Using Dual-Site Transcranial Magnetic Stimulation to Probe Connectivity between the Dorsolateral Prefrontal Cortex and Ipsilateral Primary Motor Cortex in Humans


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Electromyographic Recordings
2.3. Transcranial Magnetic Stimulation (TMS)
2.4. Localization of TMS Cortical Targets
2.5. Data Acquisiton and Analysis
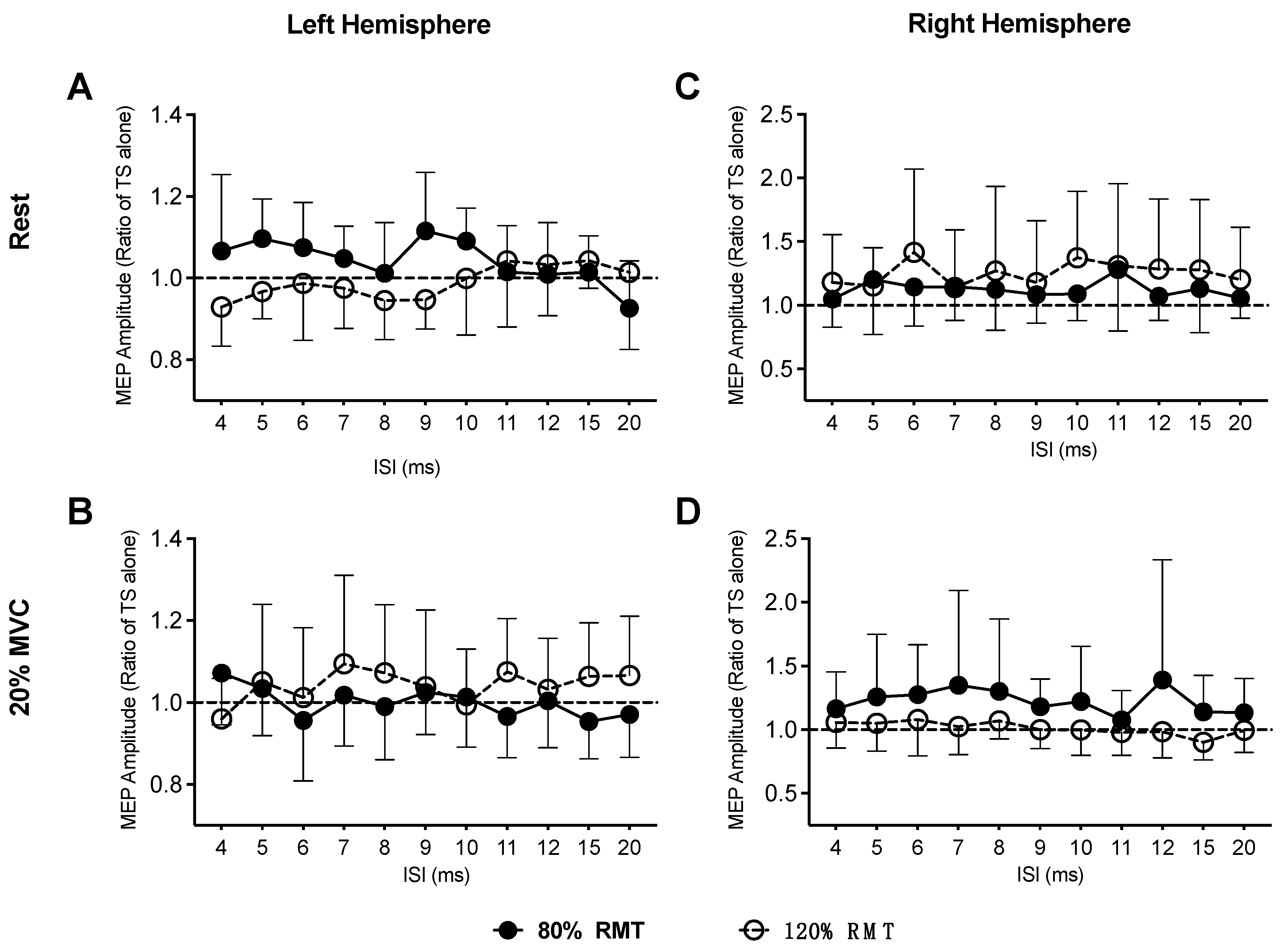
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Petrides, M. Lateral prefrontal cortex: Architectonic and functional organization. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Sallet, J.; Mars, R.B.; Noonan, M.P.; Neubert, F.X.; Jbabdi, S.; O’Reilly, J.X.; Filippini, N.; Thomas, A.G.; Rushworth, M.F. The Organization of Dorsal Frontal Cortex in Humans and Macaques. J. Neurosci. 2013, 33, 12255–12274. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.R.; Muthalib, M.; Perrey, S.; Galka, A.; Granert, O.; Wolff, S.; Heute, U.; Deuschl, G.; Raethjen, J.; Muthuraman, M. Effective Connectivity of Cortical Sensorimotor Networks During Finger Movement Tasks: A Simultaneous fNIRS, fMRI, EEG Study. Brain Topogr. 2016, 29, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Bates, J.F.; Goldman-Rakic, P.S. Prefrontal connections of medial motor areas in the rhesus monkey. J. Comp. Neurol. 1993, 336, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Gabbott, P.L.A.; Warner, T.A.; Jays, P.R.L.; Salway, P.; Busby, S.J. Prefrontal cortex in the rat: Projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol. 2005, 492, 145–177. [Google Scholar] [CrossRef] [PubMed]
- Selemon, L.D.; Goldman-Rakic, P.S. Common cortical and subcortical targets of the dorsolateral prefrontal and posterior parietal cortices in the rhesus monkey: Evidence for a distributed neural network subserving spatially guided behavior. J. Neurosci. 1988, 8, 4049–4068. [Google Scholar] [CrossRef]
- Xiao, D.; Zikopoulos, B.; Barbas, H. Laminar and modular organization of prefrontal projections to multiple thalamic nuclei. Neuroscience 2009, 161, 1067–1081. [Google Scholar] [CrossRef]
- Yeterian, E.H.; Pandya, D.N.; Tomaiuolo, F.; Petrides, M. The cortical connectivity of the prefrontal cortex in the monkey brain. Cortex 2012, 48, 58–81. [Google Scholar] [CrossRef]
- Szameitat, A.J.; Schubert, T.; Müller, K.; Von Cramon, D.Y. Localization of executive functions in dual-task performance with fMRI. J. Cogn. Neurosci. 2002, 14, 1184–1199. [Google Scholar] [CrossRef]
- Rowe, J.B.; Toni, I.; Josephs, O.; Frackowiak, R.S.; Passingham, R.E. The prefrontal cortex: response selection or maintenance within working memory? Science 2000, 288, 1656–1660. [Google Scholar] [CrossRef]
