
In Vitro Expanded Bioaccessibility of Quercetin-3-Rutinoside and Quercetin Aglycone from Buckwheat Biscuits Formulated from Flours Fermented by Lactic Acid Bacteria


Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Fermentation of Buckwheat Flours by LAB
2.3. Preparation of BBs from Fermented Flours
2.4. In Vitro Digestion of BBs
2.5. Extraction and Determination of Rutin and Quercetin by HPLC
2.6. Determination of the Expanded Bioaccessibility of Ru and Q from BBs
2.7. Statistical Analysis
3. Results and Discussion
3.1. The Effect of Fermentation on the Ru and Q Content of Raw and Roasted Buckwheat Flour
3.2. The Effect of Baking on Ru and Q Content in BBs Formulated from Fermented Buckwheat Flour
3.3. The Effect of Digestion on Ru and Q Content in BBs Formulated from Fermented Buckwheat Flour after Digestion In Vitro Procedure
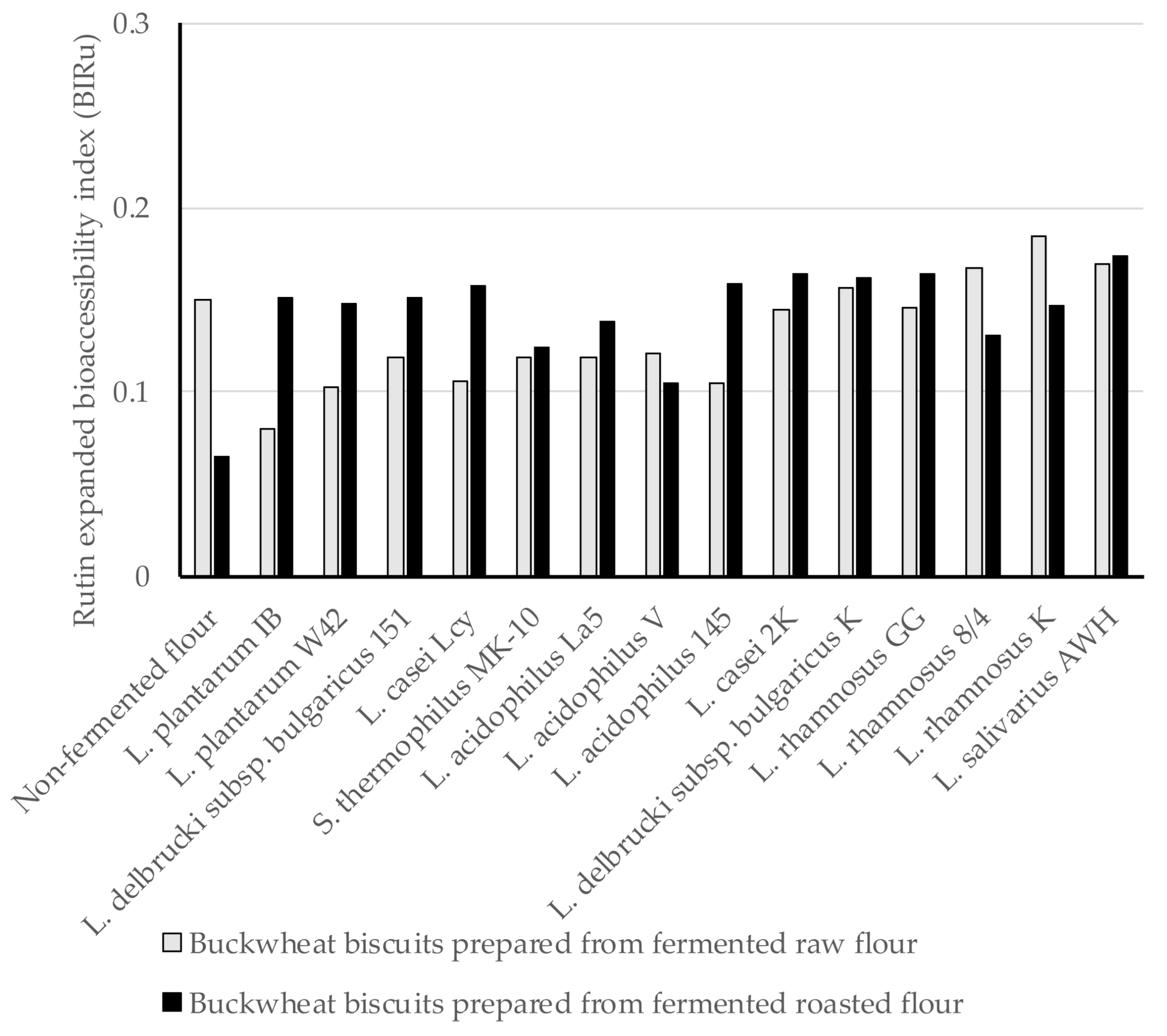
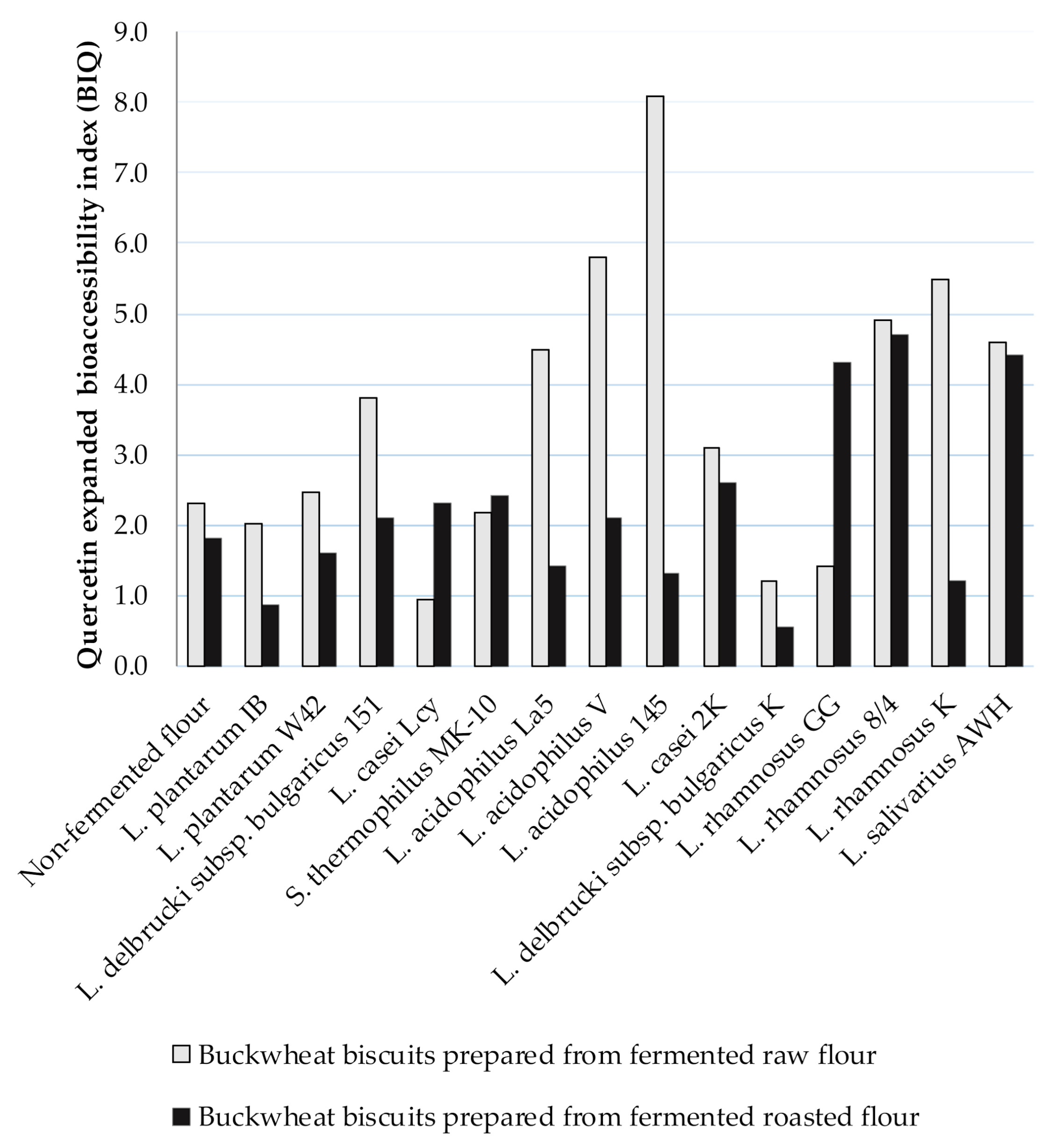
3.4. The Expanded Bioaccessibility of Ru and Q from BBs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.-L.; Zhou, M.-L.; Tang, Y.; Li, F.-L.; Tang, Y.-X.; Shao, J.-R.; Xue, W.-T.; Wu, Y.-M. Bioactive compounds in functional buckwheat food. Food Res. Int. 2012, 49, 389–395. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Zieliński, H. Buckwheat as a functional food and its effects on health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; Laparra, J.; Bączek, N.; Zielinski, H. Buckwheat and buckwheat enriched products exert anti-inflammatory effect on myofibroblasts of colon CCD-18Co. Food Funct. 2018, 9, 3387–3397. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Zieliński, H. Low molecular weight antioxidants and other biologically active components of buckwheat seeds. Eur. J. Plant Sci. Biotechnol. 2009, 3, 29–38. [Google Scholar]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2009, 101, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G.; Kay, C.D.; Crozier, A. The Bioavailability, Transport, and Bioactivity of Dietary Flavonoids: A Review from a Historical Perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Burczynski, F.; Campbell, C.; Pierce, G.; Austria, J.A.; Briggs, C.J. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F-tataricum, and F-homotropicum and their protective effects against lipid peroxidation. Food Res. Int. 2007, 40, 356–364. