
Modulation of Radiation Response by the Tetrahydrobiopterin Pathway

 ,
, 
Abstract
:1. Introduction
2. Cellular Redox Responses Following Radiation Exposure
3. Radiation Effect on BH4 and Peroxynitrite Formation
4. Radiation Effect on GFRP and the Molecular Signaling Pathways that Modulates BH4 Biosynthesis
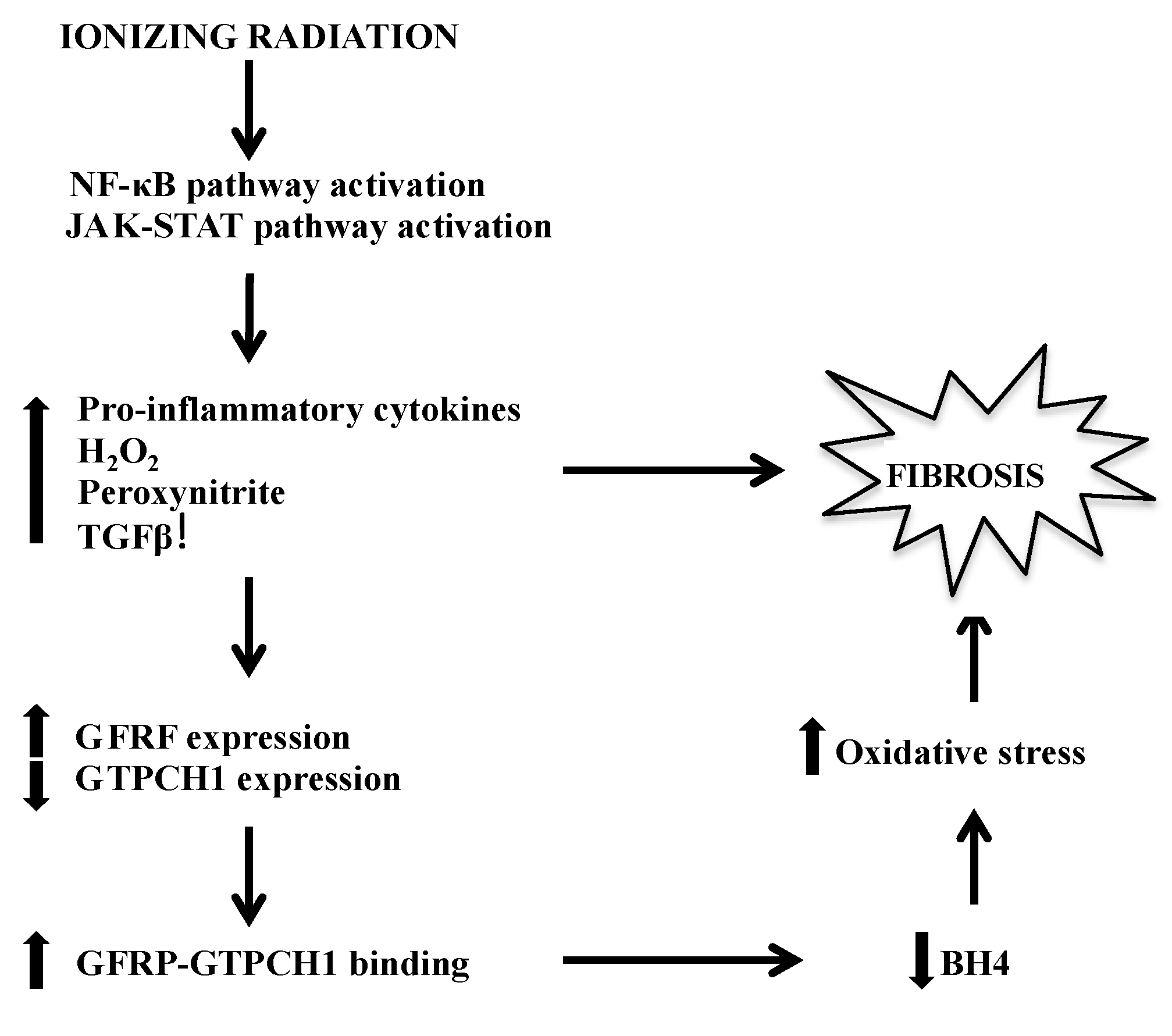
5. Possible Mechanisms of IR-Induced BH4 Deficiency-Mediated Fibrosis Development
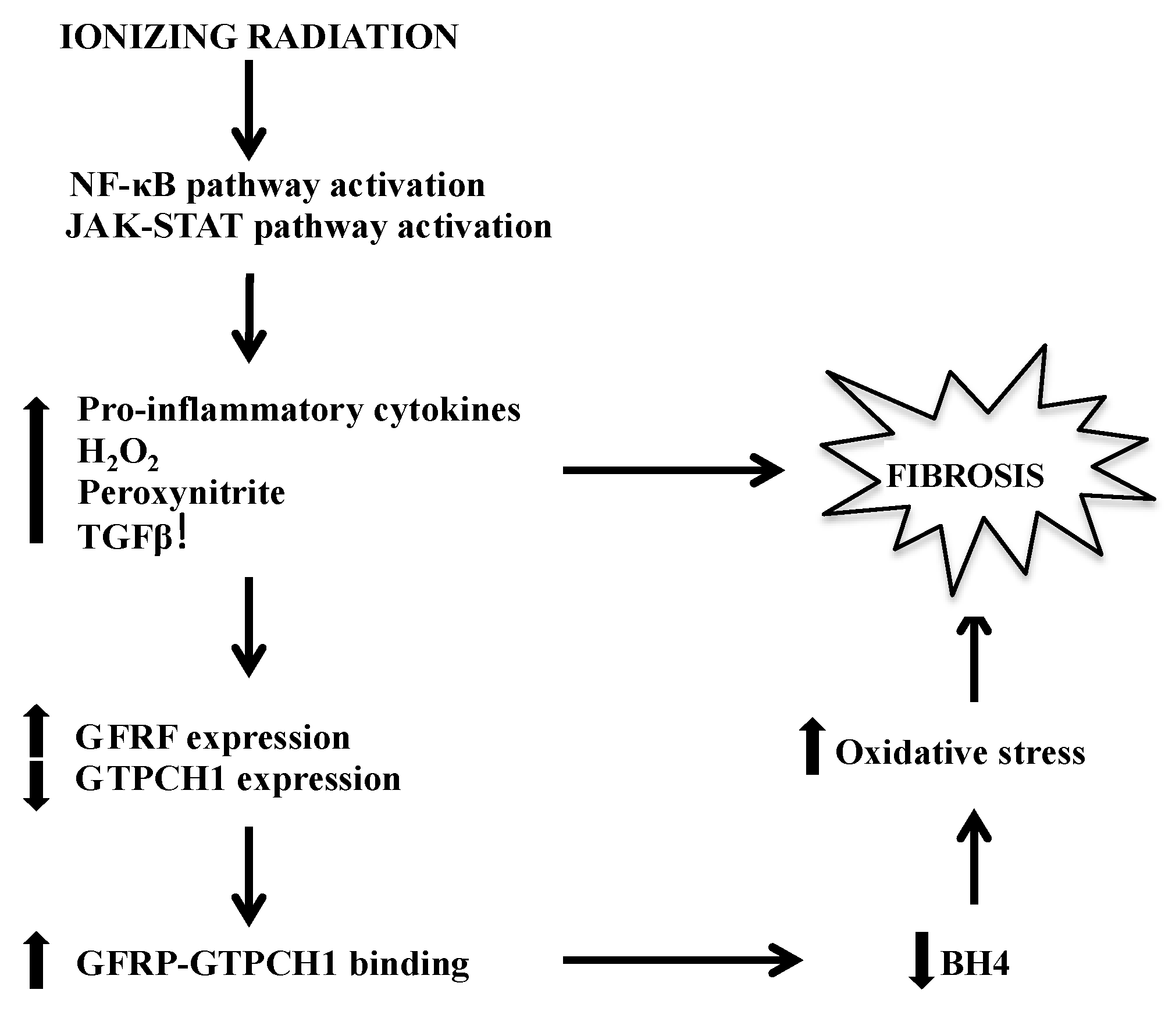
6. Metabolomics Enhances Our View of BH4’s Role in Radiation Response
7. Conclusions

{kind=link}
| Possible Biomarkers | Method Used | Animal Models | Specimen | Year Reference |
|---|---|---|---|---|
| Downregulated in WT mice | UPLC-ESI-QTOFMS, SID-MRM-MS | Mice | Liver tissue | 2014 [51] |
| Bilirubin, AMD, cytidine, glycerophospholipids, 6-acetyl-d-glucose, C16 sphingomyelin | ||||
| Upregulated in WT mice | ||||
| Secondary bile acids, PS (22:1/0:0) | ||||
| Downregulated in Gfrp mice | ||||
| Glutathione, carnitines, lysoPCs | ||||
| Upregulated in Gfrp transgenic mice | ||||
| Glycocholic acid, N-arachidonoyl taurine, flavin adenine dinucleotide, bile acids | ||||
| Upregulated | GC-TOFMS | Rats | Serum | 2012 [76] |
| Inositol, serine, lysine, glycine, threonine, glycerol | ||||
| Downregulated | ||||
| Isocitrate, gluconic acid, stearic acid | ||||
| Upregulated | UPLC-ESI-QTOFMS | Rhesus monkeys | Urine | 2012 [77] |
| N-acetyltaurine, isethionic acid, taurine, xanthine, hypoxanthine, uric acid, creatine, creatinine, tyrosol sulfate, 3-hydroxytyrosol sulfate, tyramine sulfate, N-acetylserotonin sulfate, adipic acid | ||||
| Upregulated | 1H NMR | Mice | Serum | 2011 [78] |
| Lactate, amino acids, choline, lipid signals | ||||
| Downregulated | ||||
| Glucose signals | ||||
| Upregulated | UPLC-ESI-QTOFMS | Rats | Urine | 2011 [79] |
| thymidine, 2′-deoxyuridine, 2′-deoxyxanthosine, N1-acetylspermidine, N-acetylglucosamine/galactosamine-6-sulfate, N-acetyltaurine, N-hexanoylglycine, taurine, isethionic acid (tentative) | ||||
| Upregulated | GC-MS | Rats | Urine | 2009 [80] |
| Glyoxylate, threonate, thymine, uracil, p-cresol | ||||
| Downregulated | ||||
| Citrate, 2-oxoglutarate, adipate, pimelate, suberate, azelaate | ||||
| Upregulated | UPLC-QTOFMS | Mice | Urine | 2009 [81] |
| thymidine, 2′-deoxyuridine, 2′-deoxyxanthosine, xanthine, xanthosine | ||||
| Upregulated | UPLC–TOFMS | Mice | Urine | 2008 [19] |
| N-hexanoylglycine, β-thymidine, 3-hydroxy-2-methylbenzoic acid 3-O-sulfate, taurine |
Acknowledgments
Conflicts of Interest
References
- Rana, S.; Kumar, R.; Sultana, S.; Sharma, R.K. Radiation-induced biomarkers for the detection and assessment of absorbed radiation doses. J. Pharm. Bioallied Sci. 2010, 2, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Hauer-Jensen, M.; Kumar, K.S.; Wang, J.; Berbee, M.; Fu, Q. Intestinal toxicity in radiation and combined injury: Significance, mechanisms, and countermeasures. In Global Terrorism Issues and Developments; Larche, R.A., Ed.; Nova Science Publishers: New York, NY, USA, 2007; pp. 61–100. [Google Scholar]
