
Effect of Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds and Antioxidant Activity of Fermented Aloe vera Juices
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
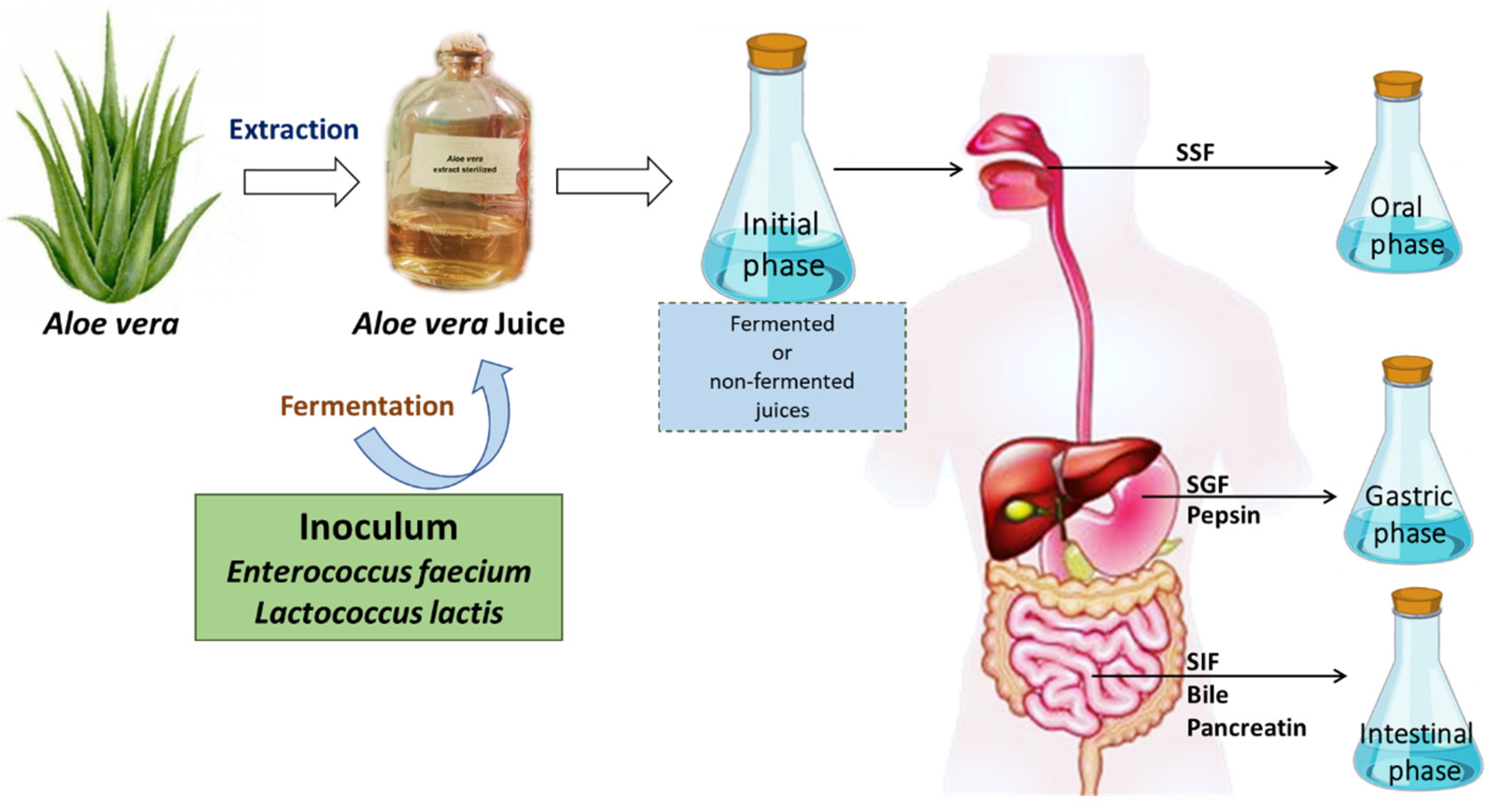
2.2. Juice Preparation and Fermentation
2.3. Procedure for In Vitro Digestion
2.4. Total Phenolic Content Analysis
2.5. Identification of Individual Phenolic Compounds by Ultra-Performance Liquid Chromatography (UPLC-DAD)
2.6. Antioxidant Activity
2.6.1. Ferric Reducing Antioxidant Power Assay (FRAP)
2.6.2. ABTS Radical Cation Scavenging Activity
2.6.3. DPPH Radical Scavenging Activity
2.7. Determination of Bioaccessibility
2.8. Statistical Analysis
3. Results
3.1. Effect of In Vitro Digestion on Phenolic Compounds
3.1.1. Total Phenolic Content (TPC)
3.1.2. Phenolic Compounds of Non-Fermented Aloe vera Juice (Av)
3.1.3. Phenolic Compounds of Aloe vera Juice Fermentation with E. faecium (AvF)
3.1.4. Phenolic Compounds of Aloe vera Juice Fermentation with L. lactis (AvL)
3.2. Bioaccessibility of Biocompounds from Aloe vera Juice
3.3. Antioxidant Activity
3.4. Correlation Analysis
4. Discussion
4.1. Effect of In Vitro Digestion on Phenolic Compounds
4.2. Bioactive Compounds Identified in Aloe vera Juice
4.3. Bioaccessibility of Biocompounds from Aloe vera Juice
4.4. Antioxidant Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, A.; Amaro, A.L.; Pintado, M. Impact of Food Matrix Components on Nutritional and Functional Properties of Fruit-Based Products. Curr. Opin. Food Sci. 2018, 22, 153–159. [Google Scholar] [CrossRef]
- Mantzourani, I.; Terpou, A.; Bekatorou, A.; Mallouchos, A.; Alexopoulos, A.; Kimbaris, A.; Bezirtzoglou, E.; Koutinas, A.A.; Plessas, S. Functional Pomegranate Beverage Production by Fermentation with a Novel Synbiotic L. Paracasei Biocatalyst. Food Chem. 2020, 308, 125658. [Google Scholar] [CrossRef] [PubMed]
- Valero-Cases, E.; Cerdá-Bernad, D.; Pastor, J.J.; Frutos, M.J. Non-Dairy Fermented Beverages as Potential Carriers to Ensure Probiotics, Prebiotics, and Bioactive Compounds Arrival to the Gut and Their Health Benefits. Nutrients 2020, 12, 1666. [Google Scholar] [CrossRef] [PubMed]
- Cuvas-Limon, R.B.; Nobre, C.; Cruz, M.; Rodriguez-Jasso, R.M.; Ruíz, H.A.; Loredo-Treviño, A.; Texeira, J.A.; Belmares, R. Spontaneously Fermented Traditional Beverages as a Source of Bioactive Compounds: An Overview. Crit. Rev. Food Sci. Nutr. 2020, 61, 2984–3006. [Google Scholar] [CrossRef] [PubMed]
- Cicenia, A.; Scirocco, A.; Carabotti, M.; Pallotta, L.; Marignani, M.; Severi, C. Postbiotic Activities of Lactobacilli-Derived Factors. J. Clin. Gastroenterol. 2014, 48, S18–S22. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, L.; Liu, Y.; Zhang, Q.; Li, Y.; Wu, Z. Release of Phenolics Compounds from Rubus Idaeus L. Dried Fruits and Seeds during Simulated In vitro Digestion and Their Bio-Activities. J. Funct. Foods 2018, 46, 57–65. [Google Scholar] [CrossRef]
