Analysis of Olive (Olea Europaea L.) Genetic Resources in Relation to the Content of Vitamin E in Virgin Olive Oil

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
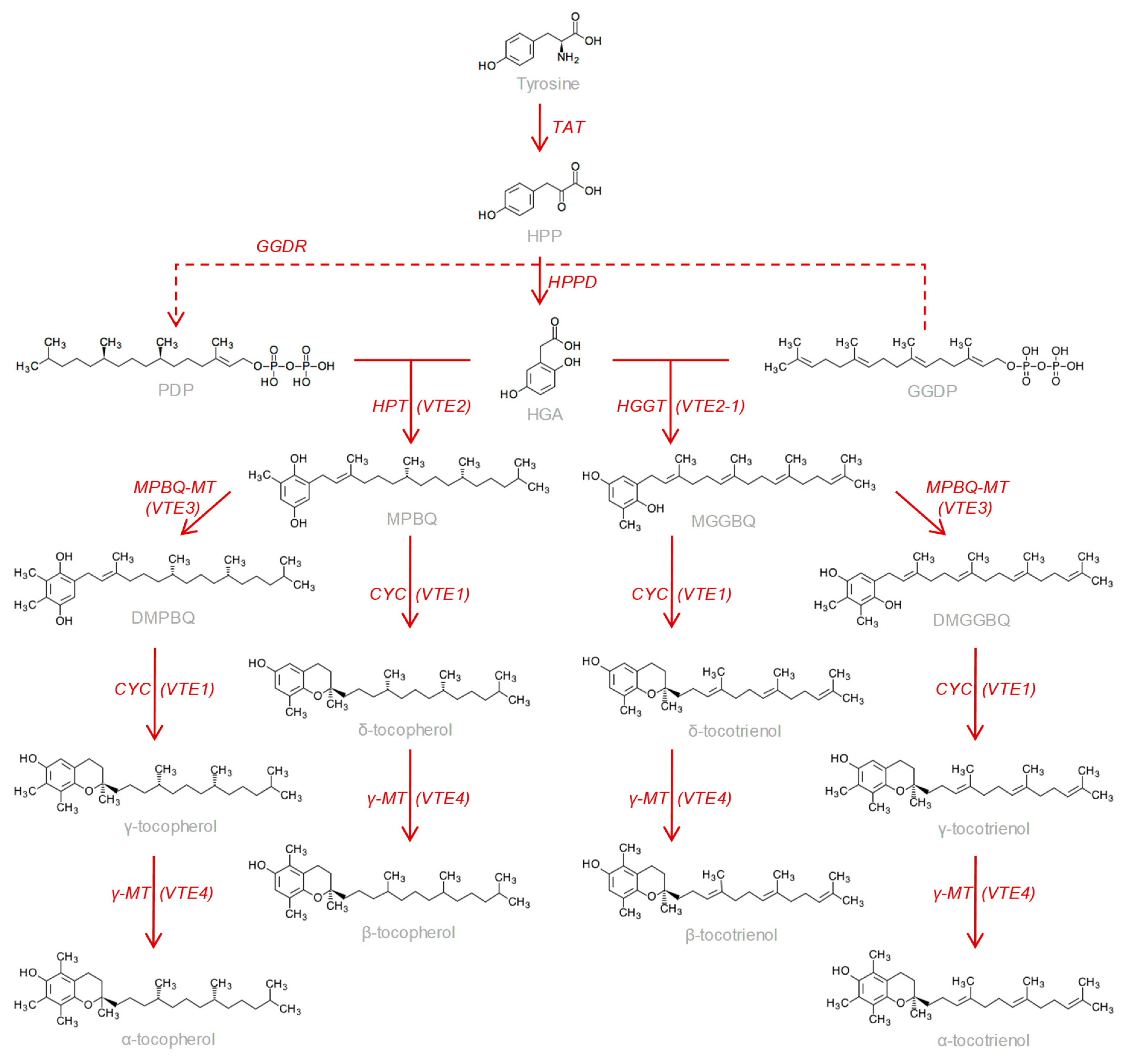
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Olive Oil Extraction
2.3. Analysis of Tocopherols
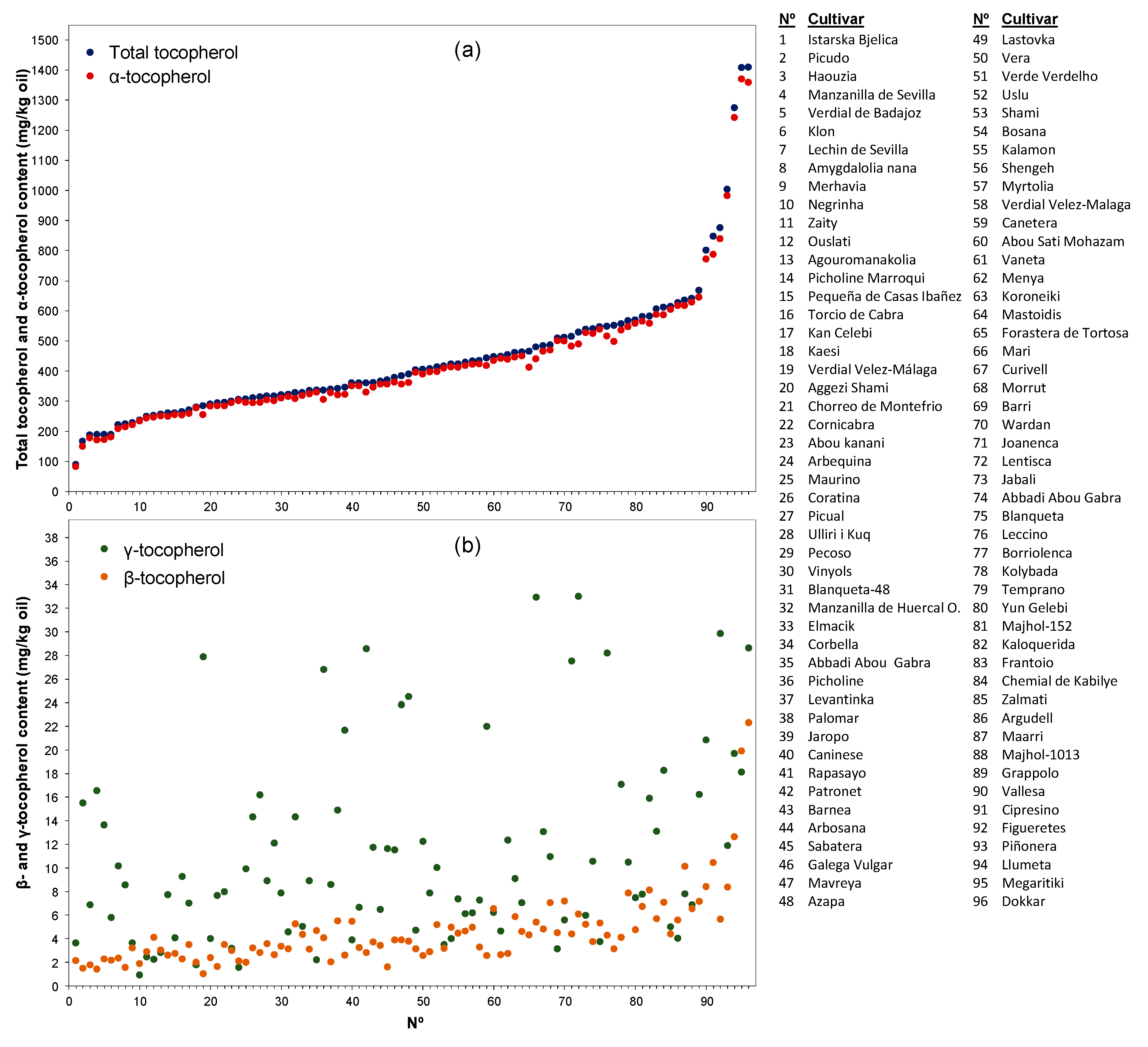
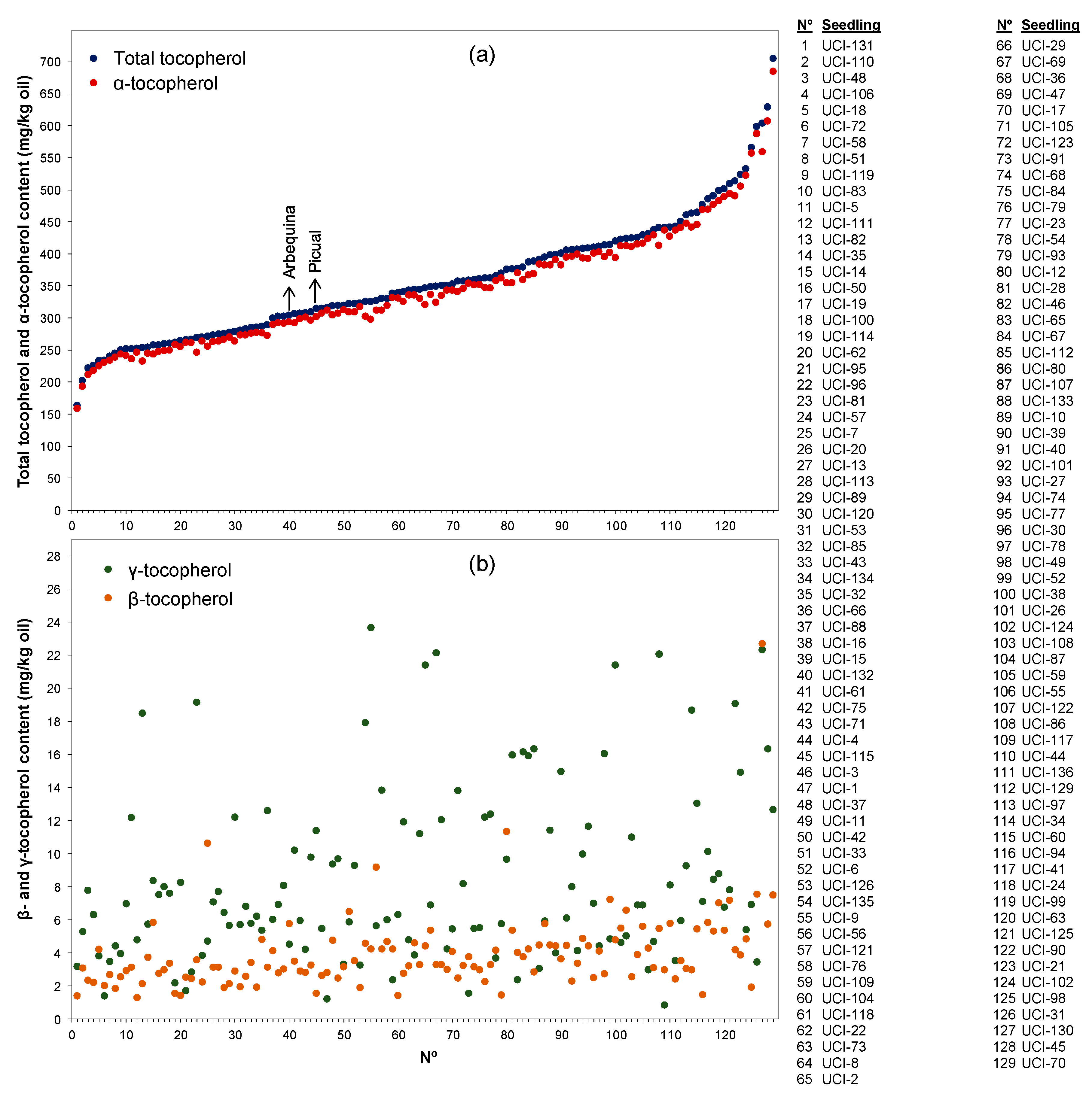
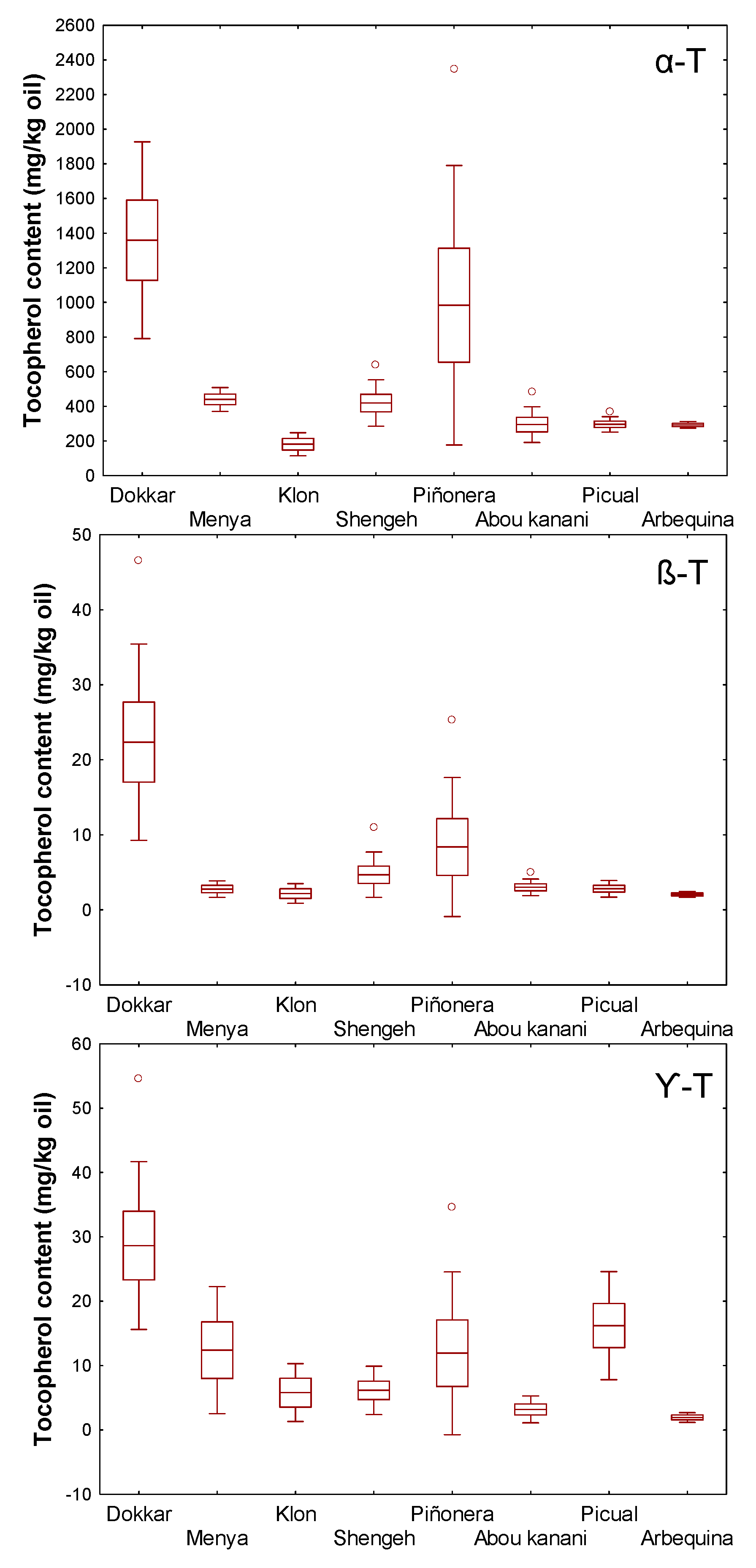
