Distinct Montages of Slow Oscillatory Transcranial Direct Current Stimulation (so-tDCS) Constitute Different Mechanisms during Quiet Wakefulness


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
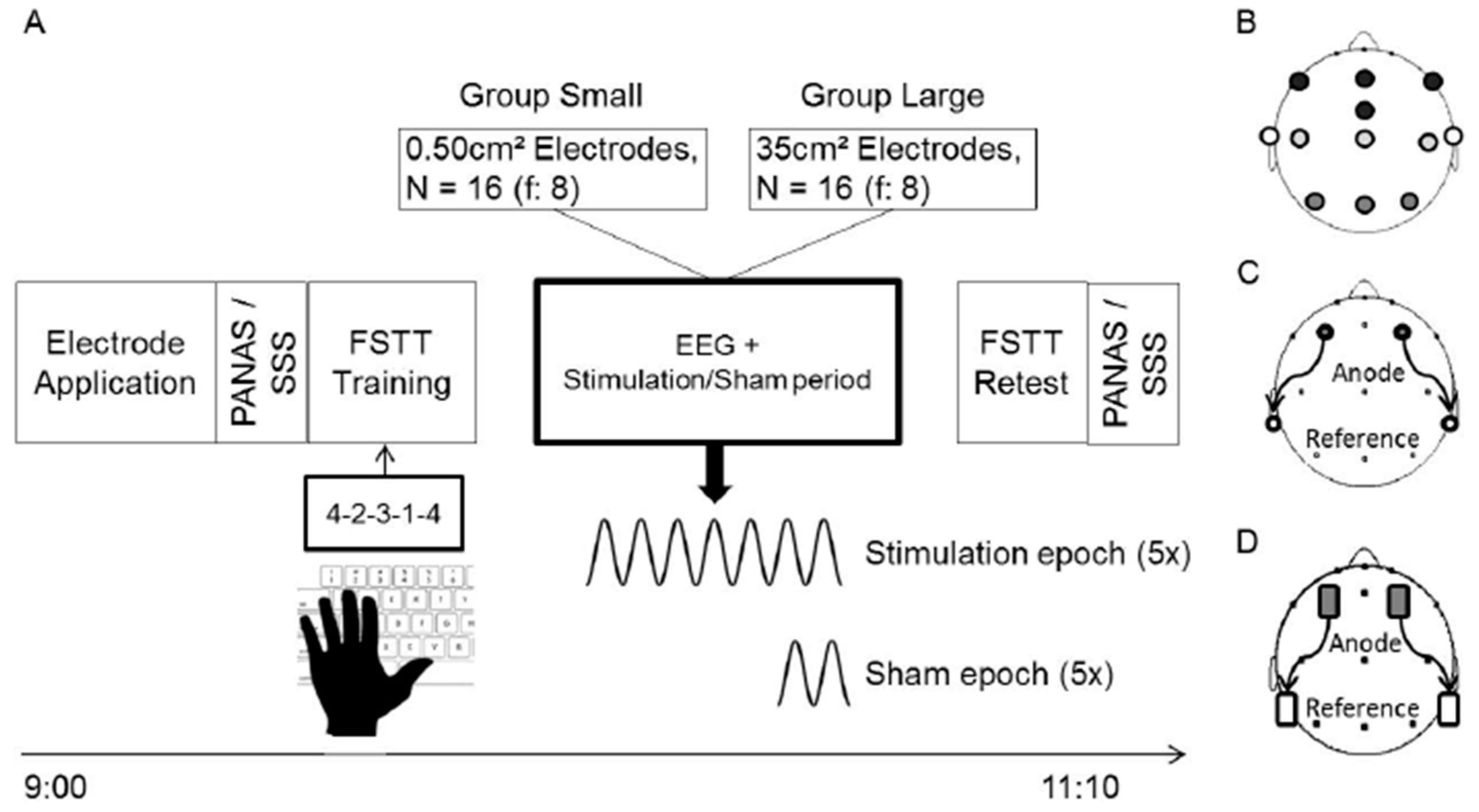
2. Materials and Methods
2.1. Procedure
2.2. Participants
2.3. EEG Acquisition/Analysis
2.4. So-tDCS
2.5. Finger Sequence Tapping Task and Psychometric Questionnaires
2.6. Statistical Analysis
2.7. Exploratory Theta Synchronization and Alpha Desynchronization
3. Results
3.1. Blinding of The Stimulation
3.2. Effect of Electrode Size and Current Strength on So-tDCS Induced Eeg Activity
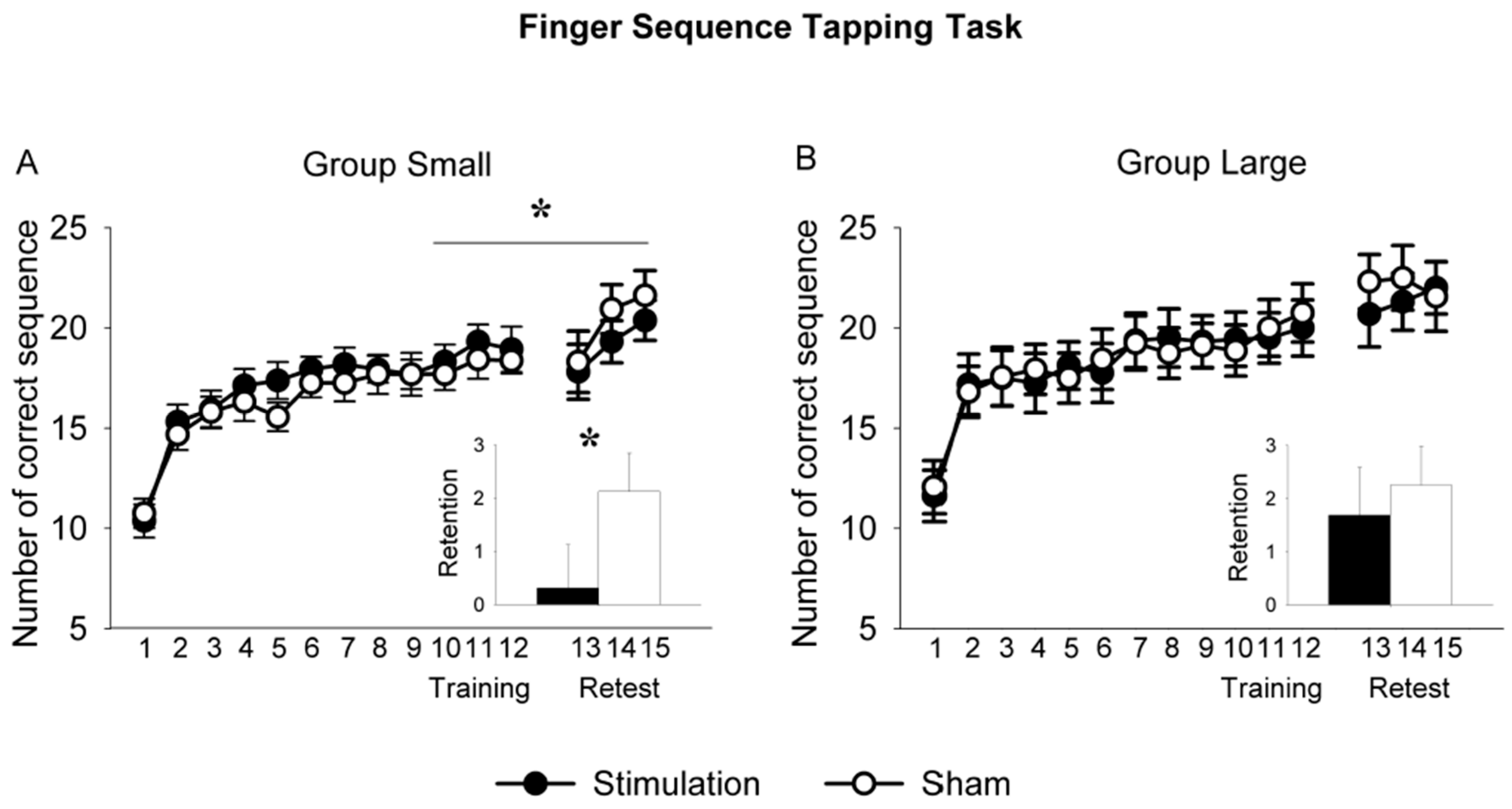
3.3. So-tDCS Effect on Behavioral Parameters
3.4. Contrasting Effects of Stimulation Parameters
3.5. Increased Theta Synchronization in Conjunction with Alpha Desynchronization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Ferrucci, R.; Bortolomasi, M.; Vergari, M.; Tadini, L.; Boggio, P.; Giacopuzzi, M.; Barbieri, S.; Priori, A. Transcranial direct current stimulation (tDCS) in unipolar vs. bipolar depressive disorder. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 2011, 35, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Del Felice, A.; Magalini, A.; Masiero, S. Slow-oscillatory transcranial direct current stimulation modulates memory in temporal lobe epilepsy by altering sleep spindle generators: A possible rehabilitation tool. Brain Stimul. 2015, 8, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Chalah, M.A.; Ayache, S.S. Noninvasive Brain Stimulation and Psychotherapy in Anxiety and Depressive Disorders: A Viewpoint. Brain Sci. 2019, 9, 82. [Google Scholar] [CrossRef]
- Antal, A.; Boros, K.; Poreisz, C.; Chaieb, L.; Terney, D.; Paulus, W. Comparatively weak after-effects of transcranial alternating current stimulation (tACS) on cortical excitability in humans. Brain Stimul. 2008, 1, 97–105. [Google Scholar] [CrossRef]
- Groppa, S.; Bergmann, T.; Siems, C.; Molle, M.; Marshall, L.; Siebner, H. Slow-oscillatory transcranial direct current stimulation can induce bidirectional shifts in motor cortical excitability in awake humans. Neuroscience 2010, 166, 1219–1225. [Google Scholar] [CrossRef]
- Ho, K.-A.; Taylor, J.L.; Chew, T.; Gálvez, V.; Alonzo, A.; Bai, S.; Dokos, S.; Loo, C.K.; Information, P.E.K.F.C. The Effect of Transcranial Direct Current Stimulation (tDCS) Electrode Size and Current Intensity on Motor Cortical Excitability: Evidence from Single and Repeated Sessions. Brain Stimul. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Learmonth, G.; Felisatti, F.; Siriwardena, N.; Checketts, M.; Benwell, C.S.Y.; Märker, G.; Thut, G.; Harvey, M. No Interaction between tDCS Current Strength and Baseline Performance: A Conceptual Replication. Front. Mol. Neurosci. 2017, 11, 664. [Google Scholar] [CrossRef]