- Jahanshahi, M.; Jenkins, I.H.; Brown, R.G.; Marsden, C.D.; Passingham, R.E.; Brooks, D.J. Self-initiated versus externally triggered movements. I. An investigation using measurement of regional cerebral blood flow with PET and movement-related potentials in normal and Parkinson’s disease subjects. Brain 1995, 118, 913–933. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahi, M.; Obeso, I.; Rothwell, J.C.; Obeso, J.A. A fronto–striato–subthalamic–pallidal network for goal-directed and habitual inhibition. Nat. Publ. Gr. 2015, 16, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Halko, M.A.; Eldaief, M.C.A. Pascual-Leone, Measuring and manipulating brain connectivity with resting state functional connectivity magnetic resonance imaging (fcMRI) and transcranial magnetic stimulation (TMS). Neuroimage 2012, 62, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Fujiyama, H.; Van Soom, J.; Rens, G.; Gooijers, J.; Leunissen, I.; Levin, O.; Swinnen, S.P. Age-Related Changes in Frontal Network Structural and Functional Connectivity in Relation to Bimanual Movement Control. J. Neurosci. 2016, 36, 1808–1822. [Google Scholar] [CrossRef] [PubMed]
- King, B.R.; van Ruitenbeek, P.; Leunissen, I.; Cuypers, K.; Heise, K.F.; Santos Monteiro, T.; Hermans, L.; Levin, O.; Albouy, G.; Mantini, D. Age-Related Declines in Motor Performance are Associated with Decreased Segregation of Large-Scale Resting State Brain Networks. Cereb. Cortex 2017, 28, 4390–4402. [Google Scholar] [CrossRef] [PubMed]
- Michely, J.; Volz, L.J.; Hoffstaedter, F.; Tittgemeyer, M.; Eickhoff, S.B.; Fink, G.R.; Grefkes, C. Network connectivity of motor control in the ageing brain. NeuroImage Clin. 2018, 18, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.B.; Lipps, D.B. Motor control and aging: Links to age-related brain structural, functional, and biochemical effects. Neurosci. Biobehav. Rev. 2010, 34, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.C.; Tran, X.; Cramer, S.C. Age-related variability in performance of a motor action selection task is related to differences in brain function and structure among older adults. NeuroImage 2014, 86, 326–334. [Google Scholar] [CrossRef]
- Grefkes, C.; Fink, G.R. Reorganization of cerebral networks after stroke: new insights from neuroimaging with connectivity approaches. Brain 2011, 134, 1264–1276. [Google Scholar] [CrossRef]
- Fox, M.D.; Buckner, R.L.; Liu, H.; Chakravarty, M.M.; Lozano, A.M.; Pascual-Leone, A. Resting-state networks link invasive and noninvasive brain stimulation across diverse psychiatric and neurological diseases. Proc. Natl. Acad. Sci. USA 2014, 111, E4367–E4375. [Google Scholar] [CrossRef]
- Rowe, J.B.; Siebner, H.R. The motor system and its disorders. NeuroImage 2012, 61, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Rothwell, J.C. TMS investigations into the task-dependent functional interplay between human posterior parietal and motor cortex. Behav. Brain Res. 2009, 202, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.C. Using transcranial magnetic stimulation methods to probe connectivity between motor areas of the brain. Hum. Mov. Sci. 2011, 30, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M.; Di Iorio, R.; Rossini, P.M.; Park, J.E.; Chen, R.; Celnik, P.; Strafella, A.P.; Matsumoto, H.; Ugawa, Y. Contribution of transcranial magnetic stimulation to assessment of brain connectivity and networks. Clin. Neurophysiol. 2017, 128, 2125–2139. [Google Scholar] [CrossRef] [PubMed]