- Rasera, G.B.; de Camargo, A.C.; de Castro, R.J.S. Bioaccessibility of Phenolic Compounds Using the Standardized INFOGEST Protocol: A Narrative Review. Compr. Rev. Food Sci. Food Saf. 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Morais, S.G.G.; da Silva Campelo Borges, G.; dos Santos Lima, M.; Martín-Belloso, O.; Magnani, M. Effects of Probiotics on the Content and Bioaccessibility of Phenolic Compounds in Red Pitaya Pulp. Food Res. Int. 2019, 126, 108681. [Google Scholar] [CrossRef]
- Gonçalves, S.; Moreira, E.; Andrade, P.B.; Valentão, P.; Romano, A. Effect of In vitro Gastrointestinal Digestion on the Total Phenolic Contents and Antioxidant Activity of Wild Mediterranean Edible Plant Extracts. Eur. Food Res. Technol. 2019, 245, 753–762. [Google Scholar] [CrossRef]
- Žuntar, I.; Petric, Z.; Bursa’c, D.; Kovačevi’c, B.K.; Putnik, P. Safety of Probiotics: Functional Fruit Beverages and Nutraceuticals. Foods 2020, 9, 947. [Google Scholar] [CrossRef]
- Bagci, U.; Ozmen Togay, S.; Temiz, A.; Ay, M. Probiotic Characteristics of Bacteriocin-Producing Enterococcus Faecium Strains Isolated from Human Milk and Colostrum. Folia Microbiol. 2019, 64, 735–750. [Google Scholar] [CrossRef]
- Choeisoongnern, T.; Sirilun, S.; Waditee-Sirisattha, R.; Pintha, K.; Peerajan, S.; Chaiyasut, C. Potential Probiotic Enterococcus Faecium OV3-6 and Its Bioactive Peptide as Alternative Bio-Preservation. Foods 2021, 10, 2264. [Google Scholar] [CrossRef]
- Huang, J.; Huang, J.; Yin, T.; Lv, H.; Zhang, P.; Li, H. Enterococcus Faecium R0026 Combined with Bacillus Subtilis R0179 Prevent Obesity-Associated Hyperlipidemia and Modulate Gut Microbiota in C57BL/6 Mice. J. Microbiol. Biotechnol. 2021, 31, 181–188. [Google Scholar] [CrossRef]
- Bandyopadhyay, B.; Das, S.; Kumar Mitra, P.; Kundu, A.; Mandal, V.; Adhikary, R.; Chandra Mandal, N. Characterization of Two New Strains of Lactococcus Lactis for Their Probiotic Efficacy over Commercial Synbiotics Consortia. Brazilian J. Microbiol. 2022, 53, 903–920. [Google Scholar] [CrossRef]
- Kondrotiene, K.; Lauciene, L.; Andruleviciute, V.; Kasetiene, N.; Serniene, L.; Sekmokiene, D.; Malakauskas, M. Safety Assessment and Preliminary In vitro Evaluation of Probiotic Potential of Lactococcus Lactis Strains Naturally Present in Raw and Fermented Milk. Curr. Microbiol. 2020, 77, 3013–3023. [Google Scholar] [CrossRef]
- Fernandes Pereira, A.L.; Rodrigues, S. Turning Fruit Juice Into Probiotic Beverages. In Fruit Juices: Extraction, Composition, Quality and Analysis; Elsevier Inc.: Cambridge, UK, 2018; pp. 279–287. [Google Scholar] [CrossRef]