- Penolazzi, B.; Pastore, M.; Mondini, S. Electrode montage dependent effects of transcranial direct current stimulation on semantic fluency. Behav. Brain Res. 2013, 248, 129–135. [Google Scholar] [CrossRef]
- Veniero, D.; Benwell, C.S.Y.; Ahrens, M.M.; Thut, G. Inconsistent Effects of Parietal alpha-tACS on Pseudoneglect across Two Experiments: A Failed Internal Replication. Front Psychol. 2017, 8, 952. [Google Scholar] [CrossRef]
- Bergmann, T.O.; Groppa, S.; Seeger, M.; Mölle, M.; Marshall, L.; Siebner, H.R. Acute Changes in Motor Cortical Excitability During Slow Oscillatory and Constant Anodal Transcranial Direct Current Stimulation. J. Neurophysiol. 2009, 102, 2303–2311. [Google Scholar] [CrossRef] [PubMed]
- Fresnoza, S.; Christova, M.; Feil, T.; Gallasch, E.; Korner, C.; Zimmer, U.; Ischebeck, A. The effects of transcranial alternating current stimulation (tACS) at individual alpha peak frequency (iAPF) on motor cortex excitability in young and elderly adults. Exp. Brain Res. 2018, 236, 2573–2588. [Google Scholar] [CrossRef] [PubMed]
- Stecher, H.I.; Pollok, T.M.; Strüber, D.; Sobotka, F.; Herrmann, C.S. Ten Minutes of α-tACS and Ambient Illumination Independently Modulate EEG α-Power. Front. Hum. Neurosci. 2017, 11, 257. [Google Scholar] [CrossRef] [PubMed]
- Zaehle, T.; Rach, S.; Herrmann, C.S. Transcranial Alternating Current Stimulation Enhances Individual Alpha Activity in Human EEG. PLoS ONE 2010, 5, e13766. [Google Scholar] [CrossRef] [PubMed]
- Zaehle, T.; Sandmann, P.; Thorne, J.D.; Jäncke, L.; Herrmann, C.S. Transcranial direct current stimulation of the prefrontal cortex modulates working memory performance: Combined behavioural and electrophysiological evidence. BMC Neurosci. 2011, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Wirth, M.; Rahman, R.A.; Kuenecke, J.; Koenig, T.; Horn, H.; Sommer, W.; Dierks, T. Effects of transcranial direct current stimulation (tDCS) on behaviour and electrophysiology of language production. Neuropsychology 2011, 49, 3989–3998. [Google Scholar] [CrossRef] [PubMed]
- Lustenberger, C.; Boyle, M.R.; Alagapan, S.; Mellin, J.M.; Vaughn, B.V.; Fröhlich, F. Feedback-Controlled Transcranial Alternating Current Stimulation Reveals a Functional Role of Sleep Spindles in Motor Memory Consolidation. Curr. Boil. 2016, 26, 2127–2136. [Google Scholar] [CrossRef]
