Effect of Acute Ingestion of Green Tea Extract and Lemon Juice on Oxidative Stress and Lipid Profile in Pigs Fed a High-Fat Diet

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Treatments
2.3. Plasma Catechin Analysis
2.4. Plasma Cholesterol and Triglyceride
2.5. Liver Function Tests
2.6. Plasma Lipid Peroxidation
2.7. Activities of Plasma Antioxidant Enzymes
2.8. Statistical Analysis
3. Results
3.1. Plasma Catechin Level
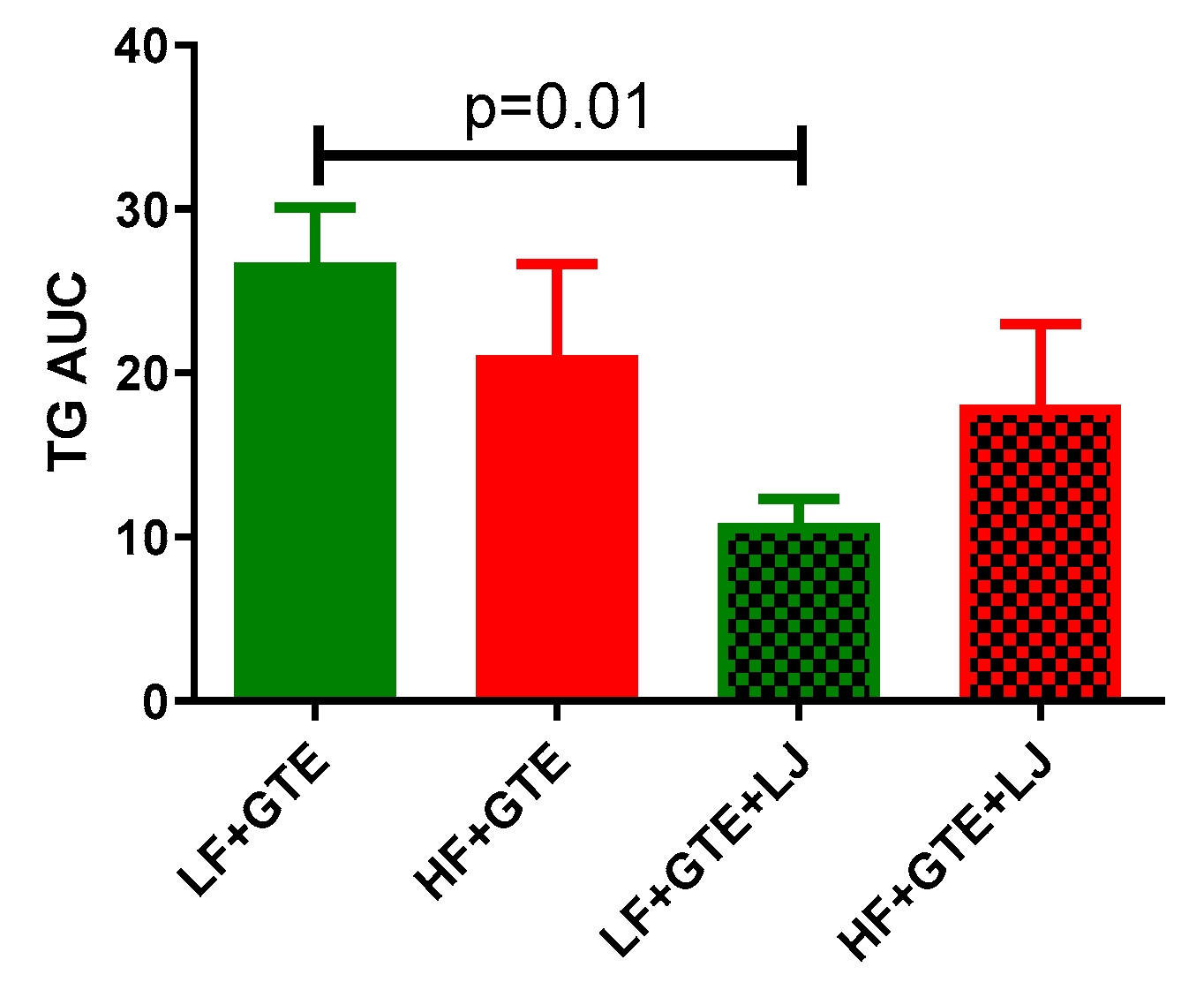
3.2. Plasma Lipid Profile
3.3. Liver Function Tests
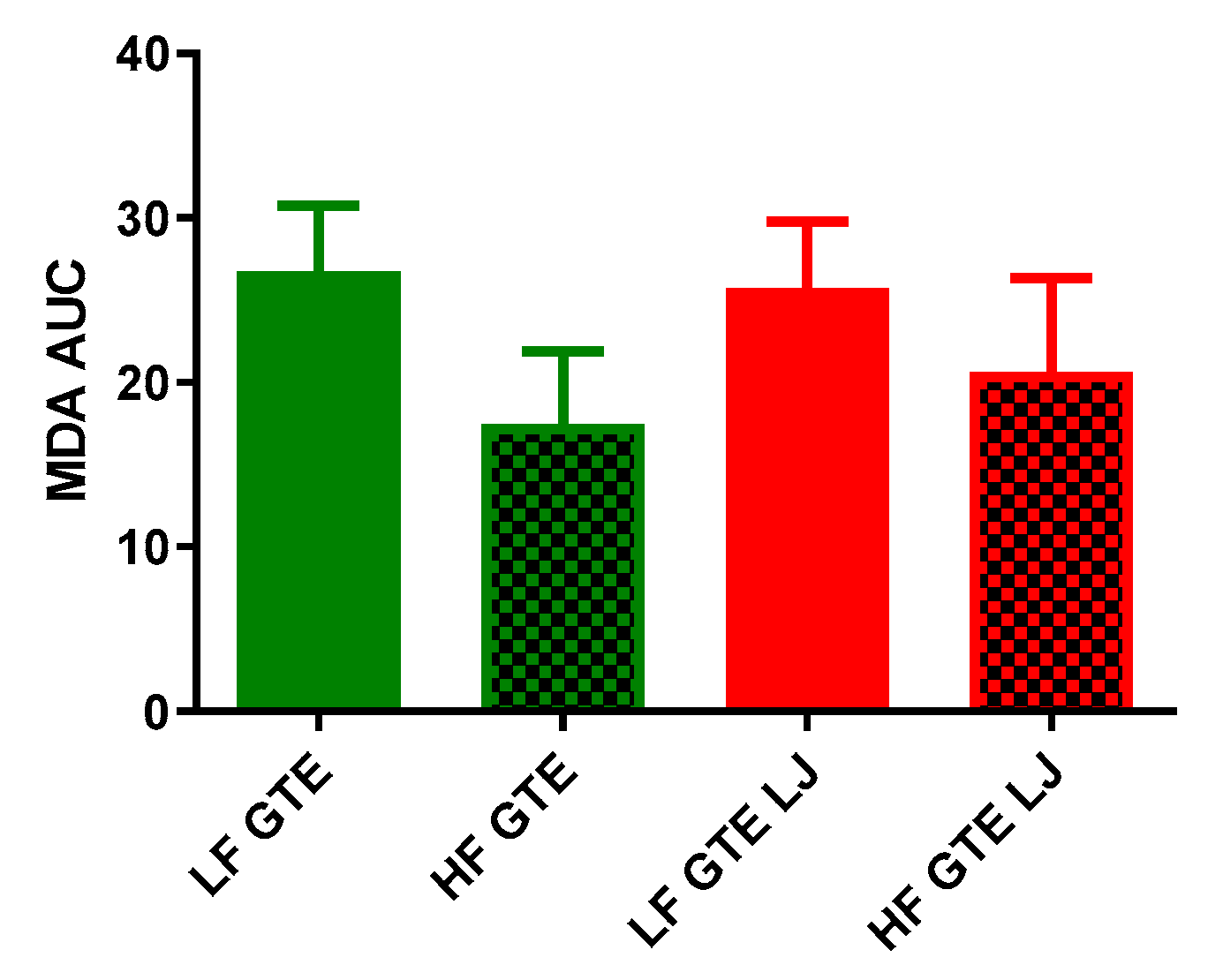
3.4. Plasma Lipid Peroxidation
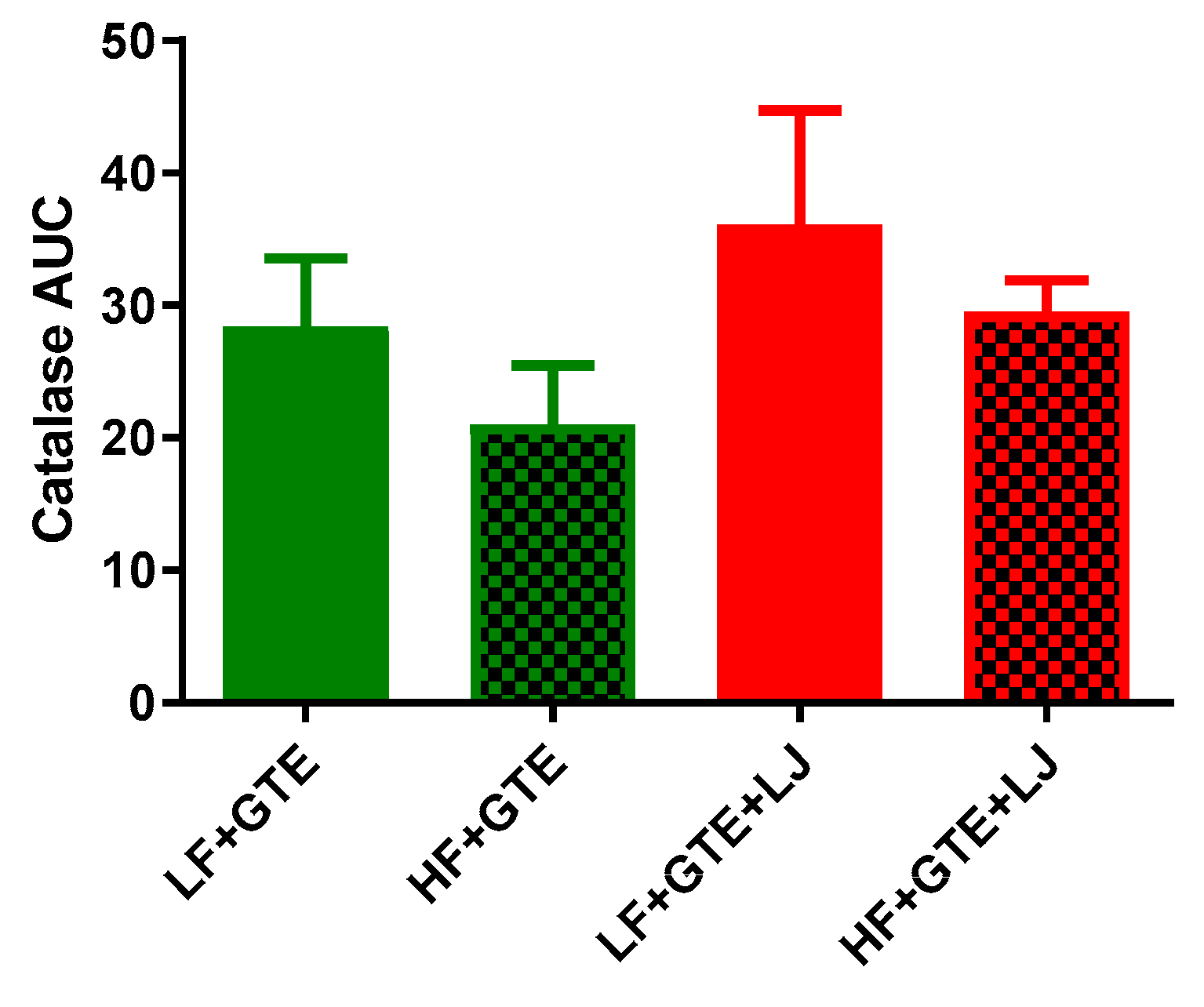
3.5. Activities of Plasma Antioxidant Enzymes
3.6. Correlation between Plasma Antioxidant Enzymes and Triglycerides
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef] [PubMed]
- Koloverou, E.; Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Georgousopoulou, E.N.; Grekas, A.; Christou, A.; Chatzigeorgiou, M.; Skoumas, I.; Tousoulis, D.; et al. Adherence to Mediterranean diet and 10-year incidence (2002–2012) of diabetes: Correlations with inflammatory and oxidative stress biomarkers in the ATTICA cohort study. Diabetes 2016, 32, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.I.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metabolism 2008, 57, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Peairs, A.D.; Rankin, J.W.; Lee, Y.W. Effects of acute ingestion of different fats on oxidative stress and inflammation in overweight and obese adults. Nutr. J. 2011, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katiyar, S.K.; Mukhtar, H. Green tea polyphenol (−)-epigallocatechin-3-gallate treatment to mouse skin prevents UVB-induced infiltration of leukocytes, depletion of antigen-presenting cells, and oxidative stress. J. Leukoc. Biol. 2001, 69, 719–726. [Google Scholar] [PubMed]