- Cuvas-Limón, R.B.; Julio, M.S.; Carlos, C.E.J.; Mario, C.H.; Mussatto, S.I.; Ruth, B.C. Aloe Vera and Probiotics: A New Alternative to Symbiotic Functional Foods. Annu. Res. Rev. Biol. 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Cuvas-Limón, R.B.; Ferreira-Santos, P.; Cruz, M.; Teixeira, J.A.; Belmares, R.; Nobre, C. Novel Bio-Functional Aloe Vera Beverages Fermented by Probiotic Enterococcus Faecium and Lactobacillus Lactis. Molecules 2022, 27, 2473. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, D.; Shi, J. Chemical Constituents, Biological Activity and Agricultural Cultivation of Aloe Vera. Asian J. Chem. 2013, 25, 6477–6485. [Google Scholar] [CrossRef]
- Baruah, A.; Bordoloi, M.; Deka Baruah, H.P. Aloe Vera: A Multipurpose Industrial Crop. Ind. Crops Prod. 2016, 94, 951–963. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST Static In vitro Simulation of Gastrointestinal Food Digestion. Nat. Protoc. 2019, 1, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Nobre, C.; González, A.; Losoya, C.; Teixeira, J.A.; Belmares, R.; Abrunhosa, L. Detoxification of Ochratoxin A and Zearalenone by Pleurotus Ostreatus during In vitro Gastrointestinal Digestion. Food Chem. 2022, 384, 132525. [Google Scholar] [CrossRef]
- González, A.; Nobre, C.; Simões, L.S.; Cruz, M.; Loredo, A.; Rodríguez-Jasso, R.M.; Contreras, J.; Texeira, J.; Belmares, R. Evaluation of Functional and Nutritional Potential of a Protein Concentrate from Pleurotus Ostreatus Mushroom. Food Chem. 2021, 346, 128884. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Santos, P.; Genisheva, Z.; Pereira, R.N.; Teixeira, J.A.; Rocha, C.M.R. Moderate Electric Fields as a Potential Tool for Sustainable Recovery of Phenolic Compounds from Pinus Pinaster Bark. ACS Sustain. Chem. Eng. 2019, 7, 8816–8826. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, L.S.; Martins, J.T.; Pinheiro, A.C.; Vicente, A.A.; Ramos, O.L. β-Lactoglobulin Micro- and Nanostructures as Bioactive Compounds Vehicle: In Vitro Studies. Food Res. Int. 2020, 131, 108979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mármol, I.; Quero, J.; Ibarz, R.; Ferreira-Santos, P.; Teixeira, J.A.; Rocha, C.M.R.; Pérez-Fernández, M.; García-Juiz, S.; Osada, J.; Martín-Belloso, O.; et al. Valorization of Agro-Food by-Products and Their Potential Therapeutic Applications. Food Bioprod. Process. 2021, 128, 247–258. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Ibarz, R.; Fernandes, J.-M.; Pinheiro, A.C.; Botelho, C.; Rocha, C.M.R.; Teixeira, J.A.; Martín-Belloso, O. Encapsulated Pine Bark Polyphenolic Extract during Gastrointestinal Digestion: Bioaccessibility, Bioactivity and Oxidative Stress Prevention. Foods 2021, 10, 328. [Google Scholar] [CrossRef]
- Xiang, Z.; Deng, J.; Yang, K.; Zhu, Y.; Xia, C.; Chen, J.; Liu, T. Effect of Processing on the Release of Phenolic Compounds and Antioxidant Activity during In vitro Digestion of Hulless Barley. Arab. J. Chem. 2021, 14, 103447. [Google Scholar] [CrossRef]
- Duan, L.; Ding, W.; Liu, X.; Cheng, X.; Cai, J.; Hua, E.; Jiang, H. Biosynthesis and Engineering of Kaempferol in Saccharomyces Cerevisiae. Microb. Cell Fact. 2017, 16, 165. [Google Scholar] [CrossRef]