- Davare, M.; Kraskov, A.; Rothwell, J.C.; Lemon, R.N. Interactions between areas of the cortical grasping network. Curr. Opin. Neurobiol. 2011, 21, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Bäumer, T.; Schippling, S.; Kroeger, J.; Zittel, S.; Koch, G.; Thomalla, G.; Rothwell, J.C.; Siebner, H.R.; Orth, M.; Münchau, A. Inhibitory and facilitatory connectivity from ventral premotor to primary motor cortex in healthy humans at rest—A bifocal TMS study. Clin. Neurophysiol. 2009, 120, 1724–1731. [Google Scholar] [CrossRef] [PubMed]
- Byblow, W.D.; Coxon, J.P.; Stinear, C.M.; Fleming, M.K.; Williams, G.; Müller, J.F.M.; Ziemann, U. Functional Connectivity Between Secondary and Primary Motor Areas Underlying Hand–Foot Coordination. J. Neurophysiol. 2007, 98, 414–422. [Google Scholar] [CrossRef]
- Civardi, C.; Cantello, R.; Asselman, P.; Rothwell, J.C. Transcranial Magnetic Stimulation Can Be Used to Test Connections to Primary Motor Areas from Frontal and Medial Cortex in Humans. NeuroImage 2001, 14, 1444–1453. [Google Scholar] [CrossRef]
- Davare, M.; Lemon, R.; Olivier, E. Selective modulation of interactions between ventral premotor cortex and primary motor cortex during precision grasping in humans. J. Physiol. 2008, 586, 2735–2742. [Google Scholar] [CrossRef]
- Davare, M.; Montague, K.; Olivier, E.; Rothwell, J.C.; Lemon, R.N. Ventral premotor to primary motor cortical interactions during object-driven grasp in humans. Cortex 2009, 45, 1050–1057. [Google Scholar] [CrossRef]
- Vesia, M.; Culham, J.C.; Jegatheeswaran, G.; Isayama, R.; Le, A.; Davare, M.; Chen, R. Functional interaction between human dorsal premotor cortex and the ipsilateral primary motor cortex for grasp plans. NeuroReport 2018, 29, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Shirota, Y.; Hamada, M.; Terao, Y.; Ohminami, S.; Tsutsumi, R.; Ugawa, Y.; Hanajima, R. Increased primary motor cortical excitability by a single-pulse transcranial magnetic stimulation over the supplementary motor area. Exp. Brain Res. 2012, 219, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, A.; Jin, S.H.; Joutsen, A.; Poston, B.; Aizen, J.; Ellenstein, A.; Hallett, M. Timing-dependent modulation of the posterior parietal cortex–primary motor cortex pathway by sensorimotor training. J. Neurophysiol. 2012, 107, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, A.N.; Chao, C.C.; Paine, R.; Hallett, M. Mapping Different Intra-Hemispheric Parietal-Motor Networks Using Twin Coil TMS. Brain Stimul. 2013, 6, 384–389. [Google Scholar] [CrossRef]
