Effects of Selenium and Cadmium on Breast Muscle Fatty-Acid Composition and Gene Expression of Liver Antioxidant Proteins in Broilers

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Procedures
2.2. Determination of Fatty-Acid Profile
2.3. RNA Isolation and Reverse Transcription
2.4. Real-Time Quantitative PCR
2.5. Statistical Analysis
3. Results
3.1. Breast Muscle Fatty-Acid Composition
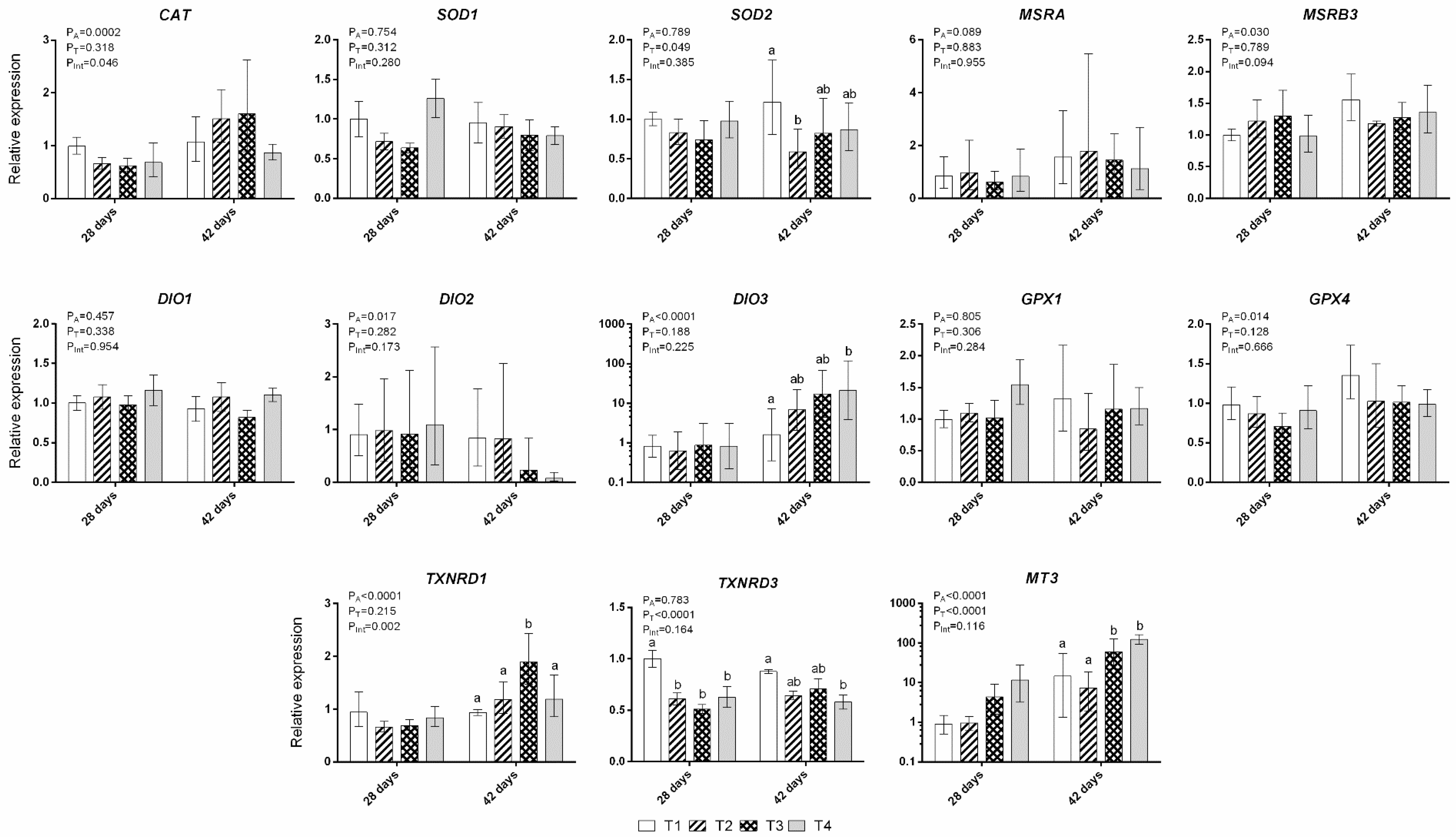
3.2. Gene Expression of Liver Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Al-Waeli, A.; Zoidis, E.; Pappas, A.; Demiris, N.; Zervas, G.; Fegeros, K. The role of organic selenium in cadmium toxicity: Effects on broiler performance and health status. Animal 2013, 7, 386–393. [Google Scholar] [CrossRef]
- Xu, F.; Liu, S.; Li, S. Effects of selenium and cadmium on changes in the gene expression of immune cytokines in chicken splenic lymphocytes. Biol. Trace Elem. Res. 2015, 165, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Shukla, G.S.; Chandra, S.V. Effects of cadmium on superoxide dismutase and lipid peroxidation in liver and kidney of growing rats: In vivo and in vitro studies. Pharmacol. Toxicol. 1987, 60, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, Z.A.; Vu, T.T.; Zaman, K. Oxidative stress as a mechanism of chronic cadmium-induced hepatotoxicity and renal toxicity and protection by antioxidants. Toxicol. Appl. Pharmacol. 1999, 154, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Waisberg, M.; Joseph, P.; Hale, B.; Beyersmann, D. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology 2003, 192, 95–117. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, W.; Chen, X.; Zhu, Y.; Zhang, Z.; Yao, H.; Xu, S. Four endoplasmic reticulum resident selenoproteins may be related to the protection of selenium against cadmium toxicity in chicken lymphocytes. Biol. Trace Elem. Res. 2014, 161, 328–333. [Google Scholar] [CrossRef]
- Li, J.L.; Jiang, C.Y.; Li, S.; Xu, S.W. Cadmium induced hepatotoxicity in chickens (Gallus domesticus) and ameliorative effect by selenium. Ecotoxicol. Environ. Saf. 2013, 96, 103–109. [Google Scholar] [CrossRef]
- Li, J.L.; Gao, R.; Li, S.; Wang, J.T.; Tang, Z.X.; Xu, S.W. Testicular toxicity induced by dietary cadmium in cocks and ameliorative effect by selenium. Biometals 2010, 23, 695–705. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hossain, K.F.B.; Banik, S.; Sikder, M.T.; Akter, M.; Bondad, S.E.C.; Rahaman, M.S.; Hosokawa, T.; Saito, T.; Kurasaki, M. Selenium and zinc protections against metal-(loids)-induced toxicity and disease manifestations: A review. Ecotoxicol. Environ. Saf. 2019, 168, 146–163. