Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing



{kind=link}
{kind=link}
{kind=link}
Abstract
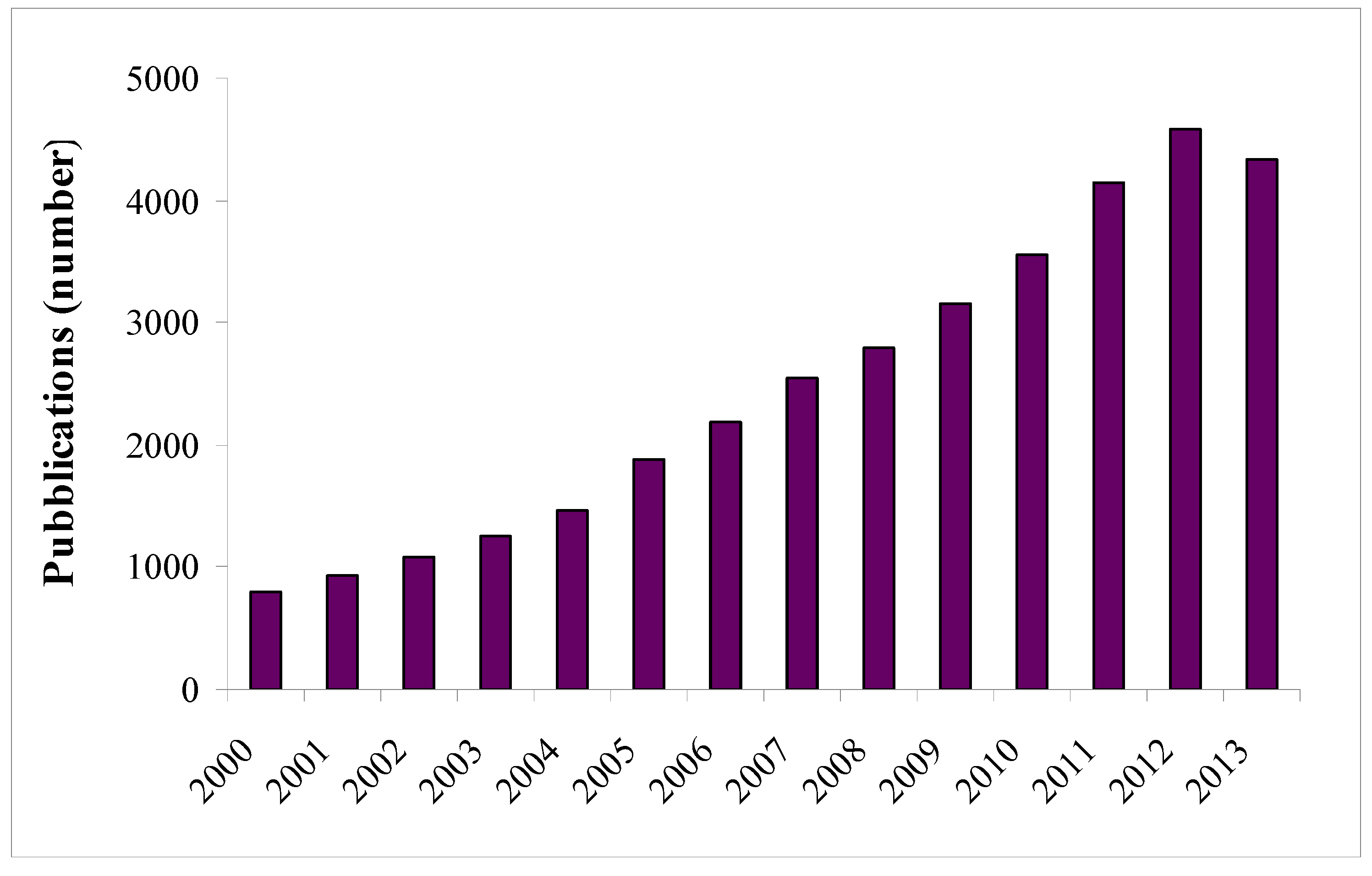