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Courraud, J.; Berger, J.; Cristol, J.P.; Avallone, S. Stability and bioaccessibility of different forms of carotenoids and vitamin A during in vitro digestion. Food Chem. 2013, 136, 871–877. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Conde-Aguilera, J.A.; Haro, A.; De La Cueva, S.P.; Rufián –Henares, J.A. A combined procedure to evaluate the global antioxidant response of bread. J. Cereal Sci. 2010, 56, 239–246. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Durak, A.; Jamioł, M.; Świeca, M.I. Interactions between antiradical and anti-inflammatory compounds from coffee and coconut affected by gastrointestinal digestion—In in vitro study. LWT-Food Sci. Technol. 2016, 69, 506–514. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Zieliński, H.; Piskuła, M.K.; Zielińska, D.; Szawara-Nowak, D. Buckwheat bioactive compounds, their derived phenolic metabolites and their health benefits. Mol. Nutr. Food Res. 2017, 61, 7–1600475. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. In vitro bioaccessibility of health-related compounds as affected by the formulation of fruit juice- and milk-based beverages. Food Res. Int. 2014, 62, 771–778. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Piskuła, M.K.; Zieliński, H. Recent advances in processing and development of buckwheat derived bakery and non-bakery products—A review. Pol. J. Food Nutr. Sci. 2015, 65, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, H.; Szawara-Nowak, D.; Bączek, N.; Wronkowska, M. Effect of liquid-state fermentation on the antioxidant and functional properties of raw and roasted buckwheat flours. Food Chem. 2019, 271, 291–297. [Google Scholar] [CrossRef]
- Simwaka, J.E.; Chamba, M.V.M.; Huiming, Z.; Masamba, K.G.; Luo, Y. Effect of fermentation on physicochemical and antinutritional factors of complementary foods from millet, sorghum, pumpkin and amaranth seed flours. Int. Food Res. J. 2017, 24, 1869–1879. [Google Scholar]
- Wronkowska, M.; Jeliński, T.; Majkowska, A.; Zieliński, H. Physical properties of buckwheat water biscuits formulated on fermented flours by selected lactic acid bacteria. Pol. J. Food Nutr. Sci. 2018, 68, 25–31. [Google Scholar] [CrossRef]
- American Association of Cereal Chemists. AACC Official Methods 10–52, Baking Quality of Cookie Flour—Micro Method. In Approved Methods of the American Association of Cereal Chemists, 9th ed.; AACC: Minneapolis, MN, USA, 1995. [Google Scholar]