- Marchese, A.; Coppo, E.; Sobolev, A.P.; Rossi, D.; Mannina, L.; Daglia, M. Influence of In vitro Simulated Gastroduodenal Digestion on the Antibacterial Activity, Metabolic Profiling and Polyphenols Content of Green Tea (Camellia sinensis). Food Res. Int. 2014, 63, 182–191. [Google Scholar] [CrossRef]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Ritieni, A. In Vitro Bioaccessibility and Antioxidant Activity of Coffee Silverskin Polyphenolic Extract and Characterization of Bioactive Compounds Using UHPLC-Q-Orbitrap HRMS. Molecules 2020, 25, 2132. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Santos, P.; Genisheva, Z.; Botelho, C.; Rocha, C.; António Teixeira, J. Valorization of Natural Antioxidants for Nutritional and Health Applications. In Antioxidants—Benefits, Sources, Mechanisms of Action; IntechOpen Limited: London, UK, 2021; ISBN 978-1-83968-865-2. [Google Scholar]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Phenolic Content and Antioxidant Activities of Selected Potato Varieties and Their Processing By-Products. J. Funct. Foods 2013, 5, 590–600. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, J.; Wei, Z.; Shahidi, F. Effect of In vitro Digestion on Phenolics and Antioxidant Activity of Red and Yellow Colored Pea Hulls. Food Chem. 2021, 337, 127606. [Google Scholar] [CrossRef] [PubMed]
- Gullon, B.; Pintado, M.E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. In Vitro Gastrointestinal Digestion of Pomegranate Peel (Punica granatum) Flour Obtained from Co-Products: Changes in the Antioxidant Potential and Bioactive Compounds Stability. J. Funct. Foods 2015, 19, 617–628. [Google Scholar] [CrossRef]
- Lucas-Gonzalez, R.; Navarro-Coves, S.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Assessment of Polyphenolic Profile Stability and Changes in the Antioxidant Potential of Maqui Berry (Aristotelia Chilensis (Molina) Stuntz) during In vitro Gastrointestinal Digestion. Ind. Crops Prod. 2016, 94, 774–782. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of Bioactive Food Compounds: A Challenging Journey to Bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J.; Rubió, L. Application of In vitro Gastrointestinal Digestion and Colonic Fermentation Models to Pomegranate Products (Juice, Pulp and Peel Extract) to Study the Stability and Catabolism of Phenolic Compounds. J. Funct. Foods 2015, 14, 529–540. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The Effects of Polyphenols and Other Bioactives on Human Health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Bohn, T.; McDougall, G.J.; Alegría, A.; Alminger, M.; Arrigoni, E.; Aura, A.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the Gap—Deficits in Our Knowledge of Aspects Impacting the Bioavailability of Phytochemicals and Their Metabolites—A Position Paper Focusing on Carotenoids and Polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef]