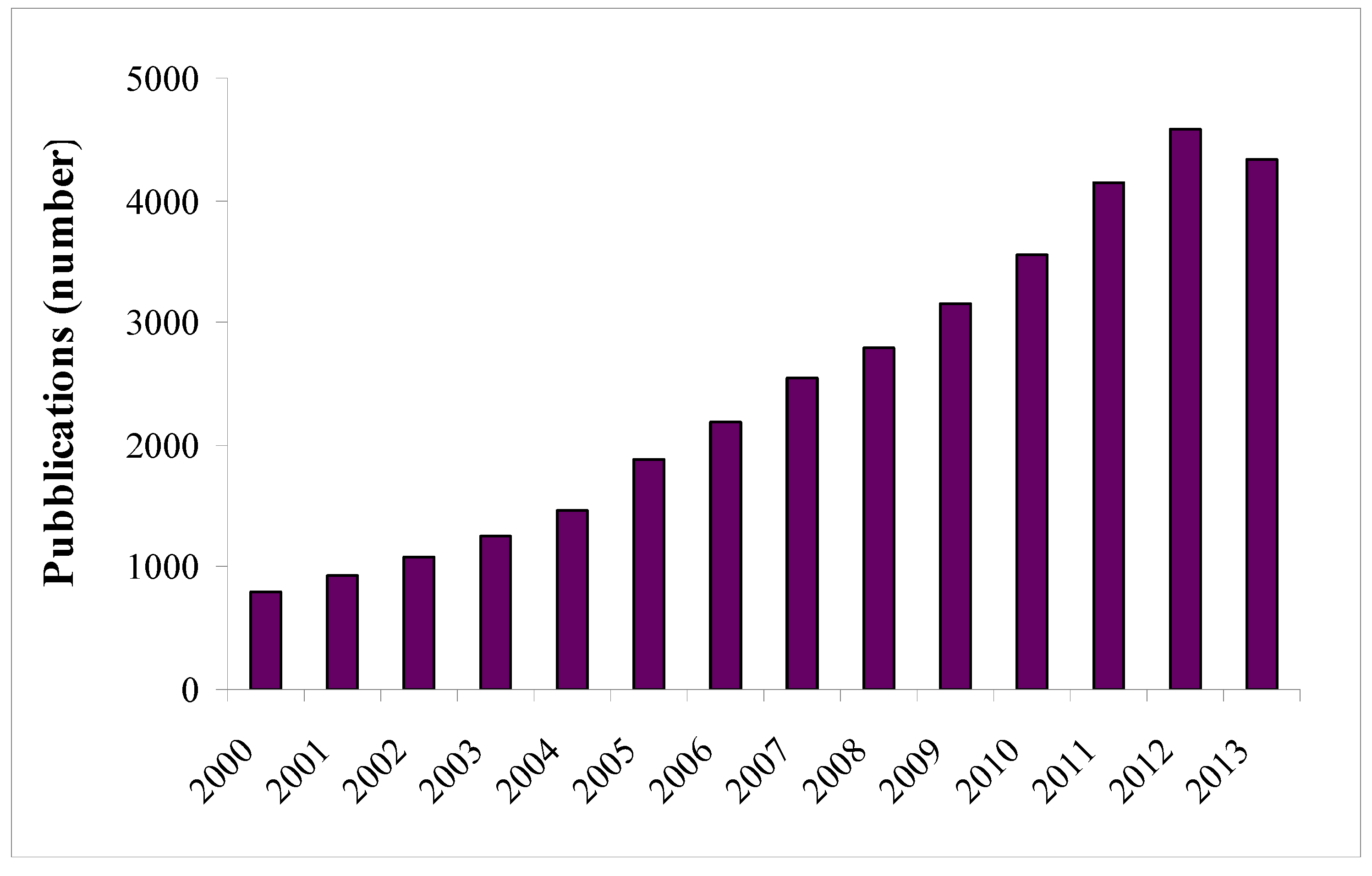
:1. Introduction
2. Apple
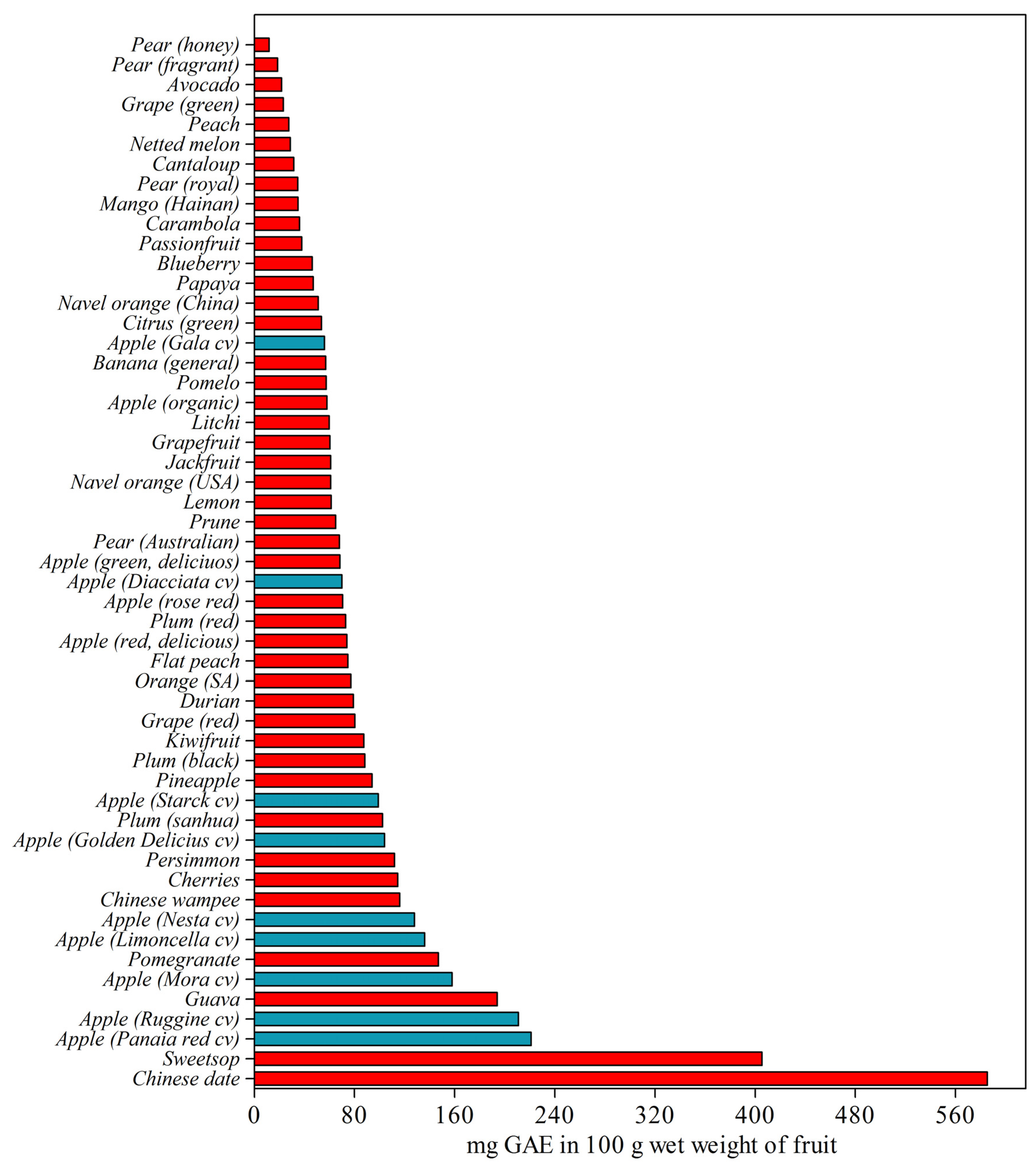
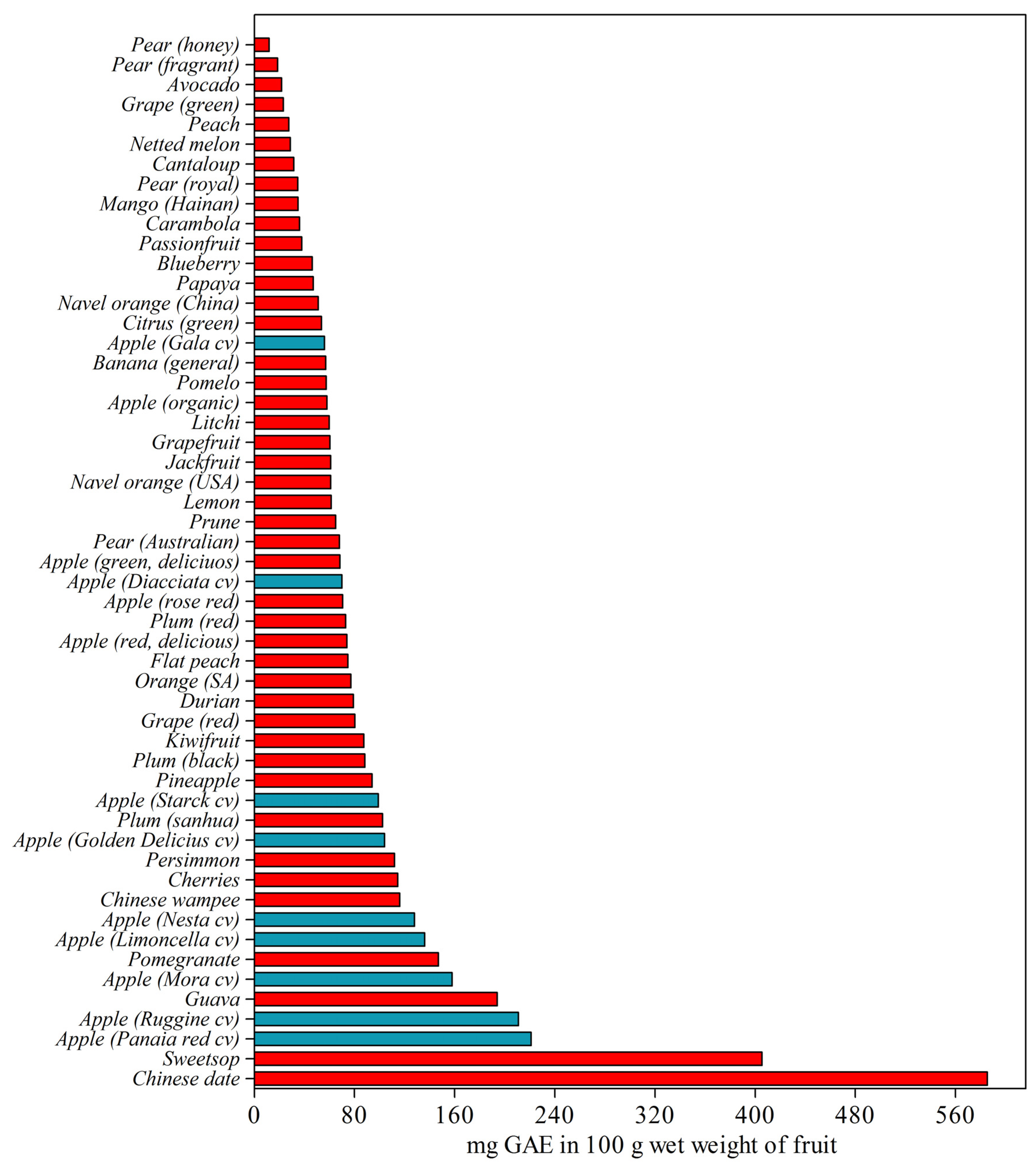