- Sabu, M.C.; Smitha, K.; Kuttan, R. Anti-diabetic activity of green tea polyphenols and their role in reducing oxidative stress in experimental diabetes. J. Ethnopharmacol. 2002, 83, 109–116. [Google Scholar] [PubMed]
- Tang, J.; Zheng, J.S.; Fang, L.; Jin, Y.X.; Cai, W.W.; Li, D. Tea consumption and mortality of all cancers, CVD and all causes: A meta-analysis of eighteen prospective cohort studies. Br. J. Nutr. 2015, 114, 673–683. [Google Scholar] [CrossRef]
- Posadino, A.M.; Phu, H.T.; Cossu, A.; Giordo, R.; Fois, M.; Thuan, D.T.B.; Piga, A.; Sotgia, S.; Zinellu, A.; Carru, C.; et al. Oxidative stress-induced Akt downregulation mediates green tea toxicity towards prostate cancer cells. Toxicol. In Vitro 2017, 42, 255–262. [Google Scholar] [CrossRef]
- Sharifzadeh, M.; Ranjbar, A.; Hosseini, A.; Khanavi, M. The Effect of Green Tea Extract on Oxidative Stress and Spatial Learning in Streptozotocin-diabetic Rats. Iran. J. Pharm. Res. 2017, 16, 201–209. [Google Scholar]
- Nogueira, L.D.; Neto, J.F.N.; Klein, M.R.S.T.; Sanjuliani, A.F. Short-term Effects of Green Tea on Blood Pressure, Endothelial Function, and Metabolic Profile in Obese Prehypertensive Women: A Crossover Randomized Clinical Trial. J. Am. Coll. Nutr. 2017, 36, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.J.; MacInnis, M.J.; Gillen, J.B.; Skelly, L.E.; Gibala, M.J. Short-term green tea extract supplementation attenuates the postprandial blood glucose and insulin response following exercise in overweight men. Appl. Physiol. Nutr. Metab. 2016, 41, 1057–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walkowiak, J.; Bajerska, J.; Kargulewicz, A.; Lisowska, A.; Siedlerski, G.; Szczapa, T.; Kobelska-Dubiel, N.; Grzymislawski, M. Single dose of green tea extract decreases lipid digestion and absorption from a test meal in humans. Acta. Biochim. Pol. 2013, 60, 481–483. [Google Scholar] [CrossRef]