- Karabanov, A.N.; Ritterband-Rosenbaum, A.; Christensen, M.S.; Siebner, H.R.; Nielsen, J.B. Modulation of fronto-parietal connections during the rubber hand illusion. Eur. J. Neurosci. 2017, 45, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Fernandez Del Olmo, M.; Cheeran, B.; Ruge, D.; Schippling, S.; Caltagirone, C.; Rothwell, J.C. Focal Stimulation of the Posterior Parietal Cortex Increases the Excitability of the Ipsilateral Motor Cortex. J. Neurosci. 2007, 27, 6815–6822. [Google Scholar] [CrossRef]
- Koch, G.; Del Olmo, M.F.; Cheeran, B.; Schippling, S.; Caltagirone, C.; Driver, J.; Rothwell, J.C. Functional Interplay between Posterior Parietal and Ipsilateral Motor Cortex Revealed by Twin-Coil Transcranial Magnetic Stimulation during Reach Planning toward Contralateral Space. J. Neurosci. 2008, 28, 5944–5953. [Google Scholar] [CrossRef] [PubMed]
- Schintu, S.; Martín-Arévalo, E.; Vesia, M.; Rossetti, Y.; Salemme, R.; Pisella, L.; Farnè, A.; Reilly, K.T. Paired-Pulse Parietal-Motor Stimulation Differentially Modulates Corticospinal Excitability across Hemispheres When Combined with Prism Adaptation. Neural Plast. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Vesia, M.; Bolton, D.A.; Mochizuki, G.; Staines, W.R. Human parietal and primary motor cortical interactions are selectively modulated during the transport and grip formation of goal-directed hand actions. Neuropsychologia 2013, 51, 410–417. [Google Scholar] [CrossRef]
- Vesia, M.; Barnett-Cowan, M.; Elahi, B.; Jegatheeswaran, G.; Isayama, R.; Neva, J.L.; Davare, M.; Staines, W.R.; Culham, J.C.; Chen, R. Human dorsomedial parieto-motor circuit specifies grasp during the planning of goal-directed hand actions. Cortex 2017, 92, 175–186. [Google Scholar] [CrossRef]
- Brown, M.J.N.; Weissbach, A.; Pauly, M.G.; Vesia, M.; Gunraj, C.; Baarbé, J.; Münchau, A.; Bäumer, T.; Chen, R. Somatosensory-motor cortex interactions measured using dual-site transcranial magnetic stimulation. Brain Stimul. 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ziluk, A.; Premji, A.; Nelson, A.J. Functional connectivity from area 5 to primary motor cortex via paired-pulse transcranial magnetic stimulation. Neurosci. Lett. 2010, 484, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, L. Transcranial magnetic mapping of the short-latency modulations of corticospinal activity from the ipsilateral hemisphere during rest. Front. Neural Circuits 2011, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lafleur, L.P.; Tremblay, S.; Whittingstall, K.; Lepage, J.F. Assessment of Effective Connectivity and Plasticity with Dual-Coil Transcranial Magnetic Stimulation. Brain Stimul. 2016, 9, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Galea, J.M.; Casula, E.P.; Falkai, P.; Bestmann, S.; Rothwell, J.C. Muscle and Timing-specific Functional Connectivity between the Dorsolateral Prefrontal Cortex and the Primary Motor Cortex. J. Cogn. Neurosci. 2013, 25, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Geyer, S.; Matelli, M.; Luppino, G.; Zilles, K. Functional neuroanatomy of the primate isocortical motor system. Anat. Embryol. 2000, 202, 443–474. [Google Scholar] [CrossRef] [PubMed]