- Bohn, T. Dietary Factors Affecting Polyphenol Bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J. Stability and Metabolism of Arbutus Unedo Bioactive Compounds (Phenolics and Antioxidants) under In vitro Digestion and Colonic Fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Deng, J.; Wen, L.; You, L.; Zhao, Z.; Zhou, L. Release of Phenolic Compounds and Antioxidant Capacity of Chinese Hawthorn “Crataegus Pinnatifida” during In vitro Digestion. J. Funct. Foods 2018, 40, 76–85. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Soszka, A. Changes in Phenolic Compounds and Antioxidant Activity of Fruit Musts and Fruit Wines during Simulated Digestion. Molecules 2020, 25, 5574. [Google Scholar] [CrossRef] [PubMed]
- Gullon, B.; Pintado, M.E.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Assessment of Polyphenolic Profile and Antibacterial Activity of Pomegranate Peel (Punica Granatum) Flour Obtained from Co-Product of Juice Extraction. Food Control 2016, 59, 94–98. [Google Scholar] [CrossRef]
- Karakaya, S. Bioavailability of Phenolic Compounds. Crit. Rev. Food Sci. Nutr. 2004, 44, 453–464. [Google Scholar] [CrossRef]
- Friedman, M.; Jürgens, H.S. Effect of PH on the Stability of Plant Phenolic Compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef]
- Khochapong, W.; Ketnawa, S.; Ogawa, Y.; Punbusayakul, N. Effect of In vitro Digestion on Bioactive Compounds, Antioxidant and Antimicrobial Activities of Coffee (Coffea arabica L.) Pulp Aqueous Extract. Food Chem. 2021, 348, 129094. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, B.; Xia, Y.; Li, H.; Shi, X.; Wang, J.; Deng, Z. Bioaccessibility and Transformation Pathways of Phenolic Compounds in Processed Mulberry (Morus alba L.) Leaves after In vitro Gastrointestinal Digestion and Faecal Fermentation. J. Funct. Foods 2019, 60, 103406. [Google Scholar] [CrossRef]
- Altunkaya, A.; Gökmen, V.; Skibsted, L.H. PH Dependent Antioxidant Activity of Lettuce (L. Sativa) and Synergism with Added Phenolic Antioxidants. Food Chem. 2016, 190, 25–32. [Google Scholar] [CrossRef]
- Filannino, P.; Bai, Y.; Di Cagno, R.; Gobbetti, M.; Gänzle, M.G. Metabolism of Phenolic Compounds by Lactobacillus Spp. during Fermentation of Cherry Juice and Broccoli Puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of Fermentation with Different Lactic Acid Bacteria and In vitro Digestion on the Biotransformation of Phenolic Compounds in Fermented Pomegranate Juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Ellagitannins, Ellagic Acid and Their Derived Metabolites: A Review about Source, Metabolism, Functions and Health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Kilic, I.; Yeşiloǧlu, Y.; Bayrak, Y. Spectroscopic Studies on the Antioxidant Activity of Ellagic Acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 130, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.T.; Chou, C.T.; Su, N.W. A Food-Grade Self-Nanoemulsifying Delivery System for Enhancing Oral Bioavailability of Ellagic Acid. J. Funct. Foods 2017, 34, 207–215. [Google Scholar] [CrossRef]