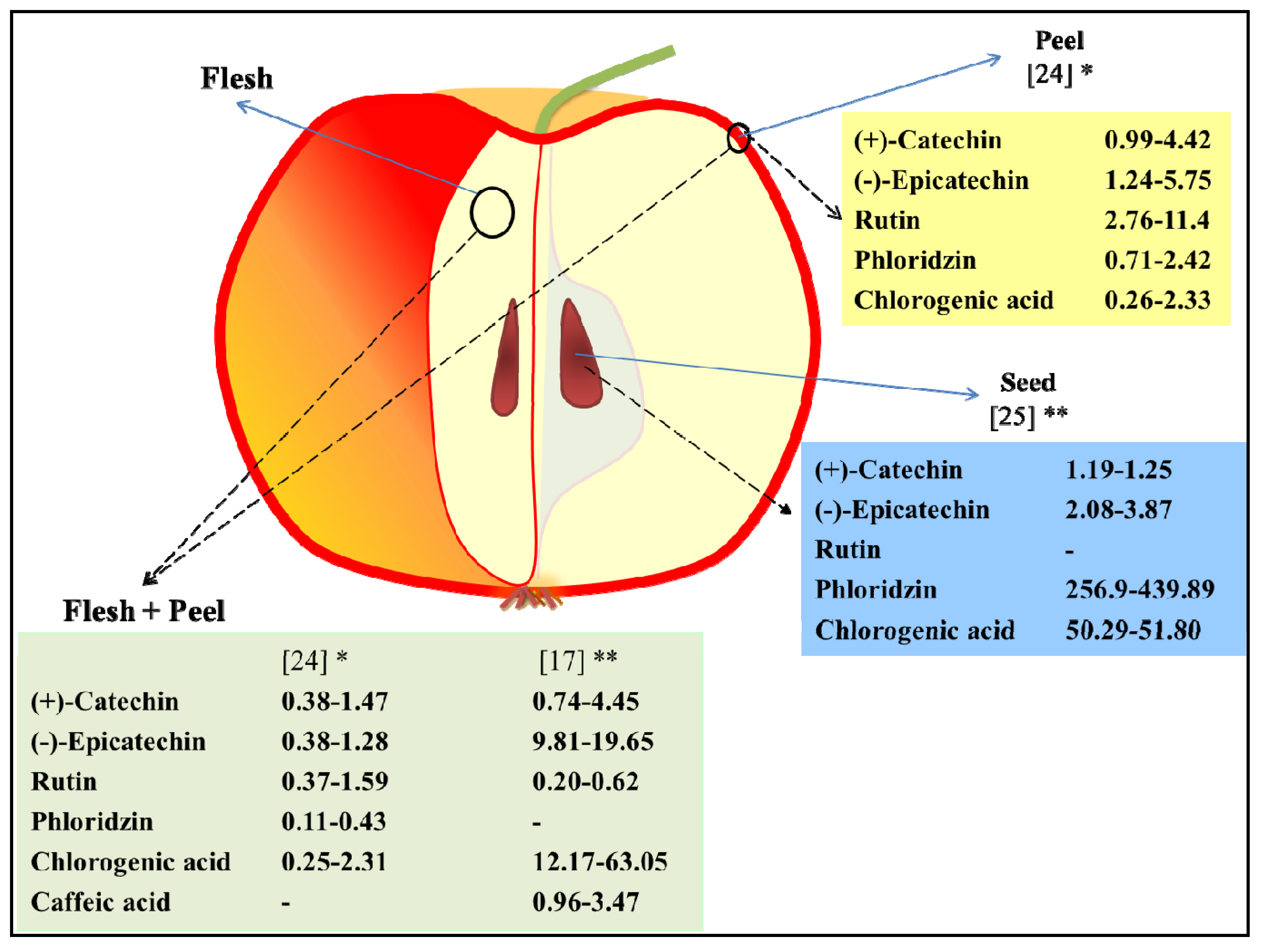
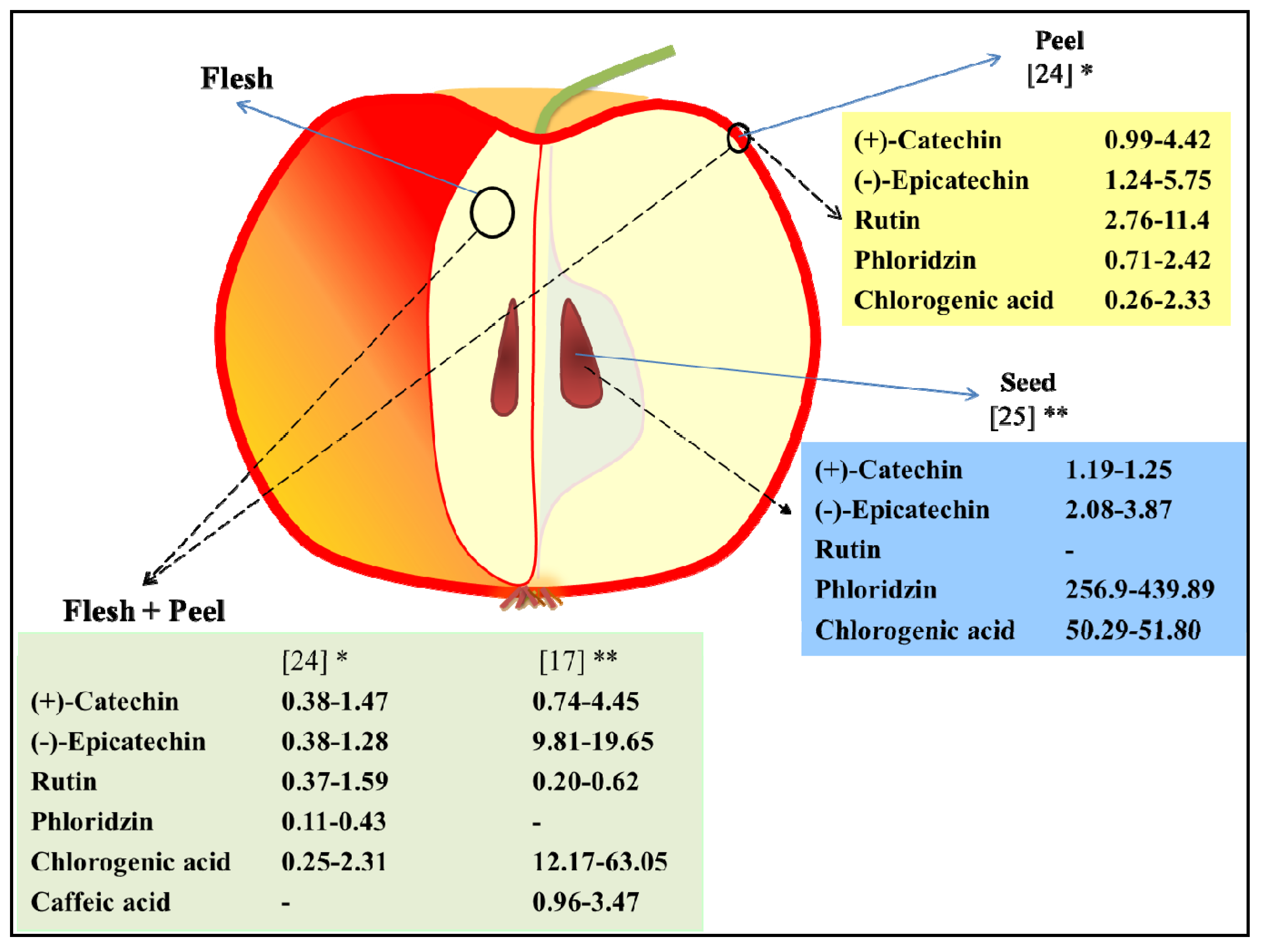
2.1. Phenolic Compounds in Apple
2.2. Health Benefits of Apple Phenolics
2.3. Stability of Phenolics in Apple Fruit during Postharvest and in Juice
2.4. Improving Phenolics Profile in Apple Fruit: From Germplasm Recovery and Classical Breeding to GM Plants
3. Conclusions
- literature on apple polyphenols properties suggest their potential use in preventing several chronic diseases in humans.