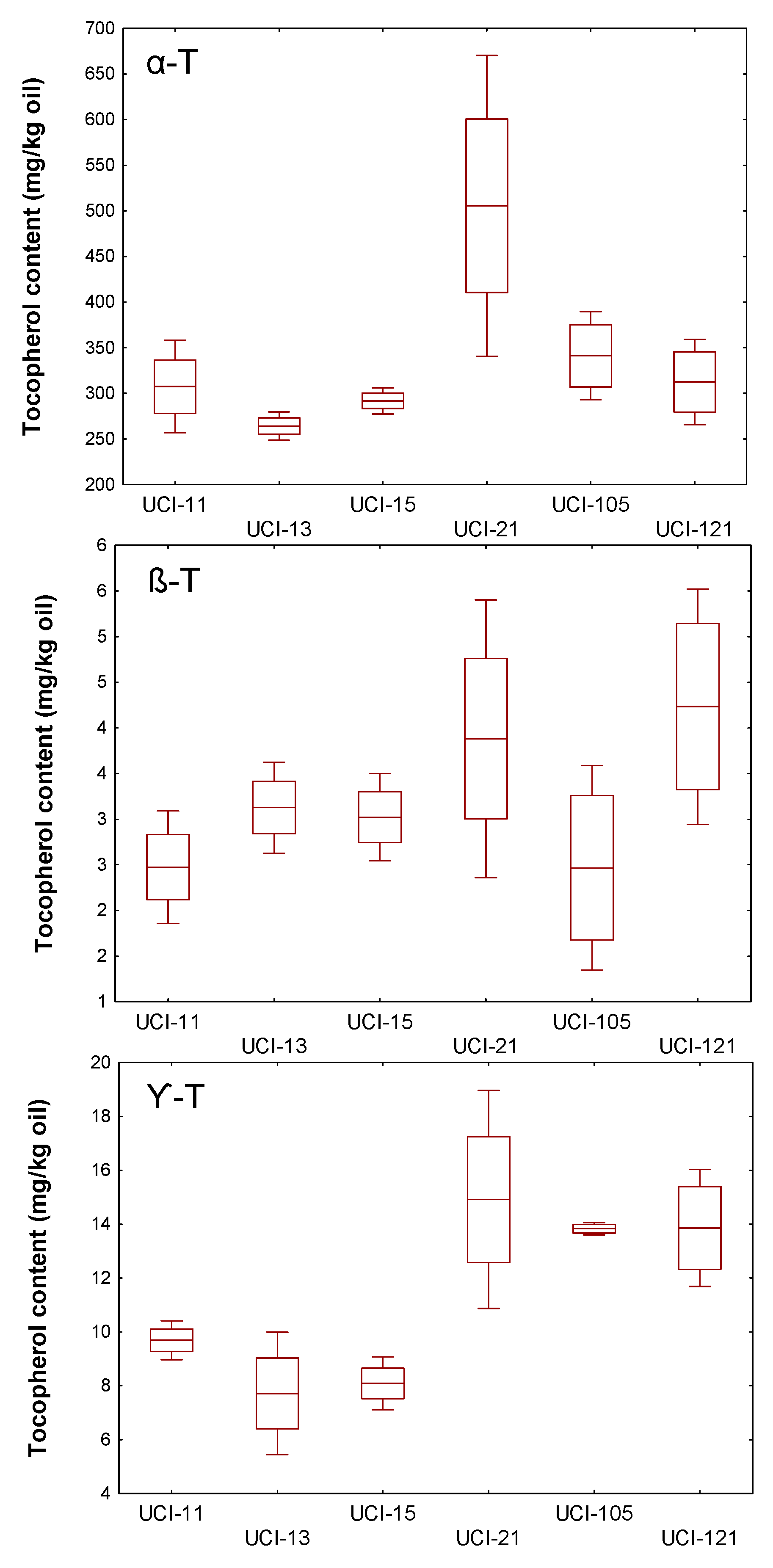
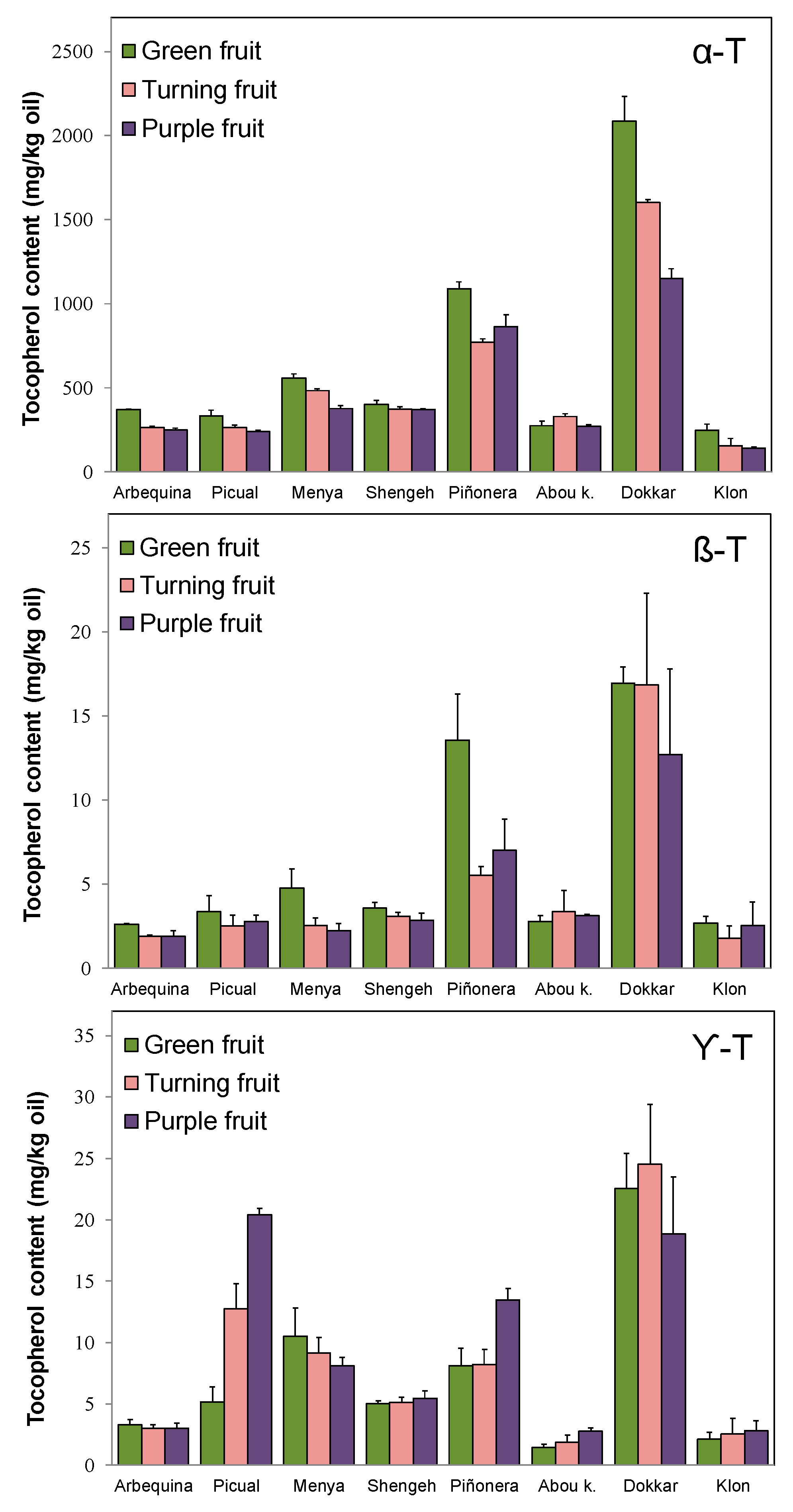
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Konstantinidou, V.; Covas, M.I.; Muñoz-Aguayo, D.; Khymenets, O.; de La Torre, R.; Saez, G.; Tormos Medel, C.; Toledo, E.; Martí, A.; Ruiz-Gutiérrez, V.; et al. In vivo nutrigenomic effects of VOO polyphenols within the frame of the Mediterranean diet: A randomized trial. FASEB J. 2010, 24, 2546–2557. [Google Scholar] [CrossRef] [PubMed]