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Zoidis, E.; Surai, P.F.; Zervas, G. Selenoproteins and maternal nutrition. Comp. Biochem. Physiol. 2008, 151, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Zoidis, E.; Pappas, A.C.; Georgiou, C.A.; Komaitis, E.; Fegeros, K. Selenium affects the expression of GPx4 and catalase in the liver of chicken. Comp. Biochem. Physiol. 2010, 155, 294–300. [Google Scholar] [CrossRef]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G. Selenium-dependent antioxidant enzymes: Actions and properties of selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, I.; Banni, M.; Said, L.; Said, K.; Kerkeni, A. Involvement of selenoprotein P and GPx4 gene expressions in cadmium-induced testicular pathophysiology in rat. Chem. Biol. Interact. 2010, 188, 94–101. [Google Scholar] [CrossRef]
- Pappas, A.C.; Zoidis, E.; Fegeros, K.; Zervas, G.; Surai, P.F. Cadmium Toxicity and the Antioxidant System. In Environmental Health-Physical, Chemical and Biological Factors Series, 1st ed.; Pappas, A.C., Zoidis, E., Fegeros, K., Zervas, G., Surai, P.F., Eds.; Nova Science Publishers: New York, NY, USA, 2010; pp. 1–63. [Google Scholar]
- Jihen, E.H.; Imed, M.; Fatima, H.; Abdelhamid, K. Protective effects of selenium (Se) and zinc (Zn) on cadmium (Cd) toxicity in the liver and kidney of the rat: histology and Cd accumulation. Food Chem. Toxicol. 2008, 46, 3522–3527. [Google Scholar] [CrossRef]
- Lazarus, M.; Orct, T.; Jurasoviae, J.; Blanuša, M. The effect of dietary selenium supplementation on cadmium absorption and retention in suckling rats. Biometals 2009, 22, 973–983. [Google Scholar] [CrossRef]
- Mahan, D.C. Organic selenium: Using nature’s model to redefine selenium supplementation for animals. In Biotechnology in the Feed Industry, 1st ed.; Lyons, T.P., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 1999. [Google Scholar]
- Mahmoud, K.Z.; Edens, F.W. Influence of selenium sources on age related and mild heat stress-related changes of blood and liver glutathione redox cycle in broiler chickens (Gallus domesticus). Comp. Biochem. Physiol. 2003, 136, 921–934. [Google Scholar] [CrossRef]