- Shim, S.M.I.; Kwon, H. Assessing Absorbability of Bioactive Components in Aloe Using In Vitro Digestion Model with Human Intestinal Cell. J. Food Biochem. 2010, 34, 425–438. [Google Scholar] [CrossRef]
- Radha, M.H.; Laxmipriya, N.P. Evaluation of Biological Properties and Clinical Effectiveness of Aloe Vera: A Systematic Review. J. Tradit. Complement. Med. 2015, 5, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, R.F.S.; Martins, J.T.; Duarte, C.M.M.; Vicente, A.A.; Pinheiro, A.C. Advances in Nutraceutical Delivery Systems: From Formulation Design for Bioavailability Enhancement to Efficacy and Safety Evaluation. Trends Food Sci. Technol. 2018, 78, 270–291. [Google Scholar] [CrossRef] [Green Version]
- Ting, Y.; Jiang, Y.; Ho, C.T.; Huang, Q. Common Delivery Systems for Enhancing in Vivo Bioavailability and Biological Efficacy of Nutraceuticals. J. Funct. Foods 2014, 7, 112–128. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Vázquez-Sánchez, K.; López-Barrera, D.; Loarca-Piña, G.; Mendoza-Díaz, S.; Oomah, B.D. Simulated Gastrointestinal Digestion and In vitro Colonic Fermentation of Spent Coffee (Coffea arabica L.): Bioaccessibility and Intestinal Permeability. Food Res. Int. 2015, 77, 156–161. [Google Scholar] [CrossRef]
- Barros, R.G.C.; Pereira, U.C.; Andrade, J.K.S.; de Oliveira, C.S.; Vasconcelos, S.V.; Narain, N. In Vitro Gastrointestinal Digestion and Probiotics Fermentation Impact on Bioaccessbility of Phenolics Compounds and Antioxidant Capacity of Some Native and Exotic Fruit Residues with Potential Antidiabetic Effects. Food Res. Int. 2020, 136, 109614. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, H.; Liu, H.; Ma, R.; Ma, J.; Fang, H. Fermentation by Multiple Bacterial Strains Improves the Production of Bioactive Compounds and Antioxidant Activity of Goji Juice. Molecules 2019, 24, 3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebo, O.A.; Medina-Meza, I.G. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the Total Antioxidant Capacity and Total Polyphenol Content of 23 Commercially Available Vegetable Juices before and after In vitro Digestion Measured by FRAP, DPPH, ABTS and Folin-Ciocalteu Methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total Phenolics, Flavonoids, Anthocyanins and Antioxidant Activity Following Simulated Gastro-Intestinal Digestion and Dialysis of Apple Varieties: Bioaccessibility and Potential Uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In Vitro Bio-Accessibility and Antioxidant Activity of Grape Polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and Pitfalls in Measuring Antioxidant Efficacy: A Critical Evaluation of ABTS, DPPH, and ORAC Assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Vilas-Boas, A.A.; Oliveira, A.; Jesus, D.; Rodrigues, C.; Figueira, C.; Gomes, A.; Pintado, M. Chlorogenic Acids Composition and the Impact of In vitro Gastrointestinal Digestion on Espresso Coffee from Single-Dose Capsule. Food Res. Int. 2020, 134, 109223. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSF (mmol/L) | SGF (mmol/L) | SIF (mmol/L) | |