- Lucas, L.; Russell, A.; Keast, R. Molecular mechanisms of inflammation. Anti-inflammatory benefits of virgin olive oil and the phenolic compound oleocanthal. Curr. Pharm. Des. 2011, 17, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Bernardini, E. Extra virgin olive oil’s polyphenols: Biological activities. Curr. Pharm. Des. 2011, 17, 786–804. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, R.; Belaj, A.; Romero-Segura, C.; Sanz, C.; Pérez, A.G. Exploration of genetic resources to improve the functional quality of virgin olive oil. J. Funct. Foods 2017, 38, 1–8. [Google Scholar] [CrossRef]
- Baldioli, M.; Servili, M.; Perretti, G.; Montedoro, G.F. Antioxidant activity of tocopherols and phenolic compounds of virgin olive oil. J. Am. Oil Chem. Soc. 1996, 73, 1589–1593. [Google Scholar] [CrossRef]
- Schneider, C. Chemistry and biology of vitamin E. Mol. Nutr. Food Res. 2005, 49, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Costarelli, L.; Giacconi, R.; Malavolta, M.; Basso, A.; Piacenza, F.; Ostan, R.; Cevenini, E.; Gonos, E.S.; Franceschi, C.; et al. Vitamin E-gene interactions in aging and inflammatory age-related diseases: Implications for treatment. A systematic review. Ageing Res. Rev. 2014, 14, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Kelly, F.J.; Salonen, J.T.; Neuzil, J.; Zingg, J.M.; Azzi, A. The European perspective on vitamin E: Current knowledge and future research. Am. J. Clin. Nutr. 2002, 76, 703–716. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies. Scientific opinion on the substantiation of health claims related to vitamin E. EFSA J. 2010, 8, 1816. Available online: https://efsa.onlinelibrary.wiley.com/doi/epdf/10.2903/j.efsa.2010.1816 (accessed on 25 July 2019).