- Johansen-Berg, H.; Behrens, T.E.J.; Robson, M.D.; Drobnjak, I.; Rushworth, M.F.S.; Brady, J.M.; Smith, S.M.; Higham, D.J.; Matthews, P.M. Changes in connectivity profiles define functionally distinct regions in human medial frontal cortex. Proc. Natl. Acad. Sci. USA 2004, 101, 13335–13340. [Google Scholar] [CrossRef]
- Picard, N.; Strick, P.L. Imaging the premotor areas. Curr. Opin. Neurobiol. 2001, 11, 663–672. [Google Scholar] [CrossRef]
- Mars, R.B.; Klein, M.C.; Neubert, F.X.; Olivier, E.; Buch, E.R.; Boorman, E.D.; Rushworth, M.F.S. Short-Latency Influence of Medial Frontal Cortex on Primary Motor Cortex during Action Selection under Conflict. J. Neurosci. 2009, 29, 6926–6931. [Google Scholar] [CrossRef]
- Picazio, S.; Veniero, D.; Ponzo, V.; Caltagirone, C.; Gross, J.; Thut, G.; Koch, G. Prefrontal Control over Motor Cortex Cycles at Beta Frequency during Movement Inhibition. Curr. Biol. 2014, 24, 2940–2945. [Google Scholar] [CrossRef]
- Ni, Z.; Gunraj, C.; Nelson, A.J.; Yeh, I.J.; Castillo, G.; Hoque, T.; Chen, R. Two Phases of Interhemispheric Inhibition between Motor Related Cortical Areas and the Primary Motor Cortex in Human. Cereb. Cortex 2009, 19, 1654–1665. [Google Scholar] [CrossRef] [PubMed]
- Varma, V.R.; Hausdorff, J.M.; Studenski, S.A.; Rosano, C.; Camicioli, R.; Alexander, N.B.; Chen, W.G.; Lipsitz, L.A.; Carlson, M.C. Aging, the Central Nervous System, and Mobility in Older Adults: Interventions. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2016, 71, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Manor, B.; Zhou, J.; Harrison, R.; Lo, O.Y.; Travison, T.G.; Hausdorff, J.M.; Pascual-Leone, A.; Lipsitz, L. Transcranial Direct Current Stimulation May Improve Cognitive-Motor Function in Functionally Limited Older Adults. Neurorehabil. Neural Repair 2018, 32, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Steenhuis, R.E.; Bryden, M.P. Different Dimensions of Hand Preference That Relate to Skilled and Unskilled Activities. Cortex 1989, 25, 289–304. [Google Scholar] [CrossRef]
- Keel, J.C.; Smith, M.J.; Wassermann, E.M. A safety screening questionnaire for transcranial magnetic stimulation. Clin. Neurophysiol. 2001, 112, 720. [Google Scholar] [CrossRef]
- Vesia, M.; Davare, M. Decoding Action Intentions in Parietofrontal Circuits. J. Neurosci. 2011, 31, 16491–16493. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Yousry, T.A.; Schmid, U.D.; Alkadhi, H.; Schmidt, D.; Peraud, A.; Buettner, A.; Winkler, P. Localization of the motor hand area to a knob on the precentral gyrus. A new landmark. Brain J. Neurol. 1997, 120, 141–157. [Google Scholar] [CrossRef]