- Citrin, D.; Cotrim, A.P.; Hyodo, F.; Baum, B.J.; Krishna, M.C.; Mitchell, J.B. Radioprotectors and mitigators of radiation-induced normal tissue injury. Oncologist 2010, 15, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Hafer, N.; Cassatt, D.; Dicarlo, A.; Ramakrishnan, N.; Kaminski, J.; Norman, M.K.; Maidment, B.; Hatchett, R. NIAID/NIH radiation/nuclear medical countermeasures product research and development program. Health Phys. 2010, 98, 903–905. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.B.; Moyer, B.R.; Prasher, J.; Cliffer, K.D.; Ramakrishnan, N.; Kaminski, J.; Coleman, C.N.; Manning, R.G.; Maidment, B.W.; Hatchett, R. Rapid radiation dose assessment for radiological public health emergencies: Roles of niaid and barda. Health Phys. 2010, 98, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, A.L.; Ramakrishnan, N.; Hatchett, R.J. Radiation combined injury: Overview of niaid research. Health Phys. 2010, 98, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Rothman, N.; Stewart, W.F.; Schulte, P.A. Incorporating biomarkers into cancer epidemiology: A matrix of biomarker and study design categories. Cancer Epidemiol. Biomark. Prev. 1995, 4, 301–311. [Google Scholar]
- Horan, J.R.; Gammill, W.P. The health physics aspects of the SL-1 accident. Health Phys. 1963, 9, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Gonzalez, F.J. Challenges and opportunities of metabolomics. J. Cell. Physiol. 2012, 227, 2975–2981. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.R.; Cohen, E.P. Chronic oxidative stress after irradiation: An unproven hypothesis. Med. Hypotheses 2013, 80, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Azzam, E.I.; Li, J.J.; Gius, D. Metabolic oxidation/reduction reactions and cellular responses to ionizing radiation: A unifying concept in stress response biology. Cancer Metastasis Rev. 2004, 23, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Coy, S.L.; Cheema, A.K.; Tyburski, J.B.; Laiakis, E.C.; Collins, S.P.; Fornace, A., Jr. Radiation metabolomics and its potential in biodosimetry. Int. J. Radiat. Biol. 2011, 87, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.D.; Lanz, C.; Gonzalez, F.J.; Idle, J.R. The role of mass spectrometry-based metabolomics in medical countermeasures against radiation. Mass Spectrom. Rev. 2010, 29, 503–521. [Google Scholar] [PubMed]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Blow, N. Metabolomics: Biochemistry’s new look. Nature 2008, 455, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xiao, J.F.; Tuli, L.; Ressom, H.W. LC-MS-based metabolomics. Mol. Biosyst. 2012, 8, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Varghese, R.S.; Cheema, A.; Cheema, P.; Bourbeau, M.; Tuli, L.; Zhou, B.; Jung, M.; Dritschilo, A.; Ressom, H.W. Analysis of LC-MS data for characterizing the metabolic changes in response to radiation. J. Proteome Res. 2010, 9, 2786–2793. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.D.; Li, H.; Eichler, G.S.; Krausz, K.W.; Weinstein, J.N.; Fornace, A.J., Jr.; Gonzalez, F.J.; Idle, J.R. UPLC-ESI-TOFMS-based metabolomics and gene expression dynamics inspector self-organizing metabolomic maps as tools for understanding the cellular response to ionizing radiation. Anal. Chem. 2008, 80, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Tyburski, J.B.; Patterson, A.D.; Krausz, K.W.; Slavik, J.; Fornace, A.J., Jr.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 1. Identification of minimally invasive urine biomarkers for γ-radiation exposure in mice. Radiat. Res. 2008, 170, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Slane, B.G.; Aykin-Burns, N.; Smith, B.J.; Kalen, A.L.; Goswami, P.C.; Domann, F.E.; Spitz, D.R. Mutation of succinate dehydrogenase subunit C results in increased O2•−, oxidative stress, and genomic instability. Cancer Res. 2006, 66, 7615–7620. [Google Scholar] [CrossRef] [PubMed]
- Aykin-Burns, N.; Slane, B.G.; Liu, A.T.; Owens, K.M.; O’Malley, M.S.; Smith, B.J.; Domann, F.E.; Spitz, D.R. Sensitivity to low-dose/low-let ionizing radiation in mammalian cells harboring mutations in succinate dehydrogenase subunit C is governed by mitochondria-derived reactive oxygen species. Radiat. Res. 2011, 175, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Oberley, L.W.; Lindgren, L.A.; Baker, S.A.; Stevens, R.H. Superoxide lon as the cause of the oxygen effect. Radiat. Res. 1976, 68, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Biaglow, J.E.; Mitchell, J.B.; Held, K. The importance of peroxide and superoxide in the X-ray response. Int. J. Radiat. Oncol. Biol. Phys. 1992, 22, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Petkau, A.; Chelack, W.S.; Pleskach, S.D. Letter: Protection of post-irradiated mice by superoxide dismutase. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1976, 29, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Dayal, D.; Martin, S.M.; Owens, K.M.; Aykin-Burns, N.; Zhu, Y.; Boominathan, A.; Pain, D.; Limoli, C.L.; Goswami, P.C.; Domann, F.E.; et al. Mitochondrial complex ii dysfunction can contribute significantly to genomic instability after exposure to ionizing radiation. Radiat. Res. 2009, 172, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lenton, E.A.; Sexton, L.; Cooke, I.D. The effect of age on the cyclical patterns of plasma LH, FSH, oestradiol and progesterone in women with regular menstrual cycles. Hum. Reprod. 1988, 3, 851–855. [Google Scholar] [PubMed]
- Wallace, M.; Hashim, Y.Z.; Wingfield, M.; Culliton, M.; McAuliffe, F.; Gibney, M.J.; Brennan, L. Effects of menstrual cycle phase on metabolomic profiles in premenopausal women. Hum. Reprod. 2010, 25, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Katz, F.H.; Romfh, P.; Smith, J.A. Diurnal variation of plasma aldosterone, cortisol and renin activity in supine man. J. Clin. Endocrinol. Metab. 1975, 40, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Limoli, C.L.; Giedzinski, E.; Morgan, W.F.; Swarts, S.G.; Jones, G.D.; Hyun, W. Persistent oxidative stress in chromosomally unstable cells. Cancer Res. 2003, 63, 3107–3111. [Google Scholar] [PubMed]