- Hidalgo, A.; Brandolini, A. Heat damage of water biscuits from einkorn, durum and bread wheat flours. Food Chem. 2011, 128, 471–478. [Google Scholar] [CrossRef]
- Zieliński, H.; Szawara-Nowak, D.; Wronkowska, M. Bioaccessibility of anti-AGEs activity, antioxidant capacity and phenolics from water biscuits prepared from fermented buckwheat flours. LWT-Food Sci. Technol. 2020, 123, 109051. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J.; Olejarz, K.; Zieliński, H.; Piskuła, M.K. Metabolites of dietary quercetin: Profile, isolation, identification, and antioxidant activity. J. Funct. Foods. 2014, 11, 121–129. [Google Scholar] [CrossRef]
- Zielińska, D.; Szawara-Nowak, D.; Zieliński, H. Comparison of spectrophotometric and electrochemical methods for the evaluation of antioxidant capacity of buckwheat products after hydrothermal treatment. J. Agric. Food Chem. 2007, 55, 6124–6131. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, H.; Michalska, A.; Amigo-Benavent, M.; del Castillo, M.D.; Piskula, M.K. Changes in protein quality and antioxidant properties of buckwheat seeds and groats induced by roasting. J. Agric. Food Chem. 2009, 57, 4771–4776. [Google Scholar] [CrossRef] [PubMed]
- Wronkowska, M.; Honke, J.; Piskuła, M.K. Effect of solid-state fermentation with Rhizopus oligosporus on bioactive compounds and antioxidant capacity of raw and roasted buckwheat groats. Ital. J. Food Sci. 2015, 27, 424–431. [Google Scholar]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged tempe-type fermentation in order to improve bioactive potential and nutritional parameters of quinoa seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Ganzle, M.G. Phenolic acids and flavonoids in nonfermented and fermented red sorghum (Sorghum bicolor (L.) Moench). J. Agric. Food Chem. 2010, 58, 9214–9220. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Berger, M.; Nepveu, F.; Paul, F.; Daydé, J. Effects of fermentation on the phytochemical composition and antioxidant properties of soy germ. Food Chem. 2008, 109, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Klewicka, E.; Grzelczyk, J.; Gałązka-Czarnecka, I.; Mostowski, R. Lactic acid fermentation of legume seed sprouts as a method of increasing the content of isoflavones and reducing microbial contamination. Food Chem. 2019, 285, 478–484. [Google Scholar] [CrossRef]