- Martin, B.J.; Tan, R.B.; Gillen, J.B.; Percival, M.E.; Gibala, M.J. No Effect of Short-Term Green Tea Extract Supplementation on Metabolism at Rest or During Exercise in the Fed State. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 656–664. [Google Scholar] [CrossRef]
- Lonac, M.C.; Richards, J.C.; Schweder, M.M.; Johnson, T.K.; Bell, C. Influence of Short-Term Consumption of the Caffeine-Free, Epigallocatechin-3-Gallate Supplement, Teavigo, on Resting Metabolism and the Thermic Effect of Feeding. Obesity 2011, 19, 298–304. [Google Scholar] [CrossRef]
- Jowko, E.; Sacharuk, J.; Balasinska, B.; Wilczak, J.; Charmas, M.; Ostaszewski, P.; Charmas, R. Effect of a single dose of green tea polyphenols on the blood markers of exercise-induced oxidative stress in soccer players. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 486–496. [Google Scholar] [CrossRef]
- Masterjohn, C.; Bruno, R.S. Therapeutic potential of green tea in nonalcoholic fatty liver disease. Nutr. Rev. 2012, 70, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Green, R.J.; Murphy, A.S.; Schulz, B.; Watkins, B.A.; Ferruzzi, M.G. Common tea formulations modulate in vitro digestive recovery of green tea catechins. Mol. Nutr. Food Res. 2007, 51, 1152–1162. [Google Scholar] [CrossRef]
- Peters, C.M.; Green, R.J.; Janle, E.M.; Ferruzzi, M.G. Formulation with ascorbic acid and sucrose modulates catechin bioavailability from green tea. Food Res. Int. 2010, 43, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Hayasaka, S.; Shibata, Y.; Ojima, T.; Saegusa, T.; Gotoh, T.; Ishikawa, S.; Nakamura, Y.; Kayaba, K. Frequency of citrus fruit intake is associated with the incidence of cardiovascular disease: The Jichi Medical School cohort study. J. Epidemiol. 2011, 21, 169–175. [Google Scholar] [CrossRef]