- Rowe, J.; Friston, K.; Frackowiak, R.; Passingham, R. Attention to Action: Specific Modulation of Corticocortical Interactions in Humans. NeuroImage 2002, 17, 988–998. [Google Scholar] [CrossRef]
- Rowe, J.B. The Prefrontal Cortex shows Context-specific Changes in Effective Connectivity to Motor or Visual Cortex during the Selection of Action or Colour. Cereb. Cortex 2004, 15, 85–95. [Google Scholar] [CrossRef]
- Sack, A.T.; Cohen Kadosh, R.; Schuhmann, T.; Moerel, M.; Walsh, V.; Goebel, R. Optimizing functional accuracy of TMS in cognitive studies: A comparison of methods. J. Cogn. Neurosci. 2009, 21, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Middleton, F.A.; Strick, P.L. Basal ganglia and cerebellar loops: motor and cognitive circuits. Brain Res. Rev. 2000, 31, 236–250. [Google Scholar] [CrossRef]
- Middleton, F.A.; Strick, P.L. Basal-ganglia “projections” to the prefrontal cortex of the primate. Cereb. Cortex 2002, 12, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Guillery, R.W. Branching Thalamic Afferents Link Action and Perception. J. Neurophysiol. 2003, 90, 539–548. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guillery, R.W.; Sherman, S.M. The thalamus as a monitor of motor outputs. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2002, 357, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Zikopoulos, B.; Barbas, H. Prefrontal Projections to the Thalamic Reticular Nucleus form a Unique Circuit for Attentional Mechanisms. J. Neurosci. 2006, 26, 7348–7361. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.; Park, J.E.; Srivanitchapoom, P.; Paine, R.; Thirugnanasambandam, N.; Kukke, S.N.; Hallett, M. Probing the interaction of the ipsilateral posterior parietal cortex with the premotor cortex using a novel transcranial magnetic stimulation technique. Clin. Neurophysiol. 2016, 127, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Latorre, A.; Rocchi, L.; Berardelli, A.; Bhatia, K.P.; Rothwell, J.C. The interindividual variability of transcranial magnetic stimulation effects: Implications for diagnostic use in movement disorders. Mov. Disord. 2019, 74, 113–114. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Participant | Left DLPFC | Right DLPFC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anatomical Location | Stimulation Location | Anatomical Location | Stimulation Location | |||||||||
| x | y | z | x | y | z | x | y | z | x | y | z | |
| 1 | −32.61 | 46.64 | 20.29 | −31.89 | 49.39 | 15.74 | 35.64 | 40.82 | 19.71 | 30.14 | 44.27 | 25.78 |
| 2 | −39.74 | 30.10 | 22.41 | −27.39 | 39.06 | 29.32 | 38.59 | 33.28 | 22.85 | 38.46 | 33.86 | 22.04 |
| 3 | −28.89 | 26.58 | 21.29 | −17.91 | 31.44 | 28.36 | 26.52 | 35.2 | 18.03 | 19.28 | 30.38 | 28.54 |
| 4 | −31.33 | 45.31 | 18.36 | −12.87 | 54.64 | 36.09 | 29.68 | 45.89 | 23.71 | 28.51 | 39.2 | 32.86 |
| 5 | −34.37 | 45.32 | 11.64 | −32.48 | 39.82 | 32.12 | 33.4 | 51.22 | 15.74 | 31.81 | 46.05 | 27.2 |