- Marshall, L.; Helgadóttir, H.; Mölle, M.; Born, J. Boosting slow oscillations during sleep potentiates memory. Nature 2006, 444, 610–613. [Google Scholar] [CrossRef]
- Neuling, T.; Rach, S.; Herrmann, C.S. Orchestrating neuronal networks: Sustained after-effects of transcranial alternating current stimulation depend upon brain states. Front. Hum. Neurosci. 2013, 7, 161. [Google Scholar] [CrossRef]
- Hoy, K.E.; Bailey, N.; Arnold, S.; Windsor, K.; John, J.; Daskalakis, Z.J.; Fitzgerald, P.B. The effect of γ-tACS on working memory performance in healthy controls. Brain Cogn. 2015, 101, 51–56. [Google Scholar] [CrossRef]
- Craddock, M.; Klepousniotou, E.; El-Deredy, W.; Poliakoff, E.; Lloyd, D. Transcranial alternating current stimulation at 10Hz modulates response bias in the Somatic Signal Detection Task. Int. J. Psychophysiol. 2019, 135, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Pahor, A.; Jausovec, N. The effects of theta transcranial alternating current stimulation (tACS) on fluid intelligence. Int. J. Psychophysiol. 2014, 93, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Paßmann, S.; Külzow, N.; Ladenbauer, J.; Antonenko, D.; Grittner, U.; Tamm, S.; Flöel, A. Boosting Slow Oscillatory Activity Using tDCS during Early Nocturnal Slow Wave Sleep Does Not Improve Memory Consolidation in Healthy Older Adults. Brain Stimul. 2016, 9, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Sahlem, G.L.; Badran, B.W.; Halford, J.J.; Williams, N.R.; Korte, J.E.; Leslie, K.; Strachan, M.; Breedlove, J.L.; Runion, J.; Bachman, D.L.; et al. Oscillating Square Wave Transcranial Direct Current Stimulation (tDCS) Delivered During Slow Wave Sleep Does Not Improve Declarative Memory More Than Sham: A Randomized Sham Controlled Crossover Study. Brain Stimul. 2015, 8, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Lopez, A.; Eggert, T.; Dorn, H.; Danker-Hopfe, H. Slow oscillatory transcranial direct current stimulation (so-tDCS) during slow wave sleep has no effects on declarative memory in healthy young subjects. Brain Stimul. 2019, 12, 948–958. [Google Scholar] [CrossRef]
- Antonenko, D.; Diekelmann, S.; Olsen, C.; Born, J.; Mölle, M. Napping to renew learning capacity: Enhanced encoding after stimulation of sleep slow oscillations. Eur. J. Neurosci. 2013, 37, 1142–1151. [Google Scholar] [CrossRef]
- Kirov, R.; Weiss, C.; Siebner, H.R.; Born, J.; Marshall, L. Slow oscillation electrical brain stimulation during waking promotes EEG theta activity and memory encoding. Proc. Natl. Acad. Sci. USA 2009, 106, 15460–15465. [Google Scholar] [CrossRef]