- Chien, C.C.; Su, M.J. 5-hydroxytryptamine has an endothelium-derived hyperpolarizing factor-like effect on coronary flow in isolated rat hearts. J. Biomed. Sci. 2015, 22, 42. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Molina, E.; Dominguez-Perles, R.; Moreno, D.A.; Garcia-Viguera, C. Natural bioactive compounds of Citrus limon for food and health. J. Pharm. Biomed. Anal. 2010, 51, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Oikeh, E.I.; Omoregie, E.S.; Oviasogie, F.E.; Oriakhi, K. Phytochemical, antimicrobial, and antioxidant activities of different citrus juice concentrates. Food Sci. Nutr. 2016, 4, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.K.; Lei, X.G.; Miller, D.D. The pig as an experimental model for elucidating the mechanisms governing dietary influence on mineral absorption. Exp. Biol. Med. 2008, 233, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K. Advances in swine biomedical model genomics. Int. J. Biol. Sci. 2007, 3, 179–184. [Google Scholar] [CrossRef]
- National Research Council (U.S.); Committee on Nutrient Requirements of Swine. Nutrient Requirements of Swine, 11th rev. ed.; National Academies Press: Washington, DC, USA, 2012; p. xvii. [Google Scholar]
- Nagao, T.; Komine, Y.; Soga, S.; Meguro, S.; Hase, T.; Tanaka, Y.; Tokimitsu, I. Ingestion of a tea rich in catechins leads to a reduction in body fat and malondialdehyde-modified LDL in men. Am. J. Clin. Nutr. 2005, 81, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Maki, K.C.; Reeves, M.S.; Farmer, M.; Yasunaga, K.; Matsuo, N.; Katsuragi, Y.; Komikado, M.; Tokimitsu, I.; Wilder, D.; Jones, F.; et al. Green tea catechin consumption enhances exercise-induced abdominal fat loss in overweight and obese adults. J. Nutr. 2009, 139, 264–270. [Google Scholar] [CrossRef]
- Wang, H.; Wen, Y.; Du, Y.; Yan, X.; Guo, H.; Rycroft, J.A.; Boon, N.; Kovacs, E.M.; Mela, D.J. Effects of catechin enriched green tea on body composition. Obes. (Silver Spring) 2010, 18, 773–779. [Google Scholar] [CrossRef]
- Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers; US Department of Health and Human Services, Food and Drug Administration Center for Drug Evaluation and Research: Rockville, MD, USA, 2005.
