Endogenous Phenolics in Hulls and Cotyledons of Mustard and Canola: A Comparative Study on Its Sinapates and Antioxidant Capacity

Abstract
:1. Introduction
2. Experimental Section
2.1. Chemicals and Materials
2.2. Phenolic Extraction from Hulls and Cotyledons of Canola and Mustard
2.3. Total Phenolic Content of Canola and Mustard Seed Fractions
2.4. DPPH Scavenging Activity
2.5. HPLC-DAD Analysis of Phenolic Compounds in Canola and Mustard Extracts
2.6. UPLC-MS Analysis of Phenolics from Hulls and Cotyledons
2.7. Data Expression and Analysis
3. Results and Discussion
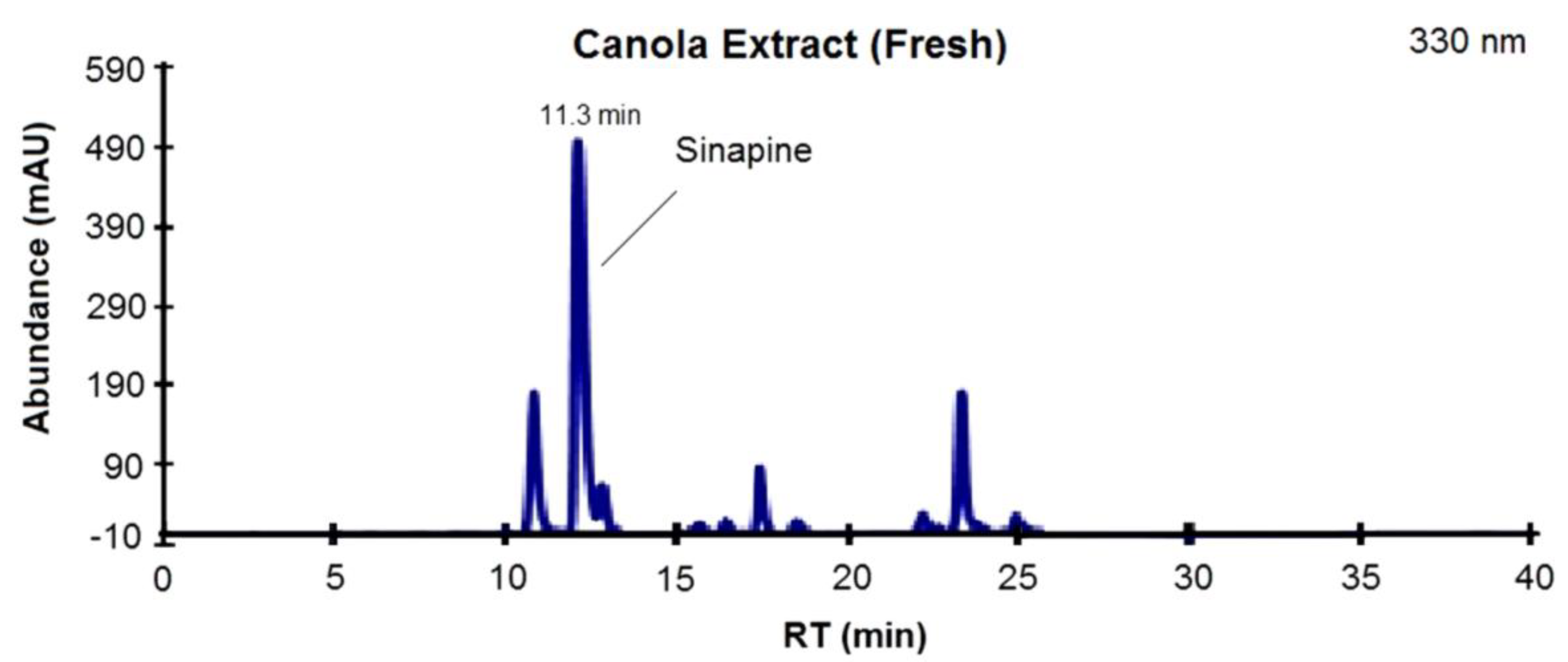
3.1. Phenolic Profile of Extracts from Canola and Mustard Seed Fractions

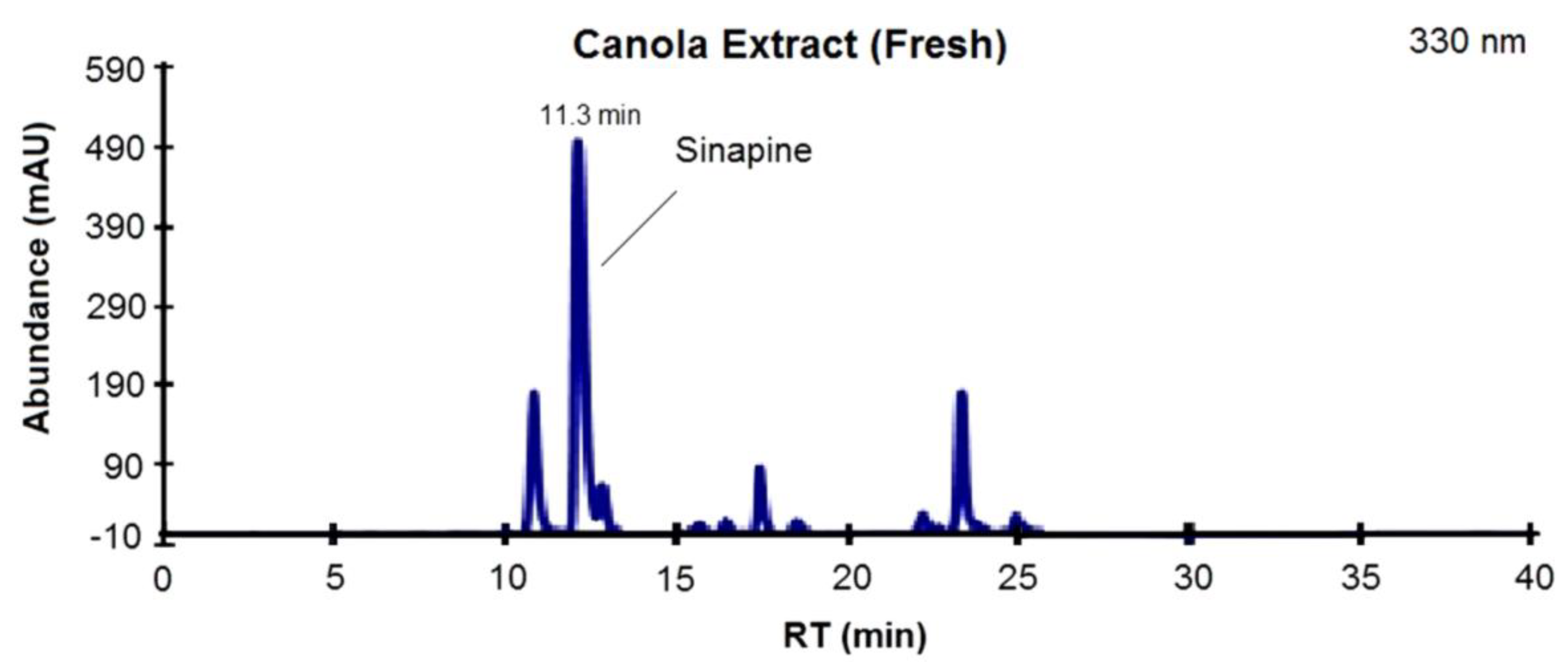{kind=link}
{kind=link}
{kind=link}
| Samples | Sinapoyl Glucose | Sinapine | Sinapic Acid | Total Phenolics (HPLC) * | Total Phenolics Folin-Ciocalteu | EC50 |
|---|---|---|---|---|---|---|
| Canola Cotyledon | 8.71 ± 0.76 a | 12.20 ± 1.16 a | 0.22 ± 0.02 b | 20.20 ± 1.85 a | 16.89 ± 0.69 a | 1.78 |
| Canola Seeds | 5.45 ± 0.35 b | 8.35 ± 0.44 c | 0.15 ± 0.01 d | 14.06 ± 0.71 b | 10.60 ± 0.81 b | 2.31 |
| Canola Hulls | 1.34 ± 0.07 c | 1.15 ± 0.07 e | 0.04 ± 0.00 e | 3.57 ± 0.20 d | 4.50 ± 0.16 d | 5.82 |
| Mustard Cotyledon | 0.67 ± 0.01 c | 10.62 ± 0.08 b | 0.18 ± 0.00 c | 11.45 ± 0.05 c | 10.60 ± 0.24 b | 2.36 |
| Mustard Seeds | 0.66 ± 0.01 c | 10.17 ± 0.27 b | 0.19 ± 0.02 c | 11.12 ± 0.39 c | 10.31 ± 0.32 b | 2.54 |
| Mustard Hulls | 0.41 ± 0.02 c | 4.74 ± 0.28 d | 0.90 ± 0.01 a | 5.67 ± 0.32 d | 6.24 ± 0.40 c | 4.40 |
| Samples | Oil content % (Dry wt) | Moiture % |
|---|---|---|
| Canola Cotyledon | 53.82 ± 0.66 a | 3.17 ± 0.00 f |
| Canola Seeds | 41.70 ± 0.62 b | 6.45 ± 0.09 c |
| Mustard Seeds | 38.35 ± 1.08 c | 5.10 ± 0.11 e |