- Clutton, S.M.; Townsend, K.M.; Walker, C.; Ansell, J.D.; Wright, E.G. Radiation-induced genomic instability and persisting oxidative stress in primary bone marrow cultures. Carcinogenesis 1996, 17, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Thony, B.; Auerbach, G.; Blau, N. Tetrahydrobiopterin biosynthesis, regeneration and functions. Biochem. J. 2000, 347 Pt 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, M.J.; Tatham, A.L.; Hale, A.B.; Alp, N.J.; Channon, K.M. Critical role for tetrahydrobiopterin recycling by dihydrofolate reductase in regulation of endothelial nitric-oxide synthase coupling: Relative importance of the de novo biopterin synthesis versus salvage pathways. J. Biol. Chem. 2009, 284, 28128–28136. [Google Scholar] [CrossRef] [PubMed]
- Kietadisorn, R.; Juni, R.P.; Moens, A.L. Tackling endothelial dysfunction by modulating nos uncoupling: New insights into its pathogenesis and therapeutic possibilities. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E481–E495. [Google Scholar] [CrossRef] [PubMed]
- Moens, A.L.; Kass, D.A. Therapeutic potential of tetrahydrobiopterin for treating vascular and cardiac disease. J. Cardiovasc. Pharmacol. 2007, 50, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Pawar, S.A.; Fu, Q.; Gupta, P.K.; Berbee, M.; Garg, S.; Sridharan, V.; Wang, W.; Biju, P.G.; Krager, K.J.; et al. Characterization of transgenic Gfrp knock-in mice: Implications for tetrahydrobiopterin in modulation of normal tissue radiation responses. Antioxid. Redox Signal. 2014, 20, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.; Fu, Q.; Boerma, M.; Pathak, R.; Zhou, D.; Kumar, K.S.; Hauer-Jensen, M. Reduction of radiation-induced vascular nitrosative stress by the vitamin E analog γ-tocotrienol: Evidence of a role for tetrahydrobiopterin. Int. J. Radiat. Oncol. Biol. Phys. 2011, 79, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Kohnen, S.L.; Mouithys-Mickalad, A.A.; Deby-Dupont, G.P.; Deby, C.M.; Lamy, M.L.; Noels, A.F. Oxidation of tetrahydrobiopterin by peroxynitrite or oxoferryl species occurs by a radical pathway. Free Radic. Res. 2001, 35, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Kuzkaya, N.; Weissmann, N.; Harrison, D.G.; Dikalov, S. Interactions of peroxynitrite, tetrahydrobiopterin, ascorbic acid, and thiols: Implications for uncoupling endothelial nitric-oxide synthase. J. Biol. Chem. 2003, 278, 22546–22554. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Li, W.B.; Zhou, A.M.; Ai, J.; Huang, S.S. Role of endogenous peroxynitrite in pulmonary injury and fibrosis induced by bleomycin A5 in rats. Acta Pharmacol. Sin. 2003, 24, 697–702. [Google Scholar] [PubMed]
- Hanaue, N.; Takeda, I.; Kizu, Y.; Tonogi, M.; Yamane, G.Y. Peroxynitrite formation in radiation-induced salivary gland dysfunction in mice. Biomed. Res. 2007, 28, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Al-Nimer, M.S.; Ali, N.H. Assessment of nitrosative stress and lipid peroxidation activity in asymptomatic exposures to medical radiation: The bystander effect of ionizing radiation. Int. J. Appl. Basic Med. Res. 2012, 2, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Chalupsky, K.; Stefani, E.; Cai, H. Mechanistic insights into folic acid-dependent vascular protection: Dihydrofolate reductase (DHFR)-mediated reduction in oxidant stress in endothelial cells and angiotensin ii-infused mice: A novel hplc-based fluorescent assay for dhfr activity. J. Mol. Cell Cardiol. 