- Kuriyama, S.; Shimazu, T.; Ohmori, K.; Kikuchi, N.; Nakaya, N.; Nishino, Y.; Tsubono, Y.; Tsuji, I. Green tea consumption and mortality due to cardiovascular disease, cancer, and all causes in Japan: The Ohsaki study. JAMA 2006, 296, 1255–1265. [Google Scholar] [CrossRef]
- Food Labeling: Serving Sizes of Foods That Can Reasonably Be Consumed At One Eating Occasion; Dual-Column Labeling; Updating, Modifying, and Establishing Certain Reference Amounts Customarily Consumed; Serving Size for Breath Mints; and Technical Amendments; Food and Drug Administration, HHS: Rockville, MD, USA, 2016.
- Pan, H.B.; Zhang, D.; Li, B.; Wu, Y.Y.; Tu, Y.Y. A Rapid UPLC Method for Simultaneous Analysis of Caffeine and 13 Index Polyphenols in Black Tea. J. Chromatogr. Sci. 2017, 55, 495–496. [Google Scholar] [CrossRef]
- Kalaivanisailaja, J.; Manju, V.; Nalini, N. Lipid profile in mice fed a high-fat diet after exogenous leptin administration. Pol. J. Pharmacol. 2003, 55, 763–769. [Google Scholar] [PubMed]
- Demori, I.; Voci, A.; Fugassa, E.; Burlando, B. Combined effects of high-fat diet and ethanol induce oxidative stress in rat liver. Alcohol 2006, 40, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Mehra, P.; Garg, M.; Koul, A.; Bansal, D.D. Effect of (+)-catechin hydrate on oxidative stress induced by high sucrose and high fat diet in male Wistar rats. Indian J. Exp. Biol. 2013, 51, 823–827. [Google Scholar] [PubMed]
- Huang, Y.B.; Tsai, M.J.; Wu, P.C.; Tsai, Y.H.; Wu, Y.H.; Fang, J.Y. Elastic liposomes as carriers for oral delivery and the brain distribution of (+)-catechin. J. Drug Target. 2011, 19, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Maliakal, P.; Chen, L.; Meng, X.; Bondoc, F.Y.; Prabhu, S.; Lambert, G.; Mohr, S.; Yang, C.S. Pharmacokinetics of tea catechins after ingestion of green tea and (−)-epigallocatechin-3-gallate by humans: Formation of different metabolites and individual variability. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1025–1032. [Google Scholar]
- Tolstoguzov, V. Thermodynamic aspects of biopolymer functionality in biological systems, foods, and beverages. Crit. Rev. Biotechnol. 2002, 22, 89–174. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xu, Y. Effect of pH on cream particle formation and solids extraction yield of black tea. Food Chem. 2001, 74, 155–160. [Google Scholar] [CrossRef]
- Liang, Y.; Lu, J.; Zhang, L. Comparative study of cream in infusions of black tea and green tea [Camellia sinensis (L.) O. Kuntze]. Int. J. Food Sci. Technol. 2002, 37, 627–634. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Zhu, Q.Y.; Wong, Y.F.; Zhang, Z.; Chung, H.Y. Stabilizing Effect of Ascorbic Acid on Green Tea Catechins. J. Agric. Food Chem. 1998, 46, 2512–2516. [Google Scholar] [CrossRef]
- Cohn, J.S.; McNamara, J.R.; Cohn, S.D.; Ordovas, J.M.; Schaefer, E.J. Postprandial plasma lipoprotein changes in human subjects of different ages. J. Lipid Res. 1988, 29, 469–479. [Google Scholar]