|---|---|---|---|
| KCl | 15.10 | 6.90 | 6.80 |
| KH2PO4 | 3.70 | 0.90 | 0.80 |
| NaHCO3 | 13.60 | 25.00 | 85.00 |
| MgCl2(H2O)6 | 0.15 | 0.12 | 0.33 |
| (NH4)2CO3 | 0.06 | 0.50 | - |
| NaCl | - | 47.20 | 38.40 |
| HCl | - | 15.60 | 8.40 |
| Compounds (mg/L) | Initial Phase (Undigested) | Oral Phase | Gastric Phase | Intestinal Phase | Bioaccessibility (%) |
|---|---|---|---|---|---|
| Vanillic acid | 1.87 ± 0.10 a | 1.30 ± 0.14 a | n.d. | n.d. | n.d. |
| Catechin | 1.19 ± 0.18 a | n.d. | n.d. | n.d. | n.d. |
| Epicatechin | 30.99 ± 2.95 a | 27.70 ± 1.38 a | 30.80 ± 4.24 a | 16.32 ± 0.41 b | 55.30 ± 1.2 |
| p-Coumaric acid | 3.30 ± 0.89 a | 3.61 ± 0.19 a | 2.41 ± 0.39 a | 0.09 ± 0.007 b | 2.62 ± 0.4 |
| Ellagic acid | 8.20 ± 1.31 a | 8.90 ± 0.68 a | 9.24 ± 0.37 a | 14.18 ± 0.03 b | 194.56 ± 0.2 |
| Naringenin | 10.49 ± 0.87 a | 9.95 ± 0.35 a | 6.46 ± 1.40 b | 2.27 ± 0.06 c | 22.01 ± 0.5 |
| Hesperidin | 17.01 ± 2.46 ab | 18.36 ± 0.85 a | 14.46 ± 1.77 b | 7.63 ± 0.32 c | 48.91 ± 0.9 |
| Resveratrol | 1.93 ± 0.07 a | 2.05 ± 0.07 a | 2.44 ± 0.14 a | 1.35 ± 0.01 a | 71.59 ± 0.0 |
| Ferulic acid | 4.43 ± 0.81 a | 3.59 ± 1.50 ab | 1.12 ± 0.50 b | 1.27 ± 0.01 b | 32.67 ± 0.3 |
| Quercetin | 0.75 ± 0.12 a | 4.27 ± 0.60 b | 3.01 ± 0.51 b | 1.24 ± 0.02 c | 160.45 ± 0.1 |
| 3,4-Dihydroxybenzoic acid | 0.39 ± 0.18 a | 0.24 ± 0.02 a | 0.24 ± 0.04 a | n.d. | n.d. |
| Taxifolin | 7.53 ± 0.74 a | 6.88 ± 0.27 a | 6.69 ± 1.14 a | 1.78 ± 0.01 b | 25.12 ± 0.3 |
| Aloin | 14.10 ± 0.78 a | 13.47 ± 0.66 a | 14.86 ± 0.80 a | 3.40 ± 0.06 b | 25.03 ± 0.2 |
| Kaempferol | 1.53 ± 0.06 a | 1.40 ± 0.13 a | 1.36 ± 0.09 a | 6.42 ± 0.18 b | 430.24 ± 1.0 |
| TOTAL | 103.61 ± 11.51 | 101.71 ± 6.86 | 93.38 ± 11.38 | 55.95 ± 1.20 | -- |
| Compounds (mg/L) | Initial Phase (Undigested) | Oral Phase | Gastric Phase | Intestinal Phase | Bioaccessibility (%) |
|---|---|---|---|---|---|
| Vanillic acid | 2.61 ± 0.02 a | n.d. | n.d. | n.d. | n.d. |
| Epicatechin | 35.29 ± 1.18 a | 22.97 ± 0.38 b | 7.61 ± 3.14 c | 17.52 ± 0.01 d | 49.7 ± 0.6 |
| p-Coumaric acid | 0.81 ± 0.02 a | n.d. | n.d. | n.d. | n.d. |
| Ellagic acid | 15.09 ± 0.23 a | 22.03 ± 0.48 b | 30.66 ± 0.43 c | 62.00 ± 1.53 d | 411.2 ± 0.9 |
| Naringenin | 4.44 ± 0.39 a | 6.0 ± 2.65 a | 3.52 ± 0.48 a | 2.51 ± 0.06 a | 57.1 ± 0.2 |
| Hesperidin | 10.03 ± 0.07 a | 13.10 ± 4.05 a | 14.35 ± 4.06 a | 11.25 ± 0.39 a | 112.2 ± 0.2 |
| Resveratrol | 3.27 ± 0.12 a | 4.13 ± 0.06 a | 5.34 ± 0.21 a | 8.23 ± 0.01 b | 251.7 ± 0.1 |
| Ferulic Acid | 6.01 ± 3.53 a | 3.42 ± 0.98 ab | 1.78 ± 0.23 b | 3.43 ± 0.24 ab | 83.3 ± 1.3 |