| Mustard Hulls | 27.69 ± 0.07 d | 5.53 ± 0.13 d |
| Canola Hulls | 18.00 ± 0.09 e | 6.71 ± 0.04 b |
| Mustard Cotyledon | 42.18 ± 0.00 b | 7.06 ± 0.05 a |
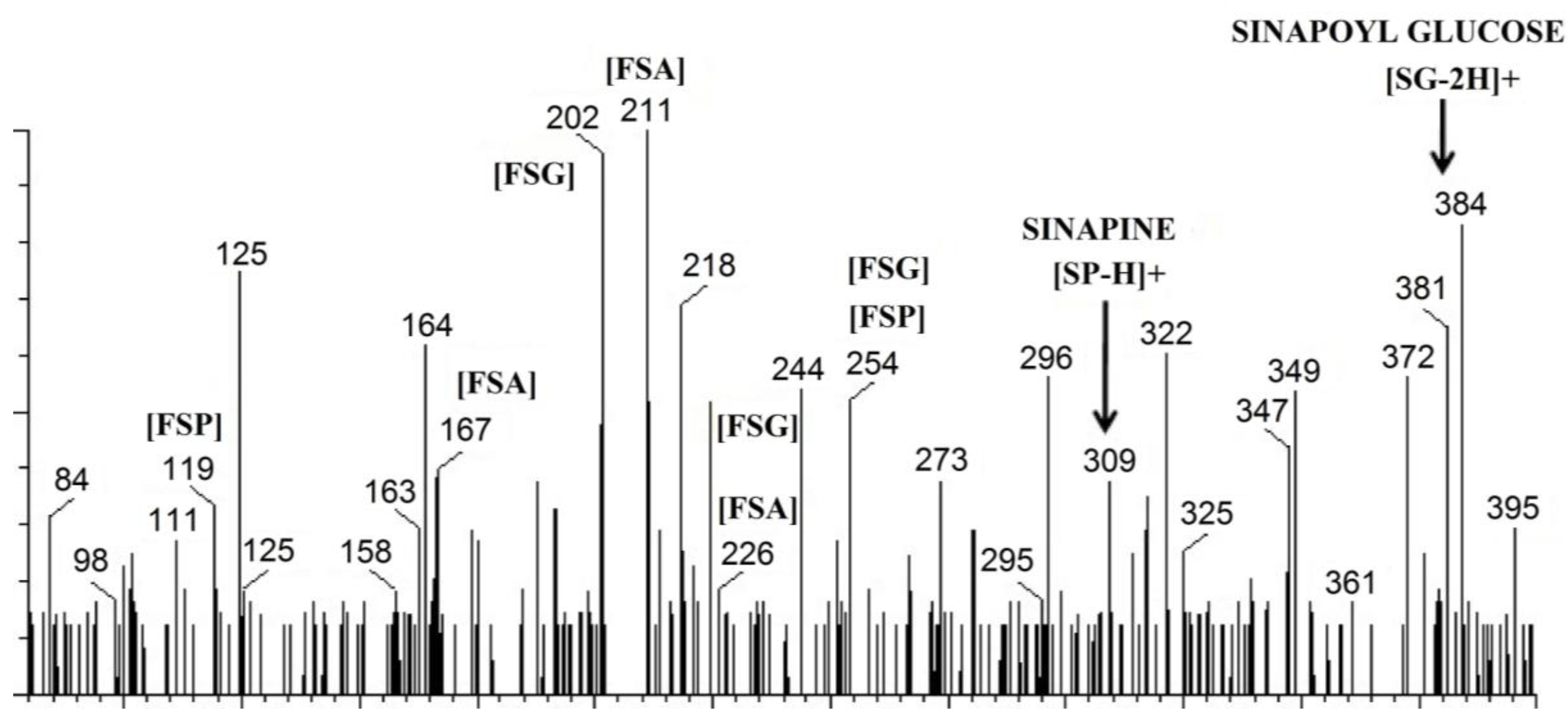
3.2. UPLC-MS Analysis of Phenolics from Mustard and Canola Cotyledons and Hulls

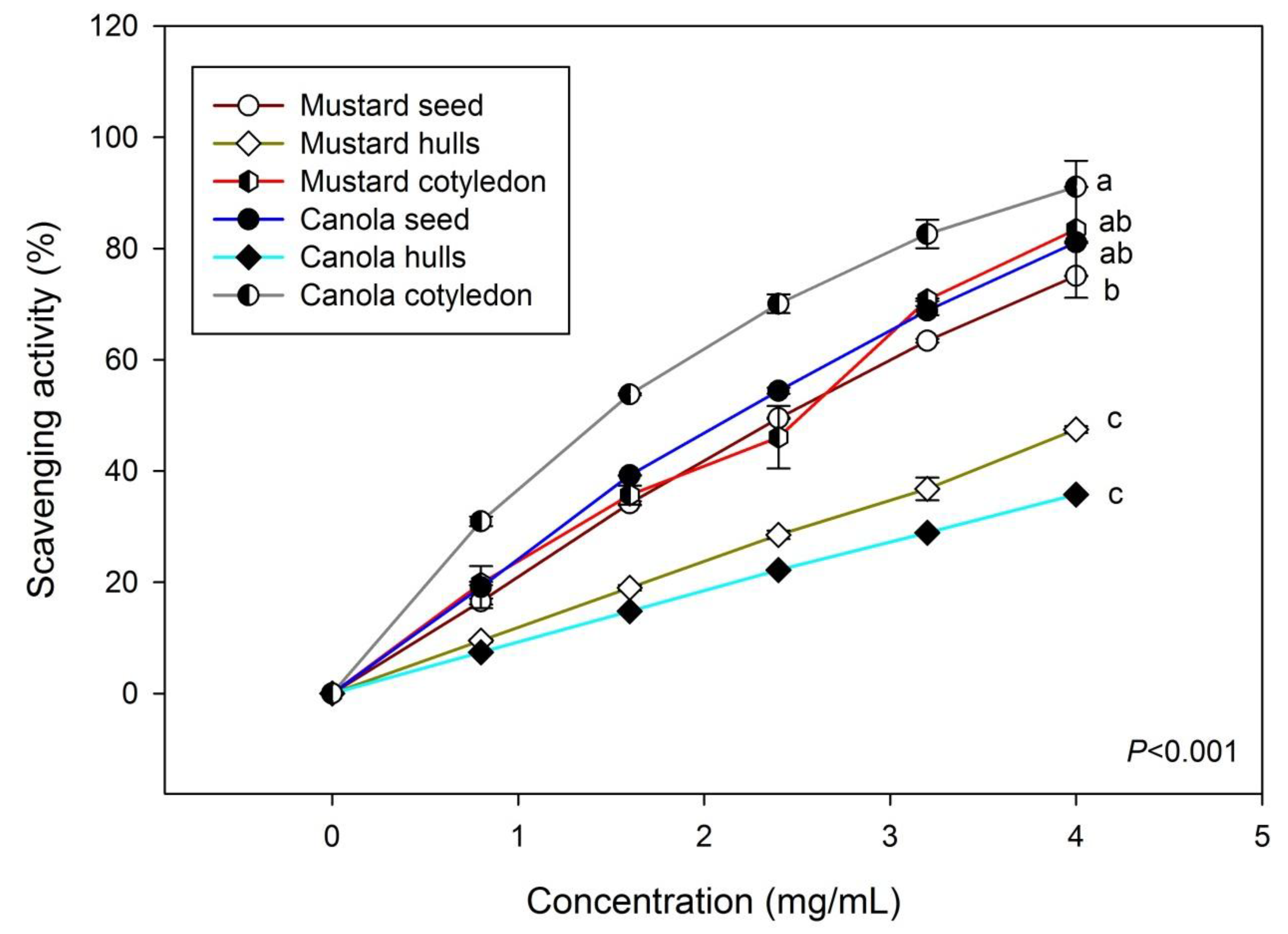
3.3. DPPH Scavenging Effects of Various Phenolic Extracts
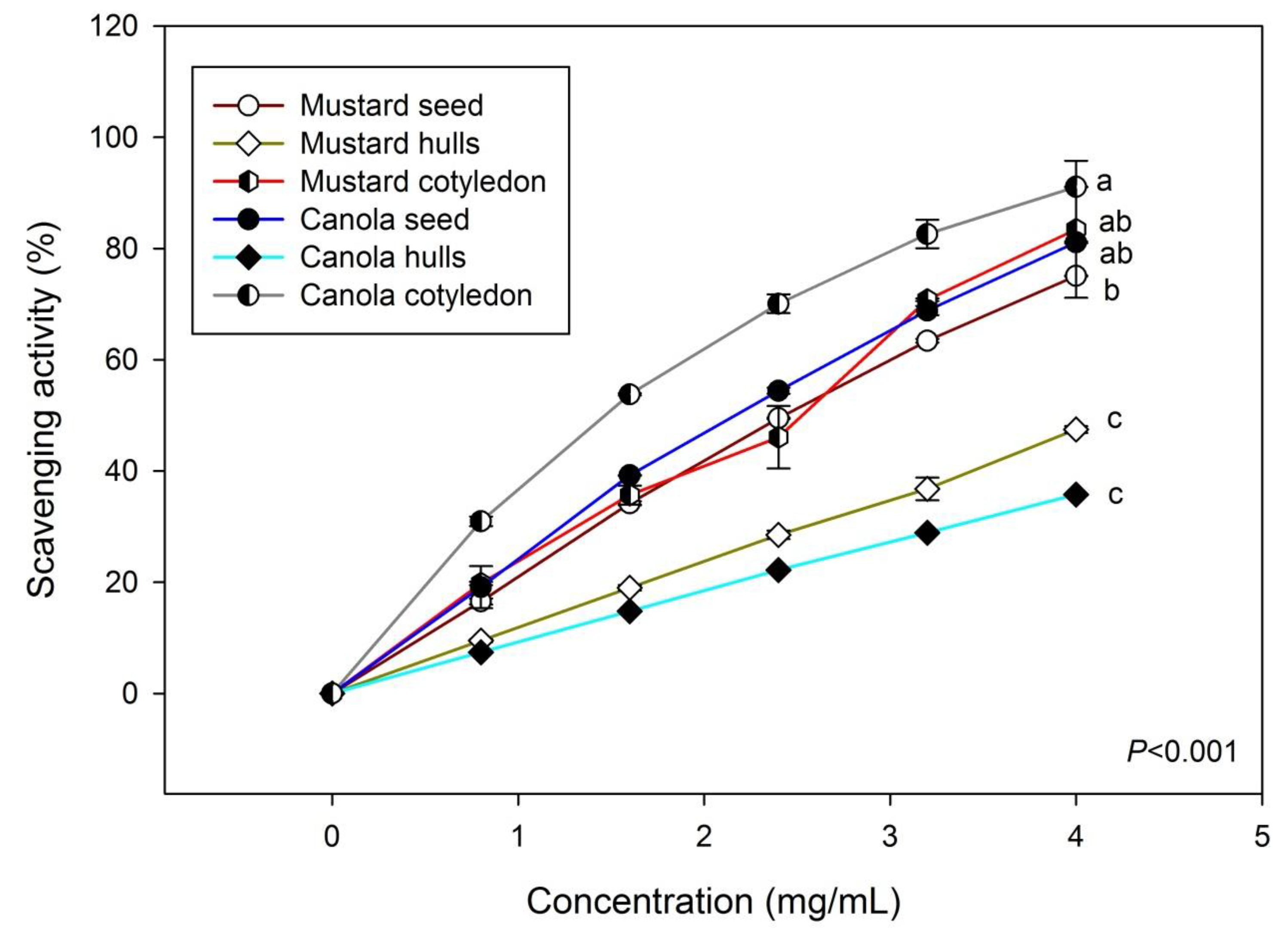
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals—Just antioxidants or much more? Cell Mol Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Aachary, A.A.; Thiyam-Hollander, U. An update on characterization and bioactivities of sinapic acid derivatives. In Canola and Rapeseed: Production, Processing, Food Quality, and Nutrition; Thiyam-Hollander, U., Eskin, N.A.M., Matthaus, B., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 21–38. [Google Scholar]
- Thiyam, U.; Claudia, P.; Jan, U.; Alfred, B. De-oiled rapeseed and a protein isolate: Characterization of sinapic acid derivatives by HPLC-DAD and LC-MS. Eur. Food Res. Tech. 2009, 229, 825–831. [Google Scholar] [CrossRef]
- Matthäus, B. Antioxidant activity of extracts obtained from residues of different oilseeds. J. Agric. Food Chem. 2002, 50, 3444–3452. [Google Scholar] [CrossRef] [PubMed]
- Baumert, A.; Milkowski, C.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Formation of a complex pattern of sinapate esters in Brassica napus seeds, catalyzed by enzymes of a serine carboxypeptidase-like acyltransferase family? Phytochemistry 2005, 66, 1334–1345. [Google Scholar] [CrossRef]
- Wanasundara, U.N.; Shahidi, F. Canola extract as an alternative natural antioxidant for canola oil. J. Am. Oil Chem. Soc. 1994, 71, 817–822. [Google Scholar] [CrossRef]