- Al-Waeli, A.; Pappas, A.C.; Zoidis, E.; Georgiou, C.A.; Zervas, G.; Fegeros, K. The role of selenium in cadmium toxicity: Interactions with essential and toxic elements. Br. Poult. Sci. 2012, 53, 817–827. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, Y.H.; Cheng, X.Y.; Zhang, Z.W.; Xu, S.W. The protection of selenium against cadmium-induced cytotoxicity via the heat shock protein pathway in chicken splenic lymphocytes. Molecules 2012, 17, 14565–14572. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, C.M.; Zhang, Z.W.; Yao, H.D.; Xu, S.W. Protective effects of selenium on cadmium-induced brain damage in chickens. Biol. Trace Elem. Res. 2014, 158, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Zoidis, E.; Papadomichelakis, G.; Fegeros, K. Supranutritional selenium level affects fatty acid composition and oxidative stability of chicken breast muscle tissue. J. Anim. Physiol. Anim. Nutr. 2012, 96, 385–394. [Google Scholar] [CrossRef] [PubMed]
- NRC (National Research Council). Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Marshall, O.J. PerlPrimer: Cross-platform, graphical primer design for standard, bisulphite and real-time PCR. Bioinformatics 2004, 20, 2471–2472. [Google Scholar] [CrossRef]
- Iqbal, M.; Philbin, V.J.; Withanage, G.S.K.; Wigley, P.; Beal, R.K.; Goodchild, M.J.; Barrow, P.; McConnell, I.; Maskell, D.J.; Young, J.; et al. Identification and functional characterization of chicken Toll-like receptor 5 reveals a fundamental role in the biology of infection with Salmonella enterica serovar Typhimurium. Infect. Immun. 2005, 73, 2344–2350. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- EC 2005. Commission Directive 05/87/EC Amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council on undesirable substances in animal feed as regards lead, fluorine and cadmium. Off. J. Eur. Union 2002, L318, 19–24. [Google Scholar]
- Li, Y.X.; Xiong, X.; Lin, C.Y.; Zhang, F.S.; Li, W.; Han, W. Cadmium in animal production and its potential hazard on Beijing and Fuxin farmlands. J. Hazard. Mater. 2010, 177, 475–480. [Google Scholar] [CrossRef]
- Nolan, T.D.; Brown, D. The influence of elevated dietary zinc, selenium, and their combination on the suppressive effect of dietary and intraperitoneal cadmium on egg production in laying hens. J. Toxicol. Environ. Health 2000, 60, 549–565. [Google Scholar]
- Jemai, H.; Messaoudi, I.; Chaouch, A.; Kerkeni, A. Protective effect of zinc supplementation on blood antioxidant defense system in rats exposed to cadmium. J. Trace Elem. Med. Biol. 2007, 21, 269–273. [Google Scholar] [CrossRef]
- Food and Drug Administration. CFR–Code of Federal Regulations Title 21. 2018. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/cfrsearch.cfm?fr=573.920 (accessed on 8 April 2019).