- Ladenbauer, J.; Külzow, N.; Passmann, S.; Antonenko, D.; Grittner, U.; Tamm, S.; Flöel, A. Brain stimulation during an afternoon nap boosts slow oscillatory activity and memory consolidation in older adults. NeuroImage 2016, 142, 311–323. [Google Scholar] [CrossRef]
- Ladenbauer, J.; Ladenbauer, J.; Külzow, N.; De Boor, R.; Avramova, E.; Grittner, U.; Flöel, A. Promoting Sleep Oscillations and Their Functional Coupling by Transcranial Stimulation Enhances Memory Consolidation in Mild Cognitive Impairment. J. Neurosci. 2017, 37, 7111–7124. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Munz, M.; Göder, R.; Wilhelm, I.; Korr, K.; Vahl, W.; Wiesner, C.D.; Baving, L. Transcranial Oscillatory Direct Current Stimulation during Sleep Improves Declarative Memory Consolidation in Children with Attention-deficit/hyperactivity Disorder to a Level Comparable to Healthy Controls. Brain Stimul. 2014, 7, 793–799. [Google Scholar] [CrossRef]
- Marshall, L.; Kirov, R.; Brade, J.; Mölle, M.; Born, J. Transcranial Electrical Currents to Probe EEG Brain Rhythms and Memory Consolidation during Sleep in Humans. PLoS ONE 2011, 6, e16905. [Google Scholar] [CrossRef] [PubMed]
- Munz, M.T.; Prehn-Kristensen, A.; Thielking, F.; Mölle, M.; Göder, R.; Baving, L. Slow oscillating transcranial direct current stimulation during non-rapid eye movement sleep improves behavioral inhibition in attention-deficit/hyperactivity disorder. Front. Cell. Neurosci. 2015, 9, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerberg, C.E.; Florczak, S.M.; Weintraub, S.; Mesulam, M.-M.; Marshall, L.; Zee, P.C.; Paller, K.A. Memory improvement via slow-oscillatory stimulation during sleep in older adults. Neurobiol. Aging 2015, 36, 2577–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-Z.; Lu, M.-K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity induced by non-invasive transcranial brain stimulation: A position paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef]
- Jacobson, L.; Koslowsky, M.; Lavidor, M. tDCS polarity effects in motor and cognitive domains: A meta-analytical review. Exp. Brain Res. 2012, 216, 1–10. [Google Scholar] [CrossRef]
- Zhu, F.; Maxwell, J.; Hu, Y.; Zhang, Z.; Lam, W.; Poolton, J.; Masters, R. EEG activity during the verbal-cognitive stage of motor skill acquisition. Boil. Psychol. 2010, 84, 221–227. [Google Scholar] [CrossRef]
- Makeig, S.; AnlloVento, L.; Jung, P.; Bell, A.J.; Sejnowski, T.J.; Hillyard, S.A. Independent component analysis of event-related potentials during a selective attention task. Psychophysiology 1996, 33, S58. [Google Scholar]