- Natella, F.; Nardini, M.; di Felice, M.; Scaccini, C. Benzoic and cinnamic acid derivatives as antioxidants: Structure-activity relation. J. Agric. Food Chem. 1999, 47, 1453–1459. [Google Scholar] [CrossRef]
- Cuvelier, M.E.; Richard, H.; Berset, C. Comparison of the antioxidative activity of some acid-phenols: Structure-activity relationship. Biosci. Biotechnol. Biochem. 1992, 56, 324–325. [Google Scholar] [CrossRef]
- Marinova, E.M.; Yanishlieva, N.V. Effect of lipid unsaturation on the oxidative activity of some phenolic acids. J. Am. Oil Chem. Soc. 1994, 71, 427–434. [Google Scholar] [CrossRef]
- Pekkarinen, S.S.; Stöckmann, H.; Schwarz, K.; Heinonen, I.M.; Hopia, A.I. Antioxidant activity and partitioning of phenolic acids in bulk and emulsified methyl linoleate. J. Agric. Food Chem. 1999, 47, 3036–3043. [Google Scholar] [CrossRef]
- Wanasundara, U.N.; Amarowicz, R.; Shahidi, F. Partial characterization of natural antioxidants in canola meal. Food Res. Int. 1995, 28, 525–530. [Google Scholar] [CrossRef]
- Amarowicz, R.; Raab, B.; Shahidi, F. Antioxidant activity of phenolic fractions of rapeseed. J. Food Lipids. 2003, 10, 51–62. [Google Scholar] [CrossRef]
- Koski, A.; Pekkarinen, S.; Hopia, A.; Wähälä, K.; Heinonen, M. Processing of rapeseed oil: Effects on sinapic acid derivative content and oxidative stability. Eur. Food Res. Technol. 2003, 217, 110–114. [Google Scholar] [CrossRef]
- Vuorela, S.; Meyer, A.S.; Heinonen, M. Impact of isolation method on the antioxidant activity of rapeseed meal phenolics. J. Agric. Food Chem. 2004, 52, 8202–8207. [Google Scholar] [CrossRef]