- Deeper knowledge on the specific molecular mechanisms of action (i.e., epigenetic modifications) of apple polyphenols in human disease is necessary.
- Phenolic compound characterization in whole apple fruit is well established, while their fate during transformation in juice need to be improved in order to better clarify the losses of these compounds and suitable strategies for their optimal conservation.
- The high variation on the localization of polyphenols composition in the fruit peel and flesh among cultivars and Malus species can be very useful for breeding program and underline the importance of germplasm conservation strategies.
- Genomic revolution and biotechnological applications will boost genetic improvement of elite apple genotypes by enabling the introduction of highly specific polyphenolic traits.
Conflicts of Interest
References
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant Polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Harborne, J.B. Biochemistry of Phenolic Compounds; Academic Press: London, UK, 1964; pp. 511–543. [Google Scholar]
- Bensath, A.; Ruysnyak, T.; Szent-Györgii, A. Vitamine nature of flavones. Nature 1936, 138, 789–793. [Google Scholar] [CrossRef]
- ScienceDirect Database. Available online: www.sciencedirect.com (accessed on 15 July 2013).
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Link, A.; Balaguer, F.; Goel, A. Cancer chemoprevention by dietary polyphenols: Promising role for epigenetics. Biochem. Pharmacol. 2010, 80, 1771–1792. [Google Scholar] [CrossRef]
- Van Duynhovena, J.; Vaughana, E.E.; Jacobsa, D.M.; Kempermana, R.A.; van Velzena, E.J.J.; Grossa, G.; Rogera, L.C.; Possemiersd, S.; Smildec, A.K.; Doréb, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 4531–4538. [Google Scholar] [CrossRef]
- Visioli, F.; Alarcón De La Lastra, C.; Andres-Lacueva, C.; Aviram, M.; Calhau, C.; Cassano, A.; D’Archivio, M.; Faria, A.; Favé, G.; Fogliano, V.; et al. Polyphenols and human health: A prospectus. Crit. Rev. Food Sci. Nut. 2011, 51, 524–546. [Google Scholar] [CrossRef]
- Frei, B.; England, L.; Ames, B.N. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Nat. Acad. Sci. USA 1989, 86, 6377–6638. [Google Scholar] [CrossRef]
- Lambert, J.D.; Elias, R.J. Antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef]
- Pan, M.H.; Lai, C.S.; Wu, J.C.; Ho, C.T. Epigenetic and diseases targets by polyphenols. Curr. Pharm. Des. 2013, in press. [Google Scholar]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Phytochemistry: Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Food and Agriculture Organization (FAO). Available online: http://faostat3.fao.org/home/index.html (accessed on 15 July 2013).