- Zieliński, H.; Ciesarová, Z.; Kukurová, K.; Zielinska, D.; Szawara-Nowak, D.; Starowicz, M.; Wronkowska, M. Effect of fermented and unfermented buckwheat flour on functional properties of gluten-free muffins. J. Food Sci. Technol. 2017, 54, 1425–1432. [Google Scholar] [CrossRef]
- Bremus, C.; Herrmann, U.; Bringer-Meyer, S.; Sahm, H. The use of microorganisms in L-ascorbic acid production. J. Biotechnol. 2006, 124, 196–205. [Google Scholar] [CrossRef]
- Dietrych-Szostak, D.; Oleszek, W. Effect of processing on the flavonoids content in buckwheat (Fagopyrum esculentum Moench) grain. J. Agric. Food Chem. 1999, 47, 4384–4387. [Google Scholar] [CrossRef]
- Kreft, I.; Fabjan, N.; Yasumoto, K. Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 2006, 98, 508–512. [Google Scholar] [CrossRef]
- Vogrinčič, M.; Timoracka, M.; Mielichacova, S.; Vollmannova, A.; Kreft, I. Degradation of Rutin and Polyphenols during the Preparation of Tartary Buckwheat Bread. J. Agric. Food Chem. 2010, 58, 4883–4887. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.; Buijsman, M.N.; van Amelsvoort, J.M.; Katan, M.B. Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Dima, S.; Jafari, S.M. Bioavailability of nutraceuticals: Role of the food matrix, processing conditions, the gastrointestinal tract, and nanodelivery systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 954–994. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Loewen, S.; Tsao, R. Bioaccessibility, in vitro antioxidant activities and in vivo anti-inflammatory activities of a purple tomato (Solanum lycopersicum L.). Food Chem. 2014, 159, 353–360. [Google Scholar] [CrossRef]
- Fernàndez-Garcìa, E.; Carvajal-Lérida, I.; Pérez-Gàlvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Wang, S.; Amigo-Benavent, M.; Mateos, R.; Bravo, L.; Sarriá, B. Effects of in vitro digestion and storage on the phenolic content and antioxidant capacity of a red grape pomace. Int. J. Food Sci. Nutr. 2017, 68, 188–200. [Google Scholar] [CrossRef]
- Dufour, C.; Loonis, M.; Delosière, M.; Buffière, C.; Hafnaoui, N.; Santé-Lhoutellier, V.; Rémond, D. The matrix of fruit & vegetables modulates the gastrointestinal bioaccessibility of polyphenols and their impact on dietary protein digestibility. Food Chem. 2018, 240 (Suppl. C), 314–322. [Google Scholar]
- Zhang, Y.; Chen, S.; Wei, C.; Gong, H.; Li, L.; Ye, H. Chemical and cellular assays combined with in vitro digestion to determine the antioxidant activity of flavonoids from Chinese bayberry (Myrica rubra Sieb. et Zucc) leaves. PLoS ONE 2016, 11, e0167484. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Strain/Sample | Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits | |
|---|---|---|---|---|
| Soluble Fraction | Insoluble Fraction | |||
| Control—no fermented flour | 376.4 ± 6.3 a | 90.53 ± 3.45 b | 1.90 ± 0.07 d | 12.09 ± 1.37 c |
| Flour fermented by: | ||||
| L. plantarum IB | 336.4 ± 4.9 *a | 150.26 ± 4.47 *b | 2.49 ± 0.05 *d | 9.47 ± 1.64 *c |
| L. plantarum W42 | 474.3 ± 10.0 *a | 145.66 ± 2.23 *b | 2.43 ± 0.06 *d | 13.51 ± 1.29 c |
| L. delbrucki subsp. bulgaricus 151 | 363.2 ± 17.9 a | 106.03 ± 5.72 *b | 2.31 ± 0.07 *c | 10.37 ± 1.74 c |
| L. casei Lcy | 405.2 ± 0.8 *a | 163.12 ± 3.20 *b | 4.60 ± 0.09 *d | 12.73 ± 0.61 c |
| Streptococcus thermophilus MK-10 | 333.2 ± 16.7 a | 104.92 ± 2.93 *b | 2.36 ± 0.11 *c | 10.17 ± 0.47 c |
| L. acidophilus La5 | 400.0 ± 1.9 *a | 182.79 ± 7.97 *b | 5.63 ± 0.30 *d | 16.26 ± 1.39 *c |
| L. acidophilus V | 245.5 ± 4.4 *a | 122.10 ± 1.38 *b | 3.88 ± 0.02 *d | 10.87 ± 0.34 c |
| L. acidophilus 145 | 348.7 ± 11.7 a | 149.68 ± 3.77 *b | 4.32 ± 0.05 *c | 11.45 ± 0.63 c |
| L. casei 2K | 309.2 ± 2.5 *a | 95.92 ± 2.71 b | 3.67 ± 0.05 *d | 10.20 ± 0.72 c |
| L. delbrucki subsp. bulgaricus K | 309.6 ± 5.7 *a | 102.60 ± 4.25 *b | 4.66 ± 0.05 *c | 11.35 ± 0.20 c |
| L. rhamnosus GG | 366.7 ± 14.2 a | 113.13 ± 5.24 *b | 6.42 ± 0.24 *c | 10.08 ± 0.26 c |
| L. rhamnosus 8/4 | 370.4 ± 0.7 a | 106.93 ± 2.52 *b | 5.40 ± 0.19 *d | 12.45 ± 0.71 c |
| L. rhamnosus K | 319.7 ± 23.1 a | 105.59 ± 3.77 *b | 5.44 ± 0.12 *c | 14.10 ± 1.61 c |
| L. salivarius AWH | 397.0 ± 0.7 *a | 122.13 ± 2.56 *b | 5.07 ± 0.19 *d | 15.52 ± 0.49 *c |
| Strain/Sample | Roasted Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits | |
|---|---|---|---|---|
| Soluble Fraction | Insoluble Fraction | |||
| Control—no fermented flour | 220.4 ± 0.3 a | 61.00 ± 3.04 b | 2.51 ± 0.10 c | 7.57 ± 0.16 c |
| Flour fermented by: | ||||
| L. plantarum IB | 127.7 ± 1.4 *a | 77.27 ± 2.84 *b | 3.09 ± 0.13 *d | 8.56 ± 0.81 c |
| L. plantarum W42 | 148.2 ± 5.4 *a | 70.18 ± 2.83 *b | 2.74 ± 0.09 *d | 7.66 ± 0.35 c |
| L. delbrucki subsp. bulgaricus 151 | 147.2 ± 3.5 *a | 61.11 ± 2.99 b | 1.95 ± 0.02 *d | 7.27 ± 1.09 c |
| L. casei Lcy | 168.6 ± 4.5 a | 60.00 ± 3.10 b | 1.69 ± 0.10 *d | 7.71 ± 0.38 c |
| Streptococcus thermophilus MK-10 | 118.7 ± 0.7 *a | 70.89 ± 1.11 *b | 1.95 ± 0.15 *d | 6.85 ± 0.15 *c |