- Gad, M.A.; Abd El-Twab, S.M. Selenium toxicosis assessment (in vivo and in vitro) and the protective role of vitamin B12 in male quail (Coturnix coturnix). Environ. Toxicol. Pharmacol. 2009, 27, 7–16. [Google Scholar] [CrossRef]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: how does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banni, M.; Messaoudi, I.; Saïd, L.; El Heni, J.; Kerkeni, A.; Saïd, K. Metallothionein gene expression in liver of rats exposed to cadmium and supplemented with zinc and selenium. Arch. Environ. Contam. Toxicol. 2010, 59, 513–519. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein protection of cadmium toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.M.; Cherian, M.G. Protective roles of metallothionein and glutathione in hepatotoxicity of cadmium. Toxicology 1992, 72, 281–290. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Liu, J. Metallothionein transgenic and knock-out mouse models in the study of cadmium toxicity. J. Toxicol. Sci. 1998, 23, 97–102. [Google Scholar] [CrossRef]
- Park, J.D.; Liu, Y.; Klaassen, C.D. Protective effect of metallothionein against the toxicity of cadmium and other metals. Toxicology 2001, 163, 93–100. [Google Scholar] [CrossRef]
- Xie, L.; Klerks, P.L. Metallothionein-like protein in the least killifish Heterandria Formosa and its role in cadmium resistance. Environ. Toxicol. Chem. 2004, 23, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Underwood, E.J.; Suttle, N.F. The Mineral Nutrition of Livestock, 3rd ed.; Underwood, E.J., Suttle, N.F., Eds.; CAB International: Wallingford, UK, 1999. [Google Scholar]
- Wangher, P.D. Selenium and the brain: a review. Nutr. Neurosci. 2001, 4, 81–97. [Google Scholar] [CrossRef]
- Choi, H.J.; Ji, J.; Chung, K.H.; Ahn, I.Y. Cadmium bioaccumulation and detoxification in the gill and digestive gland of the Antarctic bivalve Laternula elliptica. Comp. Biochem. Physiol. 2007, 145, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, M.; Andres, S.; Durrieu, G.; Boudou, A. The key role of metallothioneins in the bivalve Corbicula fluminea during the depuration phase, after in situ exposure to Cd and Zn. Aquat. Toxicol. 2003, 63, 89–102. [Google Scholar] [CrossRef]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar] [PubMed]
- Liang, Y.; Lin, S.L.; Wang, C.W.; Yao, H.D.; Zhang, Z.W.; Xu, S.W. Effect of selenium on selenoprotein expression in the adipose tissue of chickens. Biol. Trace Elem. Res. 2014, 160, 41–48. [Google Scholar] [CrossRef] [PubMed]