| 6 | −29.4 | 38.34 | 17.5 | −27.67 | 41.07 | 24.79 | 33.65 | 46.6 | 15.22 | 37.82 | 34.51 | 16 |
| 7 | −26.02 | 45.86 | 18.75 | −24.3 | 52.63 | 5.25 | 24.86 | 50.07 | 14.13 | 15.04 | 56.61 | 16.78 |
| 8 | −32.13 | 43.11 | 16.13 | −38.18 | 38.16 | 7.57 | 33.02 | 44.88 | 13.76 | 29.75 | 44.15 | 14.5 |
| 9 | −25.33 | 51.12 | 21.38 | −25.15 | 50.72 | 13.62 | 22.69 | 50.8 | 19.22 | 25.85 | 53.87 | 17.16 |
| 10 | −36.71 | 47.95 | 12.28 | −25.27 | 52.59 | 31.31 | 32.48 | 55.01 | 16.6 | 29.09 | 55.67 | 23.03 |
| 11 | −34.97 | 44.93 | 12.01 | −36.03 | 44.00 | 13.72 | 25.87 | 55.16 | 18.6 | 24.73 | 55.68 | 25.36 |
| 12 | −39.76 | 32.76 | 1.26 | −23.92 | 50.13 | 14.39 | 33.76 | 47.5 | 5.82 | 29.81 | 46.04 | 4.44 |
| Group Mean ± SD | −32.61 ± 4.77 | 41.50 ± 7.76 | 16.11 ± 6.01 | −26.92 ± 7.18 | 45.30 ± 7.36 | 21.02 ± 10.46 | 30.85 ± 4.87 | 46.37 ± 7.00 | 16.95 ± 4.71 | 28.36 ± 6.68 | 45.02 ± 9.18 | 21.14 ± 7.73 |
| Participant | Left M1 | Right M1 | ||||
|---|---|---|---|---|---|---|
| RMT | Rest 1 mV | Active 1 mV | RMT | Rest 1 mV | Active 1 mV | |
| 1 | 47 | 58 | 47 | 49 | 49 | 40 |
| 2 | 45 | 56 | 45 | 44 | 50 | 46 |
| 3 | 58 | 75 | 62 | 57 | 58 | 45 |
| 4 | 53 | 60 | 52 | 54 | 58 | 47 |
| 5 | 50 | 68 | 45 | 37 | 39 | 34 |
| 6 | 51 | 56 | 45 | 51 | 54 | 41 |
| 7 | 47 | 65 | 56 | 51 | 63 | 52 |
| 8 | 52 | 70 | 54 | 53 | 72 | 56 |
| 9 | 45 | 49 | 42 | 39 | 39 | 35 |
| 10 | 59 | 70 | 62 | 62 | 73 | 59 |
| 11 | 41 | 45 | 39 | 52 | 52 | 42 |
| 12 | 50 | 51 | 45 | 50 | 55 | 48 |
| Group Mean ± SD | 49.8 ± 5.3 | 60.3 ± 9.4 | 49.5 ± 7.6 | 49.9 ± 7.1 | 55.2 ± 10.8 | 45.4 ± 7.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, M.J.N.; Goldenkoff, E.R.; Chen, R.; Gunraj, C.; Vesia, M. Using Dual-Site Transcranial Magnetic Stimulation to Probe Connectivity between the Dorsolateral Prefrontal Cortex and Ipsilateral Primary Motor Cortex in Humans. Brain Sci. 2019, 9, 177. https://doi.org/10.3390/brainsci9080177
Brown MJN, Goldenkoff ER, Chen R, Gunraj C, Vesia M. Using Dual-Site Transcranial Magnetic Stimulation to Probe Connectivity between the Dorsolateral Prefrontal Cortex and Ipsilateral Primary Motor Cortex in Humans. Brain Sciences. 2019; 9(8):177. https://doi.org/10.3390/brainsci9080177
Chicago/Turabian StyleBrown, Matt J.N., Elana R. Goldenkoff, Robert Chen, Carolyn Gunraj, and Michael Vesia. 2019. "Using Dual-Site Transcranial Magnetic Stimulation to Probe Connectivity between the Dorsolateral Prefrontal Cortex and Ipsilateral Primary Motor Cortex in Humans" Brain Sciences 9, no. 8: 177. https://doi.org/10.3390/brainsci9080177
APA StyleBrown, M. J. N., Goldenkoff, E. R., Chen, R., Gunraj, C., & Vesia, M. (2019). Using Dual-Site Transcranial Magnetic Stimulation to Probe Connectivity between the Dorsolateral Prefrontal Cortex and Ipsilateral Primary Motor Cortex in Humans. Brain Sciences, 9(8), 177. https://doi.org/10.3390/brainsci9080177