- Naczk, M.; Amarowicz, R.; Zadernowski, R.; Shahidi, F. Antioxidant capacity of phenolics from canola hulls as affected by different solvents. In Phenolic Compounds in Foods and Natural Health Products; Shahidi, F.C.-H., Ed.; ACS Symposium Series: Washington, DC, USA, 2005; Volume 909, pp. 57–66. [Google Scholar]
- Vuorela, S.; Kreander, K.; Karonen, M.; Nieminen, R.; Hämäläinen, M.; Galkin, A.; Laitinen, L.; Salminen, J.; Moilanen, E.; Pihlaja, K.; et al. Preclinical evaluation of rapeseed, raspberry, and pine bark phenolics for health related effects. J. Agric. Food Chem. 2005, 53, 5922–5931. [Google Scholar] [CrossRef]
- Wakamatsu, D.; Morimura, S.; Sawa, T.; Kida, K.; Nakai, C.; Maeda, H. Isolation, identification, and structure of a potent alkyl-peroxyl radical scavenger in crude canola oil, canolol. Biosci. Biotechnol. Biochem. 2005, 69, 1568–1574. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Wagner, A.; Jahreis, G. Influence of thermal treatment of rapeseed on the canolol content. Food Chem. 2009, 112, 944–948. [Google Scholar] [CrossRef]
- Galano, A.; Francisco-Márquez, M.; Alvarez-Idaboy, J.R. Canolol: A promising chemical agent against oxidative stress. J. Phys. Chem. B 2011, 115, 8590–8596. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, H.; Kanazawa, A.; Wakamatu, D.; Morimura, S.; Kida, K.; Akaike, T.; Maeda, H. Antioxidative and antimutagenic activities of 4-vinyl-2,6-dimethoxyphenol (canolol) isolated from canola oil. J. Agric. Food Chem. 2004, 52, 4380–4387. [Google Scholar] [CrossRef]
- Bala, M.; Singh, M. Non destructive estimation of total phenol and crude fiber content in intact seeds of rapeseed-mustard using FTNIR. Ind. Crop. Prod. 2013, 42, 357–362. [Google Scholar] [CrossRef]
- Escarpa, A.; González, M.C. Evaluation of high-performance liquid chromatography for determination of phenolic compounds in pear horticultural cultivars. Chromatographia 2000, 51, 37–43. [Google Scholar] [CrossRef]