- El-Sharaky, A.S.; Newairy, A.A.; Badreldeen, M.M.; Eweda, S.M.; Sheweita, S.A. Protective role of selenium against renal toxicity induced by cadmium in rats. Toxicology 2007, 235, 185–193. [Google Scholar] [CrossRef]
- Surai, P.F. Selenium in Nutrition and Health; Nottingham University Press: Nottingham, UK, 2006. [Google Scholar]
- Liu, L.; Yang, B.; Cheng, Y.; Lin, H. Ameliorative effects of selenium on cadmium-induced oxidative stress and endoplasmic reticulum stress in the chicken kidney. Biol. Trace Elem. Res. 2015, 167, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Lammi, M.; Qu, C. Selenium-Related transcriptional regulation of gene expression. Int. J. Mol. Sci. 2018, 19, 2665. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Hu, X.; Smith, M.R.; Go, Y.M.; Jones, D.P. Selenium at the redox interface of the genome, metabolome and exposome. Free Radic. Biol. Med. 2018, 127, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Casalino, E.; Calzaretti, G.; Landriscina, M.; Sblano, C.; Fabiano, A.; Landriscina, C. The Nrf2 transcription factor contributes to the induction of alpha-class GST isoenzymes in liver of acute cadmium or manganese intoxicated rats: Comparison with the toxic effect on NAD(P)H:quinone reductase. Toxicology 2007, 237, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Casalino, E.; Calzaretti, G.; Sblano, C.; Landriscina, C. Cadmium-dependent enzyme activity alteration is not imputable to lipid peroxidation. Archiv. Biochem. Biophys. 2000, 383, 288–295. [Google Scholar] [CrossRef]
- McCarty, M.F. Zinc and multi-mineral supplementation should mitigate the pathogenic impact of cadmium exposure. Med. Hypotheses 2012, 79, 642–648. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Wang, C.; Zhao, P.; Liu, H.; Li, J.; Bao, J. Ameliorative effects of dietary selenium against cadmium toxicity is related to changes in trace elements in chicken kidneys. Biol. Trace Elem. Res. 2017, 176, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Branca, J.J.V.; Morucci, G.; Maresca, M.; Tenci, B.; Cascella, R.; Paternostro, F.; Ghelardini, C.; Gulisano, M.; Di Cesare Mannelli, L.; Pacini, A. Selenium and zinc: Two key players against cadmium-induced neuronal toxicity. Toxicol. In Vitro 2018, 48, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Boshy, M.E.; Risha, E.F.; Abdelhamid, F.M.; Mubarak, M.S.; Hadda, T.B. Protective effects of selenium against cadmium induced hematological disturbances, immunosuppressive, oxidative stress and hepatorenal damage in rats. J. Trace Elem. Med. Biol. 2015, 29, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]


