators: A Novel Integrative Mechanism through Allosteric Receptor–Receptor Interactions" Membranes 14, no. 5: 96. https://doi.org/10.3390/membranes14050096