- Cai, R.; Arntfield, S.D. A rapid high-performance liquid chromatographic method for the determination of sinapine and sinapic acid in canola seed and meal. J. Am. Oil Chem. Soc. 2001, 78, 903–910. [Google Scholar] [CrossRef]
- Mattila, P.; Kumpulainen, J. Determination of free and total phenolic acids in plant-derived foods by HPLC with diode-array detection. J. Agric. Food Chem. 2002, 50, 3660–3667. [Google Scholar] [CrossRef]
- Thiyam, U.; Stöckmann, H.; Zum Felde, T.; Schwarz, K. Antioxidative effect of the main sinapic acid derivatives from rapeseed and mustard oil by-products. Eur. J. Lipid Sci. Technol. 2006, 108, 239–248. [Google Scholar] [CrossRef]
- Wen, D.; Li, C.; Di, H.; Liao, Y.; Liu, H. A universal HPLC method for the determination of phenolic acids in compound herbal medicines. J. Agric. Food Chem. 2005, 53, 6624–6629. [Google Scholar] [CrossRef]
- Yang, M.; Zheng, C.; Zhou, Q.; Huang, F.; Liu, C.; Wang, H. Minor components and oxidative stability of cold-pressed oil from rapeseed cultivars in China. J. Food Com. Anal. 2013, 29, 1–9. [Google Scholar] [CrossRef]
- Thiyam, U.; Kuhlmann, A.; Stöckmann, H.; Schwarz, K. Prospects of rapeseed oil by-products with respect to antioxidative potential. Comptes Rendus Chim. 2004, 7, 611–616. [Google Scholar] [CrossRef]
- Khattab, R.; Eskin, M.; Aliani, M.; Thiyam, U. Determination of sinapic acid derivatives in canola extracts using high-performance liquid chromatography. J. Am. Oil Chem. Soc. 2010, 87, 147–155. [Google Scholar] [CrossRef]
- Schanderl, S.H. Tannins and related phenolics. In Food Science. A Series of Monograps; Stewart, G.F., Chichester, C.O., Galliver, G.B., Morgan, A.I., Eds.; Academic Press: New York, NY, USA, 1970; pp. 701–725. [Google Scholar]