{kind=link}
| Ingredients (g∙kg−1) | Starter (0–10 days) | Grower (11–20 days) | Finisher (21–42 days) |
|---|---|---|---|
| Maize | 559.5 | 631.0 | 658.4 |
| Soybean meal | 333.7 | 261.0 | 226.4 |
| Gluten | 50.0 | 50.0 | 50.0 |
| Soybean oil | 15.4 | 16.9 | 25.7 |
| Dicalcium phosphate | 15.1 | 14.4 | 13.2 |
| Limestone | 16.1 | 15.7 | 14.8 |
| Lysine | 0.2 | 1.5 | 2.1 |
| Methionine | 1.1 | 1.3 | 1.5 |
| NaCl | 4.9 | 4.2 | 3.9 |
| Premix1 | 4.0 | 4.0 | 4.0 |
| Calculated Analysis | |||
| Metabolizable energy (ME) (MJ∙kg−1) | 12.5 | 12.9 | 13.3 |
| Crude protein (CP) (g∙kg−1) | 210.0 | 190.0 | 180.0 |
| Sodium (g∙kg−1) | 2.0 | 1.7 | 1.6 |
| Ca (g∙kg−1) | 10.0 | 9.6 | 9.0 |
| Available phosphorus (g∙kg−1) | 5.0 | 4.8 | 4.5 |
| Methionine (g∙kg−1) | 5.3 | 5.1 | 5.2 |
| Methionine + cystine (g∙kg−1) | 8.9 | 8.4 | 8.2 |
| Lysine (g∙kg−1) | 12.0 | 11.0 | 10.5 |
| Fatty-Acid (FA) Profile (% of total FA) | |||
| 14:0 | 0.08 | 0.08 | 0.09 |
| 16:0 | 13.27 | 13.28 | 13.35 |
| cis7-16:1 | 0.04 | 0.05 | 0.04 |
| cis9-16:1 | 0.15 | 0.15 | 0.10 |
| 17:0 | 0.10 | 0.09 | 0.10 |
| 17:1 | 0.05 | 0.05 | 0.05 |
| 18:0 | 3.15 | 3.18 | 3.39 |
| cis9-18:1 | 26.51 | 27.20 | 26.64 |
| 18:2n-6 | 48.70 | 48.39 | 47.90 |
| 18:3n-3 | 4.06 | 3.66 | 4.14 |
| 20:0 | 0.42 | 0.40 | 0.44 |
| 20:1n-9 | 0.33 | 0.30 | 0.30 |
| 20:2n-6 | 0.08 | 0.07 | 0.09 |
| Gene Name (Gallus gallus NCBI Transcript ID) | Primer Name | Sequence 5′ to 3′ | Position in Transcript |
|---|---|---|---|
| CAT (NM_001031215.2) | F-CAT gal | TCCACGTTAAGACCGATCAG | 839–858 |
| R-CAT gal | AATGTAGAAAGACCAGGAAGGA | 960–981 | |
| SOD1 (NM_205064.1) | F-CuZnSOD gal | ATGCAGATAGGCACGTGG | 229–246 |
| R-CuZnSOD gal | ACTGCCATCTTAAGCATTTCAG | 474–495 | |
| SOD2 (NM_204211.1) | F-MnSOD gal | AGCCTAAAGGAGAATTGATGGA | 365–386 |
| R-MnSOD gal | CAGCAATGGAATGAGACCTG | 551–570 | |
| MSRA (XM_004935891.1) | F-MSRA gal | CAAAGAAGTCTGTTCAGGTTTAGG | 359–382 |
| R-MSRA gal | GCATTCCTTGTGTCGGGT | 559–576 | |
| MSRB3 (NM_001199578.1) | F-MSRB3 gal | GACGGAAAGTGCCTTTGAG | 253–271 |
| R-MSRB3 gal | GGACTGATCACATCATAGAAGGA | 377–399 | |
| DIO1 (NM_001097614.1) | F-DIO1 gal | GCTCACGCAGTAGATGGA | 404–421 |
| R-DIO1 gal | CACTCCTCCCTTGTAGATGAC | 605–625 | |
| DIO2 (NM_204114.3) | F-DIO2 gal | TACAAGCAGGTCAAACTTGGA | 806–826 |
| R-DIO2 gal | CACACTTGCCACCAACAC | 897–914 | |
| DIO3 (NM_001122648.1) | F-DIO3(2) gal | CATGTTGATTTGCCAGCCA | 942–960 |
| R-DIO3(2) gal | AACAGATCCCGAAGGAAGAG | 1078–1097 | |
| GPX1 (NM_001277853.1) | F-GPX1 gal | AACCAATTCGGGCACCAG | 254–271 |