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Iacopini, P.; Camangi, F.; Stefani, A.; Sebastiani, L. Antiradical potential of ancient Italian apple varieties of Malus x domestica Borkh. In a peroxynitrite-induced oxidative process. J. Food Comp. Anal. 2010, 23, 518–524. [Google Scholar] [CrossRef]
- Minnocci, A.; Iacopini, P.; Martinelli, F.; Sebastiani, L. Micromorphological, biochemical, and genetic characterization of two ancient, late-bearing apple varieties. Eur. J. Hort. Sci. 2010, 75, 1–7. [Google Scholar]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073–2085. [Google Scholar]
- Cuthbertson, D.; Andrews, P.K.; Reganold, J.P.; Davies, N.M.; Lange, B.M. Utility of metabolomics toward assessing the metabolic basis of quality traits in apple fruit with an emphasis on antioxidants. J. Agric. Food Chem. 2012, 60, 8552–8560. [Google Scholar] [CrossRef] [PubMed]
- Vrhovsek, U.; Rigo, A.; Tonon, D.; Mattivi, F. Quantitation of polyphenols in different apple varieties. J. Agric. Food Chem. 2004, 52, 6532–6538. [Google Scholar] [CrossRef]
- McGhie, T.K.; Hunt, M.; Barnet, L.E. Cultivar and growing region determine the antioxidant polyphenolic concentration and composition of apples grown in New Zealand. J. Agric. Food Chem. 2005, 53, 3065–3070. [Google Scholar] [CrossRef]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef]
- Łata, B.; Trampczynska, A.; Paczesna, J. Cultivar variation in apple peel and whole fruit phenolic composition. Sci. Hort. 2009, 121, 176–181. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Tuszyński, T. Antioxidant activity of apples—An impact of maturity stage and fruit part. Sci. Pol. Technol. Aliment. 2011, 10, 443–454. [Google Scholar]
- Paluszczak, J.; Krajka-Kuzniak, V.; Baer-Dubowska, W. The effect of dietary polyphenols on the epigenetic regulation of gene expression in MCF7 breast cancer cells. Toxicol. Lett. 2010, 192, 119–125. [Google Scholar] [CrossRef]
- Otake, Y.; Nolan, A.L.; Walle, U.K.; Walle, T. Quercetin and resveratrol potently reduce estrogen sulfotransferase activity in normal human mammary epithelial cells. J. Steroid Biochem. Mol. Biol. 2000, 73, 265–270. [Google Scholar] [CrossRef]
- Marchetti, F.; de Santi, C.; Vietri, M.; Pietrabissa, A.; Spisni, R.; Mosca, F.; Pacifici, G.M. Differential inhibition of human liver and duodenum sulphotransferase activities by quercetin, a flavonoid present in vegetables, fruit and wine. Xenobiotica 2001, 31, 841–847. [Google Scholar] [CrossRef]
- Coughtrie, M.W.; Sharp, S.; Maxwell, K.; Innes, N.P. Biology and function of the reversible sulfation pathway catalysed by human sulfotransferases and sulfatases. Chem. Biol. Interact. 1998, 109, 3–27. [Google Scholar] [CrossRef]
- Gerhauser, C. Cancer chemopreventive potential of apples, apple juice, and apple components. Planta Med. 2008, 74, 1608–1624. [Google Scholar] [CrossRef]
- Chan, A.; Shea, T. Dietary supplementation with apple juice decreases endogenous amyloid-beta levels in murine brain. J. Alzheimer’s Dis. 2009, 16, 167–171. [Google Scholar]
- Johnston, K.; Clifford, M.; Morgan, L. Possible role for apple juice phenolic compounds in the acute modification of glucose tolerance and gastrointestinal hormone secretion in humans. J. Sci. Food Agric. 2002, 82, 1800–1805. [Google Scholar] [CrossRef]
- Marks, S.C.; Mullen, W.; Borges, G.; Crozier, A. Absorption, metabolism, and excretion of cider dihyrochalcones in healthy humans and subjects with an ileostomy. J. Agric. Food Chem. 2009, 57, 2009–2015. [Google Scholar] [CrossRef]
- Graziani, G.; D’Argenio, G.; Tuccillo, C.; Loguercio, C.; Ritieni, A.; Morisco, F.; del Vecchio, B.; Fogliano, V.; Romano, M. Apple phenol extracts prevent damage to human gastric epithelial cells in vitro and to rat gastric mucosa in vivo. Gut 2005, 54, 193–200. [Google Scholar] [CrossRef]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 12, 3–5. [Google Scholar]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Am. Soc. Nutr. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Viggiano, A.; Monda, M.; Turco, I.; Incarnato, L.; Vinno, V.; Viggiano, E.; Baccari, M.; de Luca, B. Annurca apple-rich diet restores long-term potentiation and induces behavioural modifications in aged rats. Exp. Neurol. 2006, 199, 354–361. [Google Scholar] [CrossRef]
- Gallus, S.; Talamini, R.; Giacosa, A.; Montella, M.; Ramazzotti, V.; Franceschi, S.; Negri, E.; La Vecchia, C. Does an apple a daykeep the oncologist away? Ann. Oncol. 2005, 16, 1841–1844. [Google Scholar] [CrossRef]