| L. acidophilus La5 | 199.9 ± 5.8 a | 67.26 ± 2.05 *b | 1.91 ± 0.06 *d | 7.39 ± 0.79 c |
| L. acidophilus V | 134.7 ± 0.8 *a | 82.45 ± 3.95 *b | 2.19 ± 0.09 *d | 6.47 ± 0.64 *c |
| L. acidophilus 145 | 158.1 ± 2.9 *a | 63.50 ± 2.31 b | 2.54 ± 0.07 d | 7.58 ± 0.30 c |
| L. casei 2K | 123.8 ± 9.0 *a | 51.91 ± 1.73 *b | 2.04 ± 0.04 *d | 6.47 ± 0.11 *c |
| L. delbrucki subsp. bulgaricus K | 105.2 ± 0.9 *a | 59.14 ± 1.64 b | 2.74 ± 0.05 *d | 6.82 ± 0.60 c |
| L. rhamnosus GG | 129.3 ± 3.7 *a | 52.78 ± 1.92 *b | 1.12 ± 0.03 *d | 7.53 ± 1.16 c |
| L. rhamnosus 8/4 | 144.2 ± 1.0 *a | 69.45 ± 3.52 *b | 1.62 ± 0.06 *d | 7.51 ± 0.50 c |
| L. rhamnosus K | 107.6 ± 1.7 *a | 62.07 ± 0.78 b | 1.88 ± 0.03 *d | 7.29 ± 0.10 c |
| L. salivarius AWH | 145.3 ± 2.1 *a | 61.64 ± 2.34 b | 2.17 ± 0.02 *d | 8.54 ± 0.56 *c |
| Strain/Sample | Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits | |
|---|---|---|---|---|
| Soluble Fraction | Insoluble Fraction | |||
| Control–no fermented flour | 8.32 ± 0.02 a | 4.55 ± 0.25 b | 7.55 ± 0.28 a | 2.95 ± 0.89 c |
| Flour fermented by: | ||||
| L. plantarum IB | 7.80 ± 0.04 *c | 10.82 ± 0.31 *b | 19.11 ± 0.41 *a | 2.78 ± 0.78 d |
| L. plantarum W42 | 9.17 ± 0.10 *b | 9.64 ± 0.37 *b | 21.87 ± 0.56 *a | 1.96 ± 0.70 c |
| L. delbrucki subsp. bulgaricus 151 | 7.02 ± 0.56 b | 5.44 ± 0.97 b | 17.35 ± 0.48 *a | 3.34 ± 0.92 c |
| L. casei Lcy | 10.85 ± 0.28 *a | 8.53 ± 0.28 *b | 6.20 ± 0.05 *c | 1.86 ± 0.34 d |
| Streptococcus thermophilus MK-10 | 11.19 ± 0.05 *a | 3.81 ± 0.17 *c | 5.96 ± 0.09 *b | 2.28 ± 0.40 d |
| L. acidophilus La5 | 8.52 ± 0.15 a | 2.66 ± 0.16 *b | 9.03 ± 0.25 *a | 2.93 ± 0.64 b |
| L. acidophilus V | 8.38 ± 0.25 a | 2.07 ± 0.08 *c | 8.70 ± 0.010 *a | 3.32 ± 0.51 b |
| L. acidophilus 145 | 4.94 ± 0.03 *b | 1.86 ± 0.17 *d | 12.14 ± 0.28 *a | 2.87 ± 0.49 c |
| L. casei 2K | 11.60 ± 0.23 *b | 5.72 ± 0.28 *c | 15.98 ± 0.22 *a | 1.75 ± 0.20 d |
| L. delbrucki subsp. bulgaricus K | 6.36 ± 0.14 *a | 5.09 ± 0.20 *b | 5.02 ± 0.10 *b | 1.03 ± 0.09 *c |
| L. rhamnosus GG | 7.10 ± 0.37 *a | 5.39 ± 0.43 *c | 6.16 ± 0.23 *b | 1.56 ± 0.44 d |
| L. rhamnosus 8/4 | 7.05 ± 0.10 *b | 2.69 ± 0.05 *c | 11.35 ± 0.05 *a | 1.97 ± 0.11 d |
| L. rhamnosus K | 5.87 ± 0.04 *b | 2.06 ± 0.18 *c | 9.77 ± 0.35 *a | 1.60 ± 0.75 c |
| L. salivarius AWH | 7.05 ± 0.13 *b | 3.43 ± 0.26 *d | 9.87 ± 0.06 *a | 5.91 ± 0.80 *c |
| Strain/Sample | Roasted Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits | |
|---|---|---|---|---|
| Soluble Fraction | Insoluble Fraction | |||
| Control—no fermented flour | 3.82 ± 0.04 b | 1.37 ± 0.01 c | 7.72 ± 0.45 a | 0.03 ± 0.01 d |
| Flour fermented by: | ||||
| L. plantarum IB | 4.22 ± 0.12 *a | 2.99 ± 0.16 *b | 2.37 ± 0.08 c | 0.18 ± 0.02 *d |
| L. plantarum W42 | 6.48 ± 0.05 *a | 3.00 ± 0.08 *c | 4.83 ± 0.13 b | 0.10 ± 0.05 d |
| L. delbrucki subsp. bulgaricus 151 | 3.55 ± 0.11 b | 2.49 ± 0.13 *c | 5.06 ± 0.12 a | 0.16 ± 0.18 d |
| L. casei Lcy | 6.33 ± 0.44 *a | 2.95 ± 0.15 *b | 6.55 ± 0.08 a | 0.11 ± 0.01 *c |
| Streptococcus thermophilus MK-10 | 3.50 ± 0.05 *b | 2.74 ± 0.03 *c | 6.22 ± 0.19 a | 0.22 ± 0.02 *d |
| L. acidophilus La5 | 3.70 ± 0.14 b | 3.85 ± 0.30 *b | 5.33 ± 0.32 a | 0.11 ± 0.04 *c |
| L. acidophilus V | 3.66 ± 0.17 b | 2.69 ± 0.01 *c | 5.67 ± 0.07 a | 0.07 ± 0.00 *d |
| L. acidophilus 145 | 3.25 ± 0.10 *c | 3.68 ± 0.10 *b | 4.72 ± 0.07 a | 0.07 ± 0.04 d |
| L. casei 2K | 3.28 ± 0.09 *b | 1.92 ± 0.11 *c | 4.97 ± 0.13 a | 0.05 ± 0.01 d |
| L. delbrucki subsp. bulgaricus K | 3.33 ± 0.01 *c | 7.96 ± 0.27 *a | 4.23 ± 0.13 b | 0.09 ± 0.01 *d |
| L. rhamnosus GG | 1.58 ± 0.07 *b | 1.26 ± 0.02 *c | 5.32 ± 0.17 a | 0.14 ± 0.16 d |
| L. rhamnosus 8/4 | 2.94 ± 0.01 *b | 2.07 ± 0.03 *c | 9.74 ± 0.20 a | 0.11 ± 0.01 *d |
| L. rhamnosus K | 3.93 ± 0.06 c | 6.47 ± 0.51 *b | 7.78 ± 0.45 a | 0.09 ± 0.04 d |
| L. salivarius AWH | 4.28 ± 0.08 *b | 1.83 ± 0.01 *c | 7.94 ± 0.16 a | 0.08 ± 0.05 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieliński, H.; Wiczkowski, W.; Honke, J.; Piskuła, M.K. In Vitro Expanded Bioaccessibility of Quercetin-3-Rutinoside and Quercetin Aglycone from Buckwheat Biscuits Formulated from Flours Fermented by Lactic Acid Bacteria. Antioxidants 2021, 10, 571. https://doi.org/10.3390/antiox10040571
Zieliński H, Wiczkowski W, Honke J, Piskuła MK. In Vitro Expanded Bioaccessibility of Quercetin-3-Rutinoside and Quercetin Aglycone from Buckwheat Biscuits Formulated from Flours Fermented by Lactic Acid Bacteria. Antioxidants. 2021; 10(4):571. https://doi.org/10.3390/antiox10040571
Chicago/Turabian StyleZieliński, Henryk, Wiesław Wiczkowski, Joanna Honke, and Mariusz Konrad Piskuła. 2021. "In Vitro Expanded Bioaccessibility of Quercetin-3-Rutinoside and Quercetin Aglycone from Buckwheat Biscuits Formulated from Flours Fermented by Lactic Acid Bacteria" Antioxidants 10, no. 4: 571. https://doi.org/10.3390/antiox10040571