2009, 47, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.M.; Dorrance, A.M.; Webb, R.C. Phenylalanine improves dilation and blood pressure in GTP cyclohydrolase inhibition-induced hypertensive rats. J. Cardiovasc. Pharmacol. 2004, 43, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.M.; Dorrance, A.M.; Mack, E.A.; Webb, R.C. Glucocorticoids decrease GTP cyclohydrolase and tetrahydrobiopterin-dependent vasorelaxation through glucocorticoid receptors. J. Cardiovasc. Pharmacol. 2004, 43, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Nandi, M.; Kelly, P.; Vallance, P.; Leiper, J. Over-expression of GTP-cyclohydrolase 1 feedback regulatory protein attenuates LPS and cytokine-stimulated nitric oxide production. Vasc. Med. 2008, 13, 29–36. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, Y.; Zhu, H.; Zou, M.H. Non-covalent interaction between polyubiquitin and GTP cyclohydrolase 1 dictates its degradation. PLoS One 2012, 7, e43306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, J.; Zhu, H.; Song, P.; Zou, M.H. Peroxynitrite-dependent zinc release and inactivation of guanosine 5′-triphosphate cyclohydrolase 1 instigate its ubiquitination in diabetes. Diabetes 2013, 62, 4247–4256. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rezvan, A.; Salerno, J.C.; Husain, A.; Kwon, K.; Jo, H.; Harrison, D.G.; Chen, W. GTP cyclohydrolase I phosphorylation and interaction with GTP cyclohydrolase feedback regulatory protein provide novel regulation of endothelial tetrahydrobiopterin and nitric oxide. Circ. Res. 2010, 106, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Kalivendi, S.; Hatakeyama, K.; Whitsett, J.; Konorev, E.; Kalyanaraman, B.; Vasquez-Vivar, J. Changes in tetrahydrobiopterin levels in endothelial cells and adult cardiomyocytes induced by LPS and hydrogen peroxide—A role for GFRP? Free Radic. Biol. Med. 2005, 38, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.R.; Bahrami, S.; Heller, R.; Werner-Felmayer, G. Bacterial lipopolysaccharide down-regulates expression of GTP cyclohydrolase I feedback regulatory protein. J. Biol. Chem. 2002, 277, 10129–10133. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.K.; Pathak, R.; Zandkarimi, F.; Kaur, P.; Alkhalil, L.; Singh, R.; Zhong, X.; Ghosh, S.; Aykin-Burns, N.; Hauer-Jensen, M. Liver metabolomics reveals increased oxidative stress and fibrogenic potential in GFRP transgenic mice in response to ionizing radiation. J. Proteome Res. 2014, 13, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Tatham, A.L.; Crabtree, M.J.; Warrick, N.; Cai, S.; Alp, N.J.; Channon, K.M. GTP cyclohydrolase I expression, protein, and activity determine intracellular tetrahydrobiopterin levels, independent of gtp cyclohydrolase feedback regulatory protein expression. J. Biol. Chem. 2009, 284, 13660–13668. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, H.; Ikemoto, K.; Tada, S.; Udagawa, Y.; Ohtsuki, M.; Sumi-Ichinose, C.; Kondo, K.; Nomura, T. Cilostazol inhibits cytokine-induced tetrahydrobiopterin biosynthesis in human umbilical vein endothelial cells. J. Atheroscler. Thromb. 2011, 18, 312–317. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Magne, N.; Toillon, R.A.; Bottero, V.; Didelot, C.; Houtte, P.V.; Gerard, J.P.; Peyron, J.F. NF-κB modulation and ionizing radiation: Mechanisms and future directions for cancer treatment. Cancer Lett. 2006, 231, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Nagane, M.; Yasui, H.; Sakai, Y.; Yamamori, T.; Niwa, K.; Hattori, Y.; Kondo, T.; Inanami, O. Activation of eNOS in endothelial cells exposed to ionizing radiation involves components of the DNA damage response pathway. Biochem. Biophys. Res. Commun. 2015, 456, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S.; Matsuda, S.; Gunji, M.; Kamogawa, A. The role of nitric oxide in radiation damage. Biol. Pharm. Bull. 2007, 30, 1102–1107. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, R.K.; Suh, Y.; Cui, Y.H.; Hwang, E.; Lim, E.J.; Yoo, K.C.; Lee, G.H.; Yi, J.M.; Kang, S.G.; Lee, S.J. Fractionated radiation-induced nitric oxide promotes expansion of glioma stem-like cells. Cancer Sci. 2013, 104, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Ibuki, Y.; Mizuno, S.; Goto, R. Gamma-irradiation-induced DNA damage enhances no production via NF-κB activation in raw264.7 cells. Biochim. Biophys. Acta 2003, 1593, 159–167. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Landmesser, U.; Dikalov, S.; Price, S.R.; McCann, L.; Fukai, T.; Holland, S.M.; Mitch, W.E.; Harrison, D.G. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J. Clin. Investig. 2003, 111, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Zhang, Y.Y.; Chen, K.; Hatakeyama, K.; Keaney, J.F., Jr. Cytokine-stimulated GTP cyclohydrolase I expression in endothelial cells requires coordinated activation of nuclear factor-κB and Stat1/Stat3. Circ. Res. 2005, 96, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.; Pellegatta, S.; Favaro, R.; Pisati, F.; Roncaglia, P.; Testa, G.; Nicolis, S.K.; Finocchiaro, G.; D’Adda di Fagagna, F. DNA damage in mammalian neural stem cells leads to astrocytic differentiation mediated by BMP2 signaling through JAK-STAT. Stem Cell Rep. 2013, 1, 123–138. [Google Scholar] [CrossRef]
- Dawson, L.A.; Ten Haken, R.K.; Lawrence, T.S. Partial irradiation of the liver. Semin. Radiat. Oncol. 2001, 11, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.X.; Zhao, H.B.; Li, C.R.; Sun, Y.N.; Qian, X.H.; Wang, S.Q. Proteomic analysis of ionizing radiation-induced proteins at the subcellular level. J. Proteome Res. 2009, 8, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.A.; Normolle, D.; Balter, J.M.; McGinn, C.J.; Lawrence, T.S.; Ten Haken, R.K. Analysis of radiation-induced liver disease using the Lyman NTCP model. Int. J. Radiat. Oncol. Biol. Phys. 2002, 53, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Wu, J.K.; Huang, C.M.; Liu, H.S.; Huang, D.Y.; Cheng, S.H.; Tsai, S.Y.; Jian, J.J.; Lin, Y.M.; Cheng, T.I.; et al. Radiation-induced liver disease after three-dimensional conformal radiotherapy for patients with hepatocellular carcinoma: Dosimetric analysis and implication. Int. J. Radiat. Oncol. Biol. Phys. 2002, 54, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Anscher, M.S.; Crocker, I.R.; Jirtle, R.L. Transforming growth factor-β1 expression in irradiated liver. Radiat. Res. 1990, 122, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Flechsig, P.; Dadrich, M.; Bickelhaupt, S.; Jenne, J.; Hauser, K.; Timke, C.; Peschke, P.; Hahn, E.W.; Grone, H.J.; Yingling, J.; et al. Ly2109761 attenuates radiation-induced pulmonary murine fibrosis via reversal of TGF-β and BMP-associated proinflammatory and proangiogenic signals. Clin. Cancer Res. 2012, 18, 3616–3627. [Google Scholar] [CrossRef] [PubMed]
- Boothe, D.L.; Coplowitz, S.; Greenwood, E.; Barney, C.L.; Christos, P.J.; Parashar, B.; Nori, D.; Chao, K.S.; Wernicke, A.G. Transforming growth factor β-1 (TGF-β1) is a serum biomarker of radiation induced fibrosis in patients treated with intracavitary accelerated partial breast irradiation: Preliminary results of a prospective study. Int. J. Radiat. Oncol. Biol. Phys. 2013, 87, 1030–1036. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Winter-de Groot, K.M.; van Haren Noman, S.; Speleman, L.; Schilder, A.G.; van der Ent, C.K. Nasal nitric oxide levels and nasal polyposis in children and adolescents with cystic fibrosis. JAMA Otolaryngol. Head Neck Surg. 2013, 139, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Michl, R.K.; Hentschel, J.; Fischer, C.; Beck, J.F.; Mainz, J.G. Reduced nasal nitric oxide production in cystic fibrosis patients with elevated systemic inflammation markers. PLoS One 2013, 8, e79141. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moens, A.L.; Takimoto, E.; Tocchetti, C.G.; Chakir, K.; Bedja, D.; Cormaci, G.; Ketner, E.A.; Majmudar, M.; Gabrielson, K.; Halushka, M.K.; et al. Reversal of cardiac hypertrophy and fibrosis from pressure overload by tetrahydrobiopterin: Efficacy of recoupling nitric oxide synthase as a therapeutic strategy. Circulation 2008, 117, 2626–2636. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Vivar, J.; Martasek, P.; Whitsett, J.; Joseph, J.; Kalyanaraman, B. The ratio between tetrahydrobiopterin and oxidized tetrahydrobiopterin analogues controls superoxide release from endothelial nitric oxide synthase: An epr spin trapping study. Biochem. J. 2002, 362, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.; Unbehaun, A.; Schellenberg, B.; Mayer, B.; Werner-Felmayer, G.; Werner, E.R. l-ascorbic acid potentiates endothelial nitric oxide synthesis via a chemical stabilization of tetrahydrobiopterin. J. Biol. Chem. 2001, 276, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nakanishi, N.; Akimoto, K.; Yoshida, M.; Kasai, K. HMG-CoA reductase inhibitor increases GTP cyclohydrolase I mrna and tetrahydrobiopterin in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.P.; Kim, Y.M.; Sowa, M.B.; Robinson, R.J.; Gao, X.; Metz, T.O.; Morgan, W.F.; Zhang, Q. Metabolomic response of human skin tissue to low dose ionizing radiation. Mol. Biosyst. 2012, 8, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Z.; Zhang, X.; Qiao, Y.; Wu, S.; Dong, F.; Chen, Y. Selection of candidate radiation biomarkers in the serum of rats exposed to γ-rays by GC/TOFMS-based metabolomics. Radiat. Prot. Dosim. 2013, 154, 9–17. [Google Scholar] [CrossRef]