- Schwarz, K.; Bertelsen, G.; Nissen, L.R.; Gardner, P.T.; Heinonen, M.I.; Hopia, A.; Huynh-Ba, T.; Lambelet, P.; McPhail, D.; Skibsted, L.H.; et al. Investigation of plant extracts for the protection of processed foods against lipid oxidation. Comparison of antioxidant assays based on radical scavenging, lipid oxidation and analysis of the principal antioxidant compounds. Eur. Food Res. Technol. 2001, 212, 319–328. [Google Scholar] [CrossRef]
- Krygier, K.; Sosulski, F.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 2. Composition of phenolic acids in rapeseed flour and hulls. J. Agric. Food Chem. 1982, 30, 334–336. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, L.; Pu, H.; Li, C.; Hu, Q. Profile and distribution of soluble and insoluble phenolics in Chinese rapeseed (Brassica napus). Food Chem. 2012, 135, 616–622. [Google Scholar] [CrossRef]
- Lin, L.Z.; Sun, J.; Chen, P.; Harnly, J. UHPLC-PDA-ESI/ HRMS/MSn analysis of anthocyanins, flavonol glycosides, and hydroxycinnamic acid derivatives in red mustard greens (Brassica juncea coss variety). J. Agric. Food Chem. 2011, 59, 12059–12072. [Google Scholar] [CrossRef]
- Bouchereau, A.; Hamelin, J.; Lamour, I.; Renard, M.; Larher, F. Distribution of sinapine and related compounds in seeds of Brassica and allied genera. Phytochemistry 1991, 30, 1873–1881. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [Green Version]
- Milkowski, C.; Strack, D. Sinapate esters in brassicaceous plants: Biochemistry, molecular biology, evolution and metabolic engineering. Planta 2010, 232, 19–35. [Google Scholar] [CrossRef]
- Shahidi, F.; Naczk, M. An overview of the phenolics of canola and rapeseed: Chemical, sensory and nutritional significance. J. Am. Oil Chem. Soc. 1992, 69, 917–924. [Google Scholar] [CrossRef]