- Nakamura, K.; Miyoshi, T.; Yunoki, K.; Ito, H. Postprandial hyperlipidemia as a potential residual risk factor. J. Cardiol. 2016, 67, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Tiihonen, K.; Rautonen, N.; Alhoniemi, E.; Ahotupa, M.; Stowell, J.; Vasankari, T. Postprandial triglyceride response in normolipidemic, hyperlipidemic and obese subjects—The influence of polydextrose, a non-digestible carbohydrate. Nutr. J. 2015, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Eberspacher, E.; Grull, D.; Kowalczyk, L.; Molnar, T.; Zebeli, Q. Enzymatically Modified Starch Ameliorates Postprandial Serum Triglycerides and Lipid Metabolome in Growing Pigs. PLoS ONE 2015, 10, e0130553. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Ehlers, S.J.; Lee, J.Y.; Fernandez, M.L.; Koo, S.I. Dietary green tea extract lowers plasma and hepatic triglycerides and decreases the expression of sterol regulatory element-binding protein-1c mRNA and its responsive genes in fructose-fed, ovariectomized rats. J. Nutr. 2009, 139, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Petzke, K.J.; Raederstorff, D.; Wolfram, S.; Klaus, S. Acute effects of epigallocatechin gallate from green tea on oxidation and tissue incorporation of dietary lipids in mice fed a high-fat diet. Int. J. Obes. (Lond.) 2012, 36, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Unno, T.; Tago, M.; Suzuki, Y.; Nozawa, A.; Sagesaka, Y.M.; Kakuda, T.; Egawa, K.; Kondo, K. Effect of tea catechins on postprandial plasma lipid responses in human subjects. Br. J. Nutr. 2005, 93, 543–547. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Won, H.; Kim, Y.; Jang, M.; Jyothi, K.R.; Kim, Y.; Dandona, P.; Ha, J.; Kim, S.S. Antioxidant enzymes induced by repeated intake of excess energy in the form of high-fat, high-carbohydrate meals are not sufficient to block oxidative stress in healthy lean individuals. Br. J. Nutr. 2011, 106, 1544–1551. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Zhang, F.; Dai, Y.; Han, L.; Chen, S. Serum TNF-alpha, GTH and MDA of high-fat diet-induced obesity and obesity resistant rats. Saudi Pharm. J. 2016, 24, 333–336. [Google Scholar] [CrossRef]
- Montes-Nieto, R.; Insenser, M.; Murri, M.; Fernandez-Duran, E.; Ojeda-Ojeda, M.; Martinez-Garcia, M.A.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Plasma thiobarbituric acid reactive substances (TBARS) in young adults: Obesity increases fasting levels only in men whereas glucose ingestion, and not protein or lipid intake, increases postprandial concentrations regardless of sex and obesity. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Johnson, B.D.; Padilla, J.; Harris, R.A.; Wallace, J.P. Vascular consequences of a high-fat meal in physically active and inactive adults. Appl. Physiol. Nutr. Metab. 2011, 36, 368–375. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Solis, A.D.; Fisher-Wellman, K.H.; Smith, W.A. Postprandial oxidative stress is exacerbated in cigarette smokers. Br. J. Nutr. 2008, 99, 1055–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloomer, R.J.; Trepanowski, J.F.; Farney, T.M. Influence of acute coffee consumption on postprandial oxidative stress. Nutr. Metab. Insights 2013, 6, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, X.; Chen, J.; Jiao, R.; Wang, L.; Li, Y.M.; Zuo, Y.; Liu, Y.; Lei, L.; Ma, K.Y.; et al. Biology of ageing and role of dietary antioxidants. BioMed Res. Int. 2014, 2014, 831841. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.G.; Farney, T.M.; Canale, R.E.; Dessoulavy, M.E.; Bloomer, R.J. High-fat feeding, but not strenuous exercise, increases blood oxidative stress in trained men. Appl. Physiol. Nutr. Metab. 2013, 38, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Canale, R.E.; Farney, T.M.; McCarthy, C.G.; Bloomer, R.J. Influence of acute exercise of varying intensity and duration on postprandial oxidative stress. Eur. J. Appl. Physiol. 