| R-GPX1 gal | CCAGATGATGTACTGCGGG | 457–475 | |
| GPX4 (AF498316.2) | F-GPX4 gal | TGTGGAAGTGGCTGAAGG | 378–395 |
| R-GPX4 gal | CTCGATCACGTAAGGATCCTC | 500–520 | |
| TXNRD1 (NM_001030762.2) | F-TXNRD1 gal | AGGAATACAATACTGTGCTGCT | 1014–1035 |
| R-TXNRD1 gal | TGAACTAACAGTCTTCCTGCC | 1213–1233 | |
| TXNRD3 (NM_001122777.1) | F-TXNRD3 gal | TGAGAAGAATGGGAAAGTACCTG | 1344–1366 |
| R-TXNRD3 gal | CGTTGATATAGTCACACTTTGTGG | 1499–1522 | |
| MT3 (NM_001097538.1) | F-MT3 gal | ACGTGTGGAGACAACTGC | 42–59 |
| R-MT3 gal | GCACACTTGGCACATCCT | 119–136 | |
| GAPDH (NM_204305.1) | F-GAPDH gal | GCTGAATGGGAAGCTTACTG | 720–739 |
| R-GAPDH gal | AAGGTGGAGGAATGGCTG | 918–935 | |
| ACTB (NM_205518.1) | F-ACTB gal | TGCTGTGTTCCCATCTATCG | 152–171 |
| R-ACTB gal | TTGGTGACAATACCGTGTTCA | 281–301 | |
| HPRT1 (NM_204848.1) | F-HPRT1 gal | CGGCCAGACTTTGTTGGA | 569–586 |
| R-HPRT1 gal | ACCAGAGTTGAAGCCTGTG | 768–786 |
| Treatment (T) 1 | RMSE 2 | Day of Age (D) | RMSE 2 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | 28 | 42 | T | D | T × D | |||
| Total weights of FA | 996.33 a | 910.14 a | 609.93 b | 597.72 b | 92.882 | 799.38 | 757.68 | 65.677 | <0.001 | 0.528 | 0.171 |
| 14:0 | 0.38 | 0.39 | 0.45 | 0.44 | 0.025 | 0.43 | 0.40 | 0.018 | 0.067 | 0.038 | 0.002 |
| 14:1 | 0.09 a | 0.10 ab | 0.12 b | 0.11 ab | 0.009 | 0.11 | 0.09 | 0.006 | 0.067 | 0.001 | 0.111 |
| 15:0 | 0.08 | 0.07 | 0.06 | 0.07 | 0.006 | 0.07 | 0.07 | 0.004 | 0.191 | 0.822 | 0.342 |
| 16:0 | 20.55 | 20.26 | 21.15 | 19.96 | 0.521 | 20.51 | 20.45 | 0.367 | 0.135 | 0.865 | 0.622 |
| cis7-16:1 | 0.37 a | 0.37 a | 0.40 ab | 0.46 b | 0.031 | 0.47 | 0.33 | 0.022 | 0.022 | <0.001 | 0.041 |
| cis9-16:1 | 2.98 a | 3.35 ab | 4.04 b | 3.89 b | 0.298 | 4.15 | 2.98 | 0.210 | 0.012 | <0.001 | 0.312 |
| 17:0 | 0.10 | 0.09 | 0.09 | 0.09 | 0.006 | 0.08 | 0.11 | 0.004 | 0.452 | <0.001 | 0.581 |
| 17:1 | 0.20 ab | 0.19 a | 0.25 b | 0.24 ab | 0.018 | 0.22 | 0.23 | 0.013 | 0.011 | 0.278 | 0.692 |
| 18:0 | 9.28 | 8.65 | 9.05 | 9.14 | 0.276 | 8.68 | 9.38 | 0.194 | 0.129 | 0.001 | 0.663 |
| cis9-18:1 | 23.00 | 22.96 | 23.26 | 24.71 | 0.948 | 24.77 | 22.19 | 0.668 | 0.273 | <0.001 | 0.468 |
| cis11-18:1 | 3.21 a | 3.22 a | 3.47 ab | 3.71 a | 0.160 | 3.63 | 3.18 | 0.113 | 0.025 | <0.001 | 0.673 |
| 18:2n-6 | 18.64 a | 18.95 a | 16.49 b | 16.05 b | 0.620 | 17.34 | 17.73 | 0.437 | <0.001 | 0.375 | 0.585 |
| 18:3n-6 | 0.11 | 0.14 | 0.13 | 0.13 | 0.011 | 0.13 | 0.13 | 0.007 | 0.152 | 0.730 | 0.776 |
| 18:3n-3 | 0.74 ab | 0.79 a | 0.63 b | 0.73 ab | 0.039 | 0.78 | 0.67 | 0.028 | 0.004 | <0.001 | 0.013 |