- Kozlowska, H.; Naczk, M.; Shahidi, F.; Zadernowski, R. Phenolic acids and tannins in rapeseed and canola. In Canola and Rapeseed; Production, Chemistry, Nutrition and Processing Technology; Shahidi, F., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1990; pp. 193–210. [Google Scholar]
- Koski, A.; Psomiadou, E.; Tsimidou, M.; Hopia, A.; Kefalas, P.; Wähälä, K.; Heinonen, M. Oxidative stability and minor constituents of virgin olive oil and cold-pressed rapeseed oil. Eur. Food Res. Technol. 2002, 214, 294–298. [Google Scholar] [CrossRef]
- Siger, A.; Czubinski, J.; Dwiecki, K.; Kachlicki, P.; Nogala-Kalucka, M. Identification and antioxidant activity of sinapic acid derivatives in Brassica napus L. seed meal extracts. Eur. J. Lipid Sci. Technol. 2013, 115, 1130–1138. [Google Scholar]
- Shrestha, K.; Stevens, C.V.; de Meulenaer, B. Isolation and identification of a potent radical scavenger (canolol) from roasted high erucic mustard seed oil from nepal and its formation during roasting. J. Agric. Food Chem. 2012, 60, 7506–7512. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, K.; Gemechu, F.G.; de Meulenaer, B. A novel insight on the high oxidative stability of roastedmustard seed oil in relation to phospholipid, Maillard type reaction products, tocopherol and canolol contents. Food Res. Int. 2013, 54, 587–594. [Google Scholar] [CrossRef]
- Marles, M.A.S.; Gruber, M.Y.; Scoles, G.J.; Muir, A.D. Pigmentation in the developing seed coat and seedling leaves of Brassica carinata is controlled at the dihydroflavonol reductase level. Phytochemistry 2003, 62, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Schieber, A.; Gänzle, M.G. Sinapic acid derivatives in defatted Oriental mustard (Brassica juncea L.) seed meal extracts using UHPLC-DAD-ESI-MSn and identification of compounds with antibacterial activity. Eur. Food Res. Technol. 