2014, 114, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- Belinova, L.; Kahleova, H.; Malinska, H.; Topolcan, O.; Vrzalova, J.; Oliyarnyk, O.; Kazdova, L.; Hill, M.; Pelikanova, T. Differential acute postprandial effects of processed meat and isocaloric vegan meals on the gastrointestinal hormone response in subjects suffering from type 2 diabetes and healthy controls: A randomized crossover study. PLoS ONE 2014, 9, e107561. [Google Scholar] [CrossRef]
- Yubero-Serrano, E.M.; Delgado-Casado, N.; Delgado-Lista, J.; Perez-Martinez, P.; Tasset-Cuevas, I.; Santos-Gonzalez, M.; Caballero, J.; Garcia-Rios, A.; Marin, C.; Gutierrez-Mariscal, F.M.; et al. Postprandial antioxidant effect of the Mediterranean diet supplemented with coenzyme Q10 in elderly men and women. AGE (Dordr) 2011, 33, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Chan, H.Y.; Huang, Y.; Chen, Z.Y. Green tea catechins upregulate superoxide dismutase and catalase in fruit flies. Mol. Nutr. Food Res. 2007, 51, 546–554. [Google Scholar] [CrossRef]
- Casao, A.; Cebrian, I.; Asumpcao, M.E.; Perez-Pe, R.; Abecia, J.A.; Forcada, F.; Cebrian-Perez, J.A.; Muino-Blanco, T. Seasonal variations of melatonin in ram seminal plasma are correlated to those of testosterone and antioxidant enzymes. Reprod. Biol. Endocrinol. 2010, 8, 59. [Google Scholar] [CrossRef]
- Srivastava, K.C.; Austin, R.D.; Shrivastava, D.; Pranavadhyani, G. Oxidant-antioxidant status in tissue samples of oral leukoplakia. Dent. Res. J. (Isfahan) 2014, 11, 180–186. [Google Scholar]
- Srivastava, K.C.; Austin, R.D.; Shrivastava, D. Evaluation of oxidant-antioxidant status in tissue samples in oral cancer: A case control study. Dent. Res. J. (Isfahan) 2016, 13, 181–187. [Google Scholar] [CrossRef]
- Tinahones, F.J.; Murri-Pierri, M.; Garrido-Sanchez, L.; Garcia-Almeida, J.M.; Garcia-Serrano, S.; Garcia-Arnes, J.; Garcia-Fuentes, E. Oxidative stress in severely obese persons is greater in those with insulin resistance. Obes. (Silver Spring) 2009, 17, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.N.; Morton, H.P.; Cooney, P.T.; Winters, T.G.; Ramseur, K.R.; Rayalam, S.; Della-Fera, M.A.; Baile, C.A.; Brown, L.M. Acute exposure to high-fat diets increases hepatic expression of genes related to cell repair and remodeling in female rats. Nutr. Res. 2014, 34, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Sarma, D.N.; Barrett, M.L.; Chavez, M.L.; Gardiner, P.; Ko, R.; Mahady, G.B.; Marles, R.J.; Pellicore, L.S.; Giancaspro, G.I.; Low Dog, T. Safety of green tea extracts: A systematic review by the US Pharmacopeia. Drug Saf. 2008, 31, 469–484. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hours Post Treatment | LF + GTE (n = 5) | HF + GTE (n = 4) | LF + GTE + LJ (n = 3) | HF + GTE + LJ (n = 4) |
|---|---|---|---|---|
| Pre | 1.00 | 1.00 | 1.00 | 1.00 |
| 1 | 1.01 ± 0.05 | 1.10 ± 0.02 * | 0.92 ± 0.08 | 0.98 ± 0.06 |
| 2 | 0.94 ± 0.04 | 0.98 ± 0.04 | 0.86 ± 0.05 | 0.99 ± 0.05 |
| 3 | 0.97 ± 0.02 | 0.92 ± 0.06 | 0.86 ± 0.06 | 0.99 ± 0.07 |
| 4 | 0.93 ± 0.02 * | 0.95 ± 0.06 | 0.81 ± 0.11 | 0.95 ± 0.02 |
| 6 | 0.92 ± 0.04 | 1.06 ± 0.06 | 0.89 ± 0.11 | 0.77 ± 0.08 * |
| 12 | 0.87 ± 0.07 | 0.78 ± 0.09 | 0.94 ± 0.03 | 1.16 ± 0.07 |
| 24 | 1.02 ± 0.05 | 1.01 ± 0.02 | 0.93 ± 0.04 | 1.11 ± 0.05 |
| Hours Post Treatment | LF + GTE (n = 5) | HF + GTE (n = 4) | LF + GTE + LJ (n = 3) | HF + GTE + LJ (n = 4) |