- Jedrychowski, W.; Maugeri, U.; Popiela, T.; Kulig, J.; Sochacka-Tatara, E.; Pac, A.; Sowa, A.; Musial, A. Case-control study on beneficial effect of regular consumption of apples on colorectal cancer risk in a population with relatively low intake of fruits and vegetables. Eur. J. Cancer Prev. 2010, 19, 42–47. [Google Scholar] [CrossRef]
- Serra, A.T.; Rocha, J.; Sepodes, B.; Matias, A.A.; Feliciano, R.P.; de Carvalho, A.; Bronze, M.R.; Duarte, C.M.; Figueira, M.E. Evaluation of cardiovascular protective effect of different apple varieties—Correlation of response with composition. Food Chem. 2012, 135, 2378–2386. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P.; Tuszyński, T. The profile of polyphenols and antioxidant properties of selected apple cultivars grown in Poland. J. Fruit Ornam. Plant Res. 2010, 18, 39–50. [Google Scholar]
- Carbone, K.; Giannini, B.; Picchi, V.; Scalzo, R.L.; Cecchini, F. Phenolic composition and free radical scavenging activity of different apple varieties in relation on the cultivar, tissue type and storage. Food Chem. 2011, 127, 493–500. [Google Scholar] [CrossRef]
- Begić-Akagić, A.; Spaho, N.; Oručević, S.; Drkenda, P.; Kurtović, M.; Gaši, F.; Kopjar, M.; Piližota, V. Influence of cultivar, storage time, and processing on the phenol content of cloudy apple juice. Croat. J. Food Sci. Technol. 2011, 3, 1–8. [Google Scholar]
- Matthes, A.; Schmitz-Eiberger, M. Polyphenol content and antioxidant capacity of apple fruit: Effect of cultivar and storage conditions. J. Appl. Bot. Food Qual. 2009, 82, 152–157. [Google Scholar]
- Napolitano, A.; Cascone, A.; Graziani, G.; Ferracane, R.; Scalfi, L.; di Vaio, C.; Ritieni, A.; Fogliano, V. Influence of variety and storage on the polyphenol composition of apple flesh. J. Agric. Food Chem. 2004, 52, 6526–6531. [Google Scholar] [CrossRef]
- Cheynier, V. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223–229. [Google Scholar]
- Van der Sluis, A.A.; Dekker, M.; van Boekel, M.A.J.S. Activity and concentration of polyphenolic antioxidants in apple juice. Stability during storage. J. Agric. Food Chem. 2005, 53, 1073–1080. [Google Scholar] [CrossRef]
- Markowski, J.; Płocharski, W. Determination of phenolic compounds in apples and processed apple products. J. Fruit Ornam. Plant Res. 2006, 14, 133–142. [Google Scholar]
- Lu, Y.; Foo, L.Y. Antioxidant and radical scavenging activities of polyphenols from apple pomace. Food Chem. 2000, 68, 81–85. [Google Scholar] [CrossRef]
- Spanos, G.A.; Wrolstad, R.E.; Heatherbell, D.A. Influence of processing and storage on the phenolic composition of apple juice. J. Agric. Food Chem. 1990, 38, 1572–1579. [Google Scholar] [CrossRef]
- Miller, N.J.; Diplock, A.T.; Rice-Evans, C.A. Evaluation of the total antioxidant activity as a marker of the deterioration of apple juice on storage. J. Agric. Food Chem. 1995, 43, 1794–1801. [Google Scholar] [CrossRef]
- Volz, R.K.; McGhie, T.K. Genetic variability in apple fruit polyphenol composition in Malus x domestica and Malus sieversii germplasm grown in New Zealand. J. Agric. Food Chem. 2011, 59, 11509–11521. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus x domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Troggio, M.; Gleave, A.; Salvi, S.; Chagné, D.; Cestaro, A.; Kumar, S.; Crowhurst, R.N.; Gardiner, S.E. Apple, from genome to breeding. Tree Genet. Gen. 2012, 8, 509–529. [Google Scholar] [CrossRef]
- Volz, R.; Oraguzie, N.; Whitworth, C.; How, N.; Chagné, D.; Carlisle, C.; Gardiner, S. Red flesh breeding in apple-progress and challenges. Acta Hort. 2009, 814, 337–342. [Google Scholar]
- Espley, R.V.; Brendolise, C.; Chagne, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef]
- Espley, R.V.; Bovy, A.; Bava, C.; Jaeger, S.R.; Tomes, S.; Norling, C.; Crawford, J.; Rowan, D.; McGhie, T.K.; Brendolise, C.; et al. Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer attributes. Plant Biotechnol. J. 2013, 11, 408–419. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Francini, A.; Sebastiani, L. Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing. Antioxidants 2013, 2, 181-193. https://doi.org/10.3390/antiox2030181
Francini A, Sebastiani L. Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing. Antioxidants. 2013; 2(3):181-193. https://doi.org/10.3390/antiox2030181
Chicago/Turabian StyleFrancini, Alessandra, and Luca Sebastiani. 2013. "Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing" Antioxidants 2, no. 3: 181-193. https://doi.org/10.3390/antiox2030181
APA StyleFrancini, A., & Sebastiani, L. (2013). Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing. Antioxidants, 2(3), 181-193. https://doi.org/10.3390/antiox2030181