- Ranalli, A.; Tombesi, A.; Ferrante, M.L.; De Mattia, G. Respiratory rate of olive drupes during their ripening cycle and quality of oil extracted. J. Sci. Food Agric. 1998, 77, 359–367. [Google Scholar] [CrossRef]
- Gutiérrez, F.; Jiménez, B.; Ruiz, A.; Albi, M.A. Effect of olive ripeness on the oxidative stability of virgin olive oil extracted from the varieties ‘Picual’ and Hojiblanca and on the different components involved. J. Agric. Food Chem. 1999, 47, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Beltrán, G.; Jiménez, A.; del Rio, C.; Sánchez, S.; Martínez, L.; Uceda, M.; Aguilera, M.P. Variability of vitamin E in virgin olive oil by agronomical and genetic factors. J. Food Comp. Anal. 2010, 23, 633–639. [Google Scholar] [CrossRef]
- Georgiadou, E.C.; Goulas, V.; Ntourou, T.; Manganaris, G.A.; Kalaitzis, P.; Fotopoulos, V. Regulation of on-tree vitamin e biosynthesis in olive fruit during successive growing years: The impact of fruit development and environmental cues. Front. Plant Sci. 2016, 7, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Georgiadou, E.C.; Koubouris, G.; Goulas, V.; Sergentani, C.; Nikoloudakis, N.; Manganaris, G.A.; Kalaitzis, P.; Fotopoulos, V. Genotype-dependent regulation of vitamin E biosynthesis in olive fruits as revealed through metabolic and transcriptional profiles. Plant Biol. 2019, 21, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Bondesan, L. Secoiridoids, tocopherols, and antioxidant activity of monovarietal extra virgin olive oils extracted from destoned fruits. J. Agric. Food Chem. 2005, 53, 1102–1107. [Google Scholar] [CrossRef]
- Beltrán, G.; Aguilera, M.P.; del Rio, C.; Sánchez, S.; Martínez, L. Influence of fruit ripening process on the natural antioxidant content of Hojiblanca virgin olive oils. Food Chem. 2005, 89, 207–215. [Google Scholar] [CrossRef]
- Inarejos-García, A.M.; Salvador, M.D.; Fregapane, G. Effect of crushing on olive paste and virgin olive oil minor components. Eur. Food Res. Technol. 2011, 232, 441–451. [Google Scholar] [CrossRef]
- Lavee, S. Aims, methods and advances in breeding of new olive (Olea europaea L.) cultivars. Acta Hortic. 1989, 286, 23–40. [Google Scholar] [CrossRef]
- León, L.; Beltrán, G.; Aguilera, M.P.; Rallo, L.; Barranco, D.; de la Rosa, R. Oil composition of advanced selections from an olive breeding program. Eur. J. Lipid Sci. Technol. 2011, 113, 870–875. [Google Scholar] [CrossRef]
- Rjiba, I.; Gazzah, N.; Dabbou, S.; Hammami, M. Evaluation of virgin olive oil minor compounds in progenies of controlled crosses. J. Food Biochem. 2011, 35, 1413–1423. [Google Scholar] [CrossRef]
- Pérez, A.G.; León, L.; Sanz, C.; de la Rosa, R. Fruit phenolic profiling: A new selection criterion in olive breeding programs. Front. Plant Sci. 2018, 9, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Belaj, A.; Domínguez-Garcia, M.C.; Atienza, S.G.; Martín-Urdiroz, N.; de la Rosa, R.; Satovic, Z.; Martin, A.; Kilian, A.; Trujillo, I.; Valpuesta, C.; et al. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Pérez, A.G.; León, L.; Pascual, M.; Romero-Segura, C.; Sánchez-Ortiz, A.; de la Rosa, R.; Sanz, C. Variability of virgin olive oil phenolic compounds in a segregating progeny from a single cross in Olea europaea L. and sensory and nutritional quality implications. PLoS ONE 2014, 9, 92898–92908. [Google Scholar] [CrossRef] [PubMed]