| Quercetin | 3.38 ± 1.96 a | 5.04 ± 0.22 a | 6.16 ± 0.47 a | n.d. | n.d. |
| 3,4-Dihydroxybenzoic acid | 0.25 ± 0.01 a | n.d. | n.d. | n.d. | n.d. |
| Taxifolin | 4.25 ± 1.84 a | 5.18 ± 0.63 a | 3.06 ± 0.27 a | 4.88 ± 0.33 a | 137.2 ± 0.3 |
| Aloin | 15.49 ± 0.90 a | 13.61 ± 0.69 a | 13.84 ± 0.67 a | 4.46 ± 0.01 b | 28.9 ± 0.5 |
| Kaempferol | 1.88 ± 0.06 a | 2.73 ± 0.09 ab | 3.45 ± 0.04 b | 13.18 ± 2.32 c | 699.1 ± 1.1 |
| TOTAL | 102 ± 10 | 98 ± 10 | 89 ± 10 | 127 ± 4 | -- |
| Compounds (mg/L) | Initial Phase (Undigested) | Oral Phase | Gastric Phase | Intestinal Phase | Bioaccessibility (%) |
|---|---|---|---|---|---|
| Epicatechin | 48.18 ± 2.19 a | 22.55 ± 3.72 b | 4.91 ± 0.72 c | 8.08 ± 2.86 c | 16.54 ± 1.4 |
| p-Coumaric acid | 0.38 ± 0.07 a | n.d. | n.d. | n.d. | n.d. |
| Ellagic acid | 15.39 ± 0.89 a | 21.97 ± 0.64 b | 30.30 ± 0.05 c | 61.31 ± 3.71d | 398.21 ± 1.9 |
| Naringenin | 6.27 ± 0.04 a | 2.93 ± 1.06 b | 2.49 ± 1.13 b | 2.93 ± 0.21 b | 46.70 ± 0.1 |
| Hesperidin | 5.26 ± 0.72 a | 8.31 ± 1.76 ab | 5.28 ± 0.13 a | 10.03 ± 0.17 b | 193.77 ± 0.1 |
| Resveratrol | 4.01 ± 0.14 a | 4.44 ± 0.22 a | 5.14 ± 0.60 a | 8.07 ± 0.01 b | 201.40 ± 0.0 |
| Ferulic acid | 3.45 ± 0.07 a | 2.50 ± 0.28 a | 2.13 ± 0.44 a | 3.41 ± 0.13 a | 98.77 ± 0.0 |
| Quercetin | 1.13 ± 0.09 a | 3.07 ± 0.78 b | 4.96 ± 1.27 b | n.d. | n.d. |
| 3,4-Dihydroxybenzoic acid | 0.24 ± 0.08 a | n.d. | n.d. | n.d. | n.d. |
| Taxifolin | 7.60 ± 0.60 a | 4.46 ± 0.74 b | 3.39 ± 0.22 b | 4.67 ± 0.0 b | 61.88 ± 0.0 |
| Aloin | 20.82 ± 2.33 a | 16.95 ± 2.11 ab | 16.89 ± 0.0 ab | 15.59 ± 0.69 b | 76.22 ± 0.3 |
| Kaempferol | 2.07 ± 0.10 a | 2.68 ± 0.17 a | 3.51 ± 0.0 a | 11.86 ± 0.31 b | 575.33 ± 0.2 |
| TOTAL | 114 ± 7 | 89 ± 11 | 78 ± 4 | 125 ± 8 | -- |
| TPC | FRAP | ABTS | DPPH |
|---|---|---|---|
| Unfermented | 0.9657 * | 0.9843 * | −0.3807 ns |
| Fermented with E. faecium | 0.9246 ns | 0.9359 ns | −0.0181 ns |
| Fermented with L. lactis | 0.9909 ** | 0.9934 ** | 0.2894 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuvas-Limon, R.B.; Ferreira-Santos, P.; Cruz, M.; Teixeira, J.A.; Belmares, R.; Nobre, C. Effect of Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds and Antioxidant Activity of Fermented Aloe vera Juices. Antioxidants 2022, 11, 2479. https://doi.org/10.3390/antiox11122479
Cuvas-Limon RB, Ferreira-Santos P, Cruz M, Teixeira JA, Belmares R, Nobre C. Effect of Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds and Antioxidant Activity of Fermented Aloe vera Juices. Antioxidants. 2022; 11(12):2479. https://doi.org/10.3390/antiox11122479
Chicago/Turabian StyleCuvas-Limon, Ruth B., Pedro Ferreira-Santos, Mario Cruz, José A. Teixeira, Ruth Belmares, and Clarisse Nobre. 2022. "Effect of Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds and Antioxidant Activity of Fermented Aloe vera Juices" Antioxidants 11, no. 12: 2479. https://doi.org/10.3390/antiox11122479