2012, 234, 535–542. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Chen, C.W.; Ho, C.T. Antioxidant properties of polyphenols extracted from green and black teas. J. Food Lipids. 1995, 2, 35–46. [Google Scholar] [CrossRef]
- Amarowicz, R.; Naczk, M.; Shahidi, F. Antioxidant activity of crude tannins of canola and rapeseed hulls. J. Am. Oil Chem. Soc. 2000, 77, 957–961. [Google Scholar] [CrossRef]
- Khattab, R.; Goldberg, E.; Lin, L.; Thiyam, U. Quantitative analysis and free-radical-scavenging activity of chlorophyll, phytic acid, and condensed tannins in canola. Food Chem. 2010, 122, 1266–1272. [Google Scholar] [CrossRef]
- Naczk, M.; Pegg, R.B.; Zadernowski, R.; Shahidi, F. Radical scavenging activity of canola hull phenolics. J. Am. Oil Chem. Soc. 2005, 82, 255–260. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mayengbam, S.; Aachary, A.; Thiyam-Holländer, U. Endogenous Phenolics in Hulls and Cotyledons of Mustard and Canola: A Comparative Study on Its Sinapates and Antioxidant Capacity. Antioxidants 2014, 3, 544-558. https://doi.org/10.3390/antiox3030544
Mayengbam S, Aachary A, Thiyam-Holländer U. Endogenous Phenolics in Hulls and Cotyledons of Mustard and Canola: A Comparative Study on Its Sinapates and Antioxidant Capacity. Antioxidants. 2014; 3(3):544-558. https://doi.org/10.3390/antiox3030544
Chicago/Turabian StyleMayengbam, Shyamchand, Ayyappan Aachary, and Usha Thiyam-Holländer. 2014. "Endogenous Phenolics in Hulls and Cotyledons of Mustard and Canola: A Comparative Study on Its Sinapates and Antioxidant Capacity" Antioxidants 3, no. 3: 544-558. https://doi.org/10.3390/antiox3030544