| 20:0 | 0.05 | 0.05 | 0.15 | 0.07 | 0.071 | 0.05 | 0.11 | 0.050 | 0.477 | 0.312 | 0.506 |
| 20:1n-9 | 0.49 | 0.45 | 0.41 | 0.47 | 0.043 | 0.55 | 0.36 | 0.030 | 0.337 | <0.001 | 0.634 |
| 20:2n-6 | 0.82 a | 0.85 a | 0.54 b | 0.50 b | 0.045 | 0.72 | 0.64 | 0.032 | <0.001 | 0.010 | 0.077 |
| 20:3n-6 | 1.30 | 1.35 | 1.46 | 1.46 | 0.074 | 1.42 | 1.37 | 0.052 | 0.178 | 0.353 | 0.118 |
| 20:4n-6 | 4.30 | 4.35 | 3.69 | 3.67 | 0.403 | 3.30 | 4.70 | 0.284 | 0.299 | <0.001 | 0.967 |
| 20:5n-3 | 0.29 a | 0.31 ab | 0.40 bc | 0.44 c | 0.031 | 0.33 | 0.39 | 0.022 | <0.001 | 0.015 | 0.325 |
| 22:2n-6 | 1.84 | 1.75 | 1.66 | 1.65 | 0.109 | 1.80 | 1.66 | 0.077 | 0.438 | 0.074 | 0.908 |
| 22:4n-6 | 1.09 a | 1.13 a | 0.73 b | 0.71 b | 0.069 | 0.78 | 1.05 | 0.049 | <0.001 | <0.001 | 0.256 |
| 22:5n-3 | 0.64 | 0.64 | 0.58 | 0.63 | 0.058 | 0.50 | 0.74 | 0.041 | 0.714 | <0.001 | 0.807 |
| 22:6n-3 | 0.51 | 0.53 | 0.52 | 0.59 | 0.047 | 0.50 | 0.57 | 0.033 | 0.451 | 0.035 | 0.723 |
| SFA3 | 30.50 | 29.56 | 31.00 | 29.86 | 0.570 | 29.89 | 30.57 | 0.402 | 0.063 | 0.093 | 0.853 |
| MUFA3 | 30.29 | 30.58 | 31.86 | 33.54 | 1.294 | 33.81 | 29.32 | 0.912 | 0.119 | <0.001 | 0.391 |
| PUFA3 | 30.28 a | 30.77 a | 26.83 b | 26.55 b | 1.020 | 27.59 | 29.63 | 0.719 | <0.001 | 0.006 | 0.823 |
| PUFA:SFA | 1.00 ac | 1.04 a | 0.87 b | 0.89 bc | 0.042 | 0.93 | 0.97 | 0.030 | 0.001 | 0.141 | 0.697 |
| n-34 | 2.19 | 2.26 | 2.14 | 2.38 | 0.115 | 2.11 | 2.37 | 0.081 | 0.183 | 0.002 | 0.606 |
| n-64 | 22.98 a | 23.31 a | 20.47 b | 20.00 b | 0.639 | 21.45 | 21.92 | 0.450 | <0.001 | 0.300 | 0.533 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoidis, E.; Papadomichelakis, G.; Pappas, A.C.; Theodorou, G.; Fegeros, K. Effects of Selenium and Cadmium on Breast Muscle Fatty-Acid Composition and Gene Expression of Liver Antioxidant Proteins in Broilers. Antioxidants 2019, 8, 147. https://doi.org/10.3390/antiox8050147
Zoidis E, Papadomichelakis G, Pappas AC, Theodorou G, Fegeros K. Effects of Selenium and Cadmium on Breast Muscle Fatty-Acid Composition and Gene Expression of Liver Antioxidant Proteins in Broilers. Antioxidants. 2019; 8(5):147. https://doi.org/10.3390/antiox8050147
Chicago/Turabian StyleZoidis, Evangelos, George Papadomichelakis, Athanasios C. Pappas, Georgios Theodorou, and Kostas Fegeros. 2019. "Effects of Selenium and Cadmium on Breast Muscle Fatty-Acid Composition and Gene Expression of Liver Antioxidant Proteins in Broilers" Antioxidants 8, no. 5: 147. https://doi.org/10.3390/antiox8050147