|---|---|---|---|---|
| Pre | 1.00 | 1.00 | 1.00 | 1.00 |
| 1 | 0.80 ± 0.09 * | 0.99 ± 0.30 | 0.73 ± 0.13 | 0.96 ± 0.10 |
| 2 | 0.99 ± 0.11 | 0.73 ± 0.14 | 0.57 ± 0.07 * | 1.01 ± 0.15 |
| 3 | 1.19 ± 0.12 | 0.74 ± 0.13 | 0.54 ± 0.15 * | 0.85 ± 0.04 * |
| 4 | 0.74 ± 0.22 | 0.60 ± 0.12 * | 0.53 ± 0.17 | 0.68 ± 0.05 * |
| 6 | 0.84 ± 0.07 * | 0.56 ± 0.07 * | 0.47 ± 0.04 * | 0.54 ± 0.17 * |
| 12 | 1.28 ± 0.24 | 1.68 ± 0.10 * | 0.55 ± 0.08 * | 1.42 ± 0.00 |
| 24 | 1.21 ± 0.24 | 0.94 ± 0.26 | 0.59 ± 0.07 | 1.03 ± 0.25 |
| Hours Post Treatment | LF + GTE (n = 5) | HF + GTE (n = 4) | LF + GTE + LJ (n = 3) | HF + GTE + LJ (n = 4) |
|---|---|---|---|---|
| Pre | 1.00 | 1.00 | 1.00 | 1.00 |
| 1 | 1.12 ± 0.16 | 0.79 ± 0.02 * | 1.08 ± 0.13 | 0.60 ± 0.08 * |
| 2 | 1.10 ± 0.16 | 0.96 ± 0.14 | 0.93 ± 0.08 | 0.55 ± 0.15 * |
| 3 | 0.81 ± 0.21 | 0.84 ± 0.05 * | 1.09 ± 0.18 | 0.80 ± 0.15 |
| 4 | 1.05 ± 0.14 | 0.91 ± 0.11 | 0.79 ± 0.00 | 1.17 ± 0.35 |
| 6 | 1.30 ± 0.22 | 0.85 ± 0.13 | 0.87 ± 0.08 | 0.99 ± 0.23 |
| 12 | 1.08 ± 0.17 | 0.91 ± 0.06 | 0.95 ± 0.19 | 0.89 ± 0.34 |
| 24 | 1.10 ± 0.20 | 0.88 ± 0.18 | 1.47 ± 0.33 | 0.99 ± 0.15 |
| Hours Post Treatment | LF + GTE (n = 5) | HF + GTE (n = 4) | LF + GTE + LJ (n = 3) | HF + GTE + LJ (n = 4) |
|---|---|---|---|---|
| Pre | 1.00 | 1.00 | 1.00 | 1.00 |
| 1 | 1.68 ± 0.18 * | 1.06 ± 0.36 | 1.11 ± 0.29 | 1.15 ± 0.27 |
| 2 | 1.95 ± 0.63 | 1.14 ± 0.29 | 1.11 ± 0.44 | 1.33 ± 0.33 |
| 3 | 1.86 ± 0.46 | 1.39 ± 0.44 | 1.51 ± 0.33 | 1.45 ± 0.49 |
| 4 | 1.97 ± 0.58 | 1.41 ± 0.37 | 1.80 ± 1.07 | 1.30 ± 0.24 |
| 6 | 1.05 ± 0.35 | 1.83 ± 0.29 | 0.56 ± 0.35 | 0.81 ± 0.18 |
| 12 | 2.25 ± 0.51 * | 1.32 ± 0.48 | 1.92 ± 0.43 | 0.89 ± 0.18 |
| 24 | 2.39 ± 0.52 * | 1.32 ± 0.34 | 1.93 ± 0.61 | 1.28 ± 0.06 |
| Hours Post Treatment | LF + GTE (n = 5) | HF + GTE (n = 4) | LF + GTE + LJ (n = 3) | HF + GTE + LJ (n = 3) |
|---|---|---|---|---|
| Pre | 1.00 | 1.00 | 1.00 | 1.00 |
| 1 | 1.08 ± 0.11 | 1.05 ± 0.13 | 1.23 ± 0.21 | 0.93 ± 0.06 |
| 2 | 0.99 ± 0.17 | 1.06 ± 0.20 | 1.43 ± 0.69 | 0.96 ± 0.19 |
| 3 | 0.75 ± 0.17 | 0.86 ± 0.08 | 0.74 ± 0.28 | 0.84 ± 0.09 |
| 4 | 1.24 ± 0.13 | 1.54 ± 0.53 | 1.11 ± 0.16 | 1.15 ± 0.20 |
| 6 | 1.07 ± 0.21 | 1.21 ± 0.16 | 1.56 ± 0.47 | 1.34 ± 0.24 |
| 12 | 1.36 ± 0.27 | 0.98 ± 0.03 | 1.40 ± 0.19 | 1.30 ± 0.07 * |
| 24 | 1.25 ± 0.29 | 1.11 ± 0.08 | 1.40 ± 0.40 | 1.23 ± 0.14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, X.; Azain, M.; Crowe-White, K.; Mumaw, J.; Grimes, J.A.; Schmiedt, C.; Barletta, M.; Rayalam, S.; Park, H.J. Effect of Acute Ingestion of Green Tea Extract and Lemon Juice on Oxidative Stress and Lipid Profile in Pigs Fed a High-Fat Diet. Antioxidants 2019, 8, 195. https://doi.org/10.3390/antiox8060195
Fang X, Azain M, Crowe-White K, Mumaw J, Grimes JA, Schmiedt C, Barletta M, Rayalam S, Park HJ. Effect of Acute Ingestion of Green Tea Extract and Lemon Juice on Oxidative Stress and Lipid Profile in Pigs Fed a High-Fat Diet. Antioxidants. 2019; 8(6):195. https://doi.org/10.3390/antiox8060195
Chicago/Turabian StyleFang, Xi, Michael Azain, Kristi Crowe-White, Jennifer Mumaw, Janet A. Grimes, Chad Schmiedt, Michele Barletta, Srujana Rayalam, and Hea Jin Park. 2019. "Effect of Acute Ingestion of Green Tea Extract and Lemon Juice on Oxidative Stress and Lipid Profile in Pigs Fed a High-Fat Diet" Antioxidants 8, no. 6: 195. https://doi.org/10.3390/antiox8060195