- Johnson, C.H.; Patterson, A.D.; Krausz, K.W.; Kalinich, J.F.; Tyburski, J.B.; Kang, D.W.; Luecke, H.; Gonzalez, F.J.; Blakely, W.F.; Idle, J.R. Radiation metabolomics. 5. Identification of urinary biomarkers of ionizing radiation exposure in nonhuman primates by mass spectrometry-based metabolomics. Radiat. Res. 2012, 178, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Rana, P.; Devi, M.M.; Chaturvedi, S.; Javed, S.; Tripathi, R.P.; Khushu, S. Nuclear magnetic resonance spectroscopy-based metabonomic investigation of biochemical effects in serum of γ-irradiated mice. Int. J. Radiat. Biol. 2011, 87, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Patterson, A.D.; Krausz, K.W.; Lanz, C.; Kang, D.W.; Luecke, H.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 4. UPLC-ESI-QTOFMS-Based metabolomics for urinary biomarker discovery in γ-irradiated rats. Radiat. Res. 2011, 175, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Lanz, C.; Patterson, A.D.; Slavik, J.; Krausz, K.W.; Ledermann, M.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 3. Biomarker discovery in the urine of γ-irradiated rats using a simplified metabolomics protocol of gas chromatography-mass spectrometry combined with random forests machine learning algorithm. Radiat. Res. 2009, 172, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Tyburski, J.B.; Patterson, A.D.; Krausz, K.W.; Slavik, J.; Fornace, A.J., Jr.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 2. Dose- and time-dependent urinary excretion of deaminated purines and pyrimidines after sublethal γ-radiation exposure in mice. Radiat. Res. 2009, 172, 42–57. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pathak, R.; Cheema, A.K.; Boca, S.M.; Krager, K.J.; Hauer-Jensen, M.; Aykin-Burns, N. Modulation of Radiation Response by the Tetrahydrobiopterin Pathway. Antioxidants 2015, 4, 68-81. https://doi.org/10.3390/antiox4010068
Pathak R, Cheema AK, Boca SM, Krager KJ, Hauer-Jensen M, Aykin-Burns N. Modulation of Radiation Response by the Tetrahydrobiopterin Pathway. Antioxidants. 2015; 4(1):68-81. https://doi.org/10.3390/antiox4010068
Chicago/Turabian StylePathak, Rupak, Amrita K. Cheema, Simina M. Boca, Kimberly J. Krager, Martin Hauer-Jensen, and Nukhet Aykin-Burns. 2015. "Modulation of Radiation Response by the Tetrahydrobiopterin Pathway" Antioxidants 4, no. 1: 68-81. https://doi.org/10.3390/antiox4010068
APA StylePathak, R., Cheema, A. K., Boca, S. M., Krager, K. J., Hauer-Jensen, M., & Aykin-Burns, N. (2015). Modulation of Radiation Response by the Tetrahydrobiopterin Pathway. Antioxidants, 4(1), 68-81. https://doi.org/10.3390/antiox4010068