- Beltrán, G.; Uceda, M.; Hermoso, M.; Frías, L. Maduración. In El Cultivo del Olivo, 5th ed.; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; MundiPrensa-Junta de Andalucia: Madrid, Spain, 2004; pp. 156–183. [Google Scholar]
- Martínez, J.M.; Muñoz, E.; Alba, J.; Lanzón, A. Report about the use of the ‘Abencor’ analyser. Grasas Aceites 1975, 26, 379–385. [Google Scholar]
- IUPAC. Determination of tocopherols and tocotrienols in vegetable fats by HPLC. In Standard Methods of Analyses of Oils Fats and Derivatives, 7th ed.; Dieffenbacher, A., Pocklington, W.D., Eds.; Blackwell Science: Oxford, UK, 1992. [Google Scholar]
- Špika, M.J.; Kraljić, K.; Škevin, D. Tocopherols: Chemical structure, bioactivity, and variability in Croatian virgin olive oils. In Products from Olive Tree; Boskou, D., Clodoveo, M.L., Eds.; IntechOpen Limited: London, UK, 2016. [Google Scholar]
- Cunha, S.C.; Amaral, J.S.; Fernandes, J.O.; Oliveira, M.B. Quantification of tocopherols and tocotrienols in portuguese olive oils using HPLC with three different detection systems. J. Agric. Food Chem. 2006, 54, 3351–3356. [Google Scholar] [CrossRef] [PubMed]
- Baccouri, B.; Zarrouk, W.; Baccouri, O.; Guerfel, M.; Nouairi, I.; Krichene, D.; Daoud, D.; Zarrouk, M. Composition, quality and oxidative stability of virgin olive oils from some selected wild olives (Olea europaea L. subsp. Oleaster). Grasas Aceites 2008, 59, 346–351. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Hall, S.E.; Ripp, K.G.; Ganzke, T.S.; Hitz, W.D.; Coughlan, S.J. Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content. Nat. Biotechnol. 2003, 21, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; D’Agostino, N.; Torchia, L.; Servili, M.; Rao, R.; Pietrella, M.; Giuliano, G.; Chiusano, M.L.; Baldoni, L.; Perrotta, G. Comparative 454 pyrosequencing of transcripts from two olive genotypes during fruit development. BMC Genomics 2009, 10, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Uceda, M.; Hermoso, M.; Aguilera, M.P. La calidad del aceite de oliva. In El Cultivo del Olivo, 5th ed.; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; MundiPrensa-Junta de Andalucia: Madrid, Spain, 2004; pp. 656–684. [Google Scholar]
- Velasco, L.; de La Rosa, R.; León, L. Advanced olive selections with enhanced quality for minor constituents. Grasas Aceites 2015, 66, 100–107. [Google Scholar] [CrossRef]
- El Riachy, M.; Priego-Capote, F.; Rallo, L.; Luque de Castro, M.D.; León, L. Phenolic composition of virgin olive oils from cross breeding segregating populations. Eur. J. Lipid Sci. Technol. 2012, 114, 542–551. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez, A.G.; León, L.; Pascual, M.; de la Rosa, R.; Belaj, A.; Sanz, C. Analysis of Olive (Olea Europaea L.) Genetic Resources in Relation to the Content of Vitamin E in Virgin Olive Oil. Antioxidants 2019, 8, 242. https://doi.org/10.3390/antiox8080242
Pérez AG, León L, Pascual M, de la Rosa R, Belaj A, Sanz C. Analysis of Olive (Olea Europaea L.) Genetic Resources in Relation to the Content of Vitamin E in Virgin Olive Oil. Antioxidants. 2019; 8(8):242. https://doi.org/10.3390/antiox8080242
Chicago/Turabian StylePérez, Ana G., Lorenzo León, Mar Pascual, Raúl de la Rosa, Angjelina Belaj, and Carlos Sanz. 2019. "Analysis of Olive (Olea Europaea L.) Genetic Resources in Relation to the Content of Vitamin E in Virgin Olive Oil" Antioxidants 8, no. 8: 242. https://doi.org/10.3390/antiox8080242