- Haegens, S.; Cousijn, H.; Wallis, G.; Harrison, P.J.; Nobre, A.C. Inter- and intra-individual variability in alpha peak frequency. NeuroImage 2014, 92, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Grandy, T.H.; Werkle-Bergner, M.; Chicherio, C.; Schmiedek, F.; Lövdén, M.; Lindenberger, U. Peak individual alpha frequency qualifies as a stable neurophysiological trait marker in healthy younger and older adults. Psychophysiology 2013, 50, 570–582. [Google Scholar] [CrossRef] [Green Version]
- Olbrich, S.; Mulert, C.; Karch, S.; Trenner, M.; Leicht, G.; Pogarell, O.; Hegerl, U. EEG-vigilance and BOLD effect during simultaneous EEG/fMRI measurement. NeuroImage 2009, 45, 319–332. [Google Scholar] [CrossRef]
- Mierau, A.; Klimesch, W.; Lefebvre, J. State-dependent alpha peak frequency shifts: Experimental evidence, potential mechanisms and functional implications. Neuroscience 2017, 360, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.P.; Brakefield, T.; Morgan, A.; Hobson, J.A.; Stickgold, R. Practice with sleep makes perfect: Sleep-dependent motor skill learning. Neuron 2002, 35, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Hoddes, E.; Zarcone, V.; Dement, W. Development and use of stanford sleepiness scale (SSS). Psychophysiology 1972, 9, 150. [Google Scholar]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and validation of brief measures of positive and negative affect: The PANAS scales. J. Pers. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Mölle, M.; Marshall, L.; Fehm, H.L.; Born, J. EEG theta synchronization conjoined with alpha desynchronization indicate intentional encoding. Eur. J. Neurosci. 2002, 15, 923–928. [Google Scholar] [CrossRef]
- Marshall, L.; Born, J. Brain stimulation during sleep. Sleep Med. Clin. 2011, 6, 85–95. [Google Scholar] [CrossRef]
- Kanai, R.; Chaieb, L.; Antal, A.; Walsh, V.; Paulus, W. Frequency-Dependent Electrical Stimulation of the Visual Cortex. Curr. Boil. 2008, 18, 1839–1843. [Google Scholar] [CrossRef] [Green Version]
- Hutcheon, B.; Yarom, Y. Resonance, oscillation and the intrinsic frequency preferences of neurons. Trends Neurosci. 2000, 23, 216–222. [Google Scholar] [CrossRef]
- Vossen, A.; Gross, J.; Thut, G. Alpha Power Increase After Transcranial Alternating Current Stimulation at Alpha Frequency (alpha-tACS) Reflects Plastic Changes Rather Than Entrainment. Brain Stimul. 2015, 8, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Nunez, P.L. Generation of human EEG by a combination of long and short range neocortical interactions. Brain Topogr. 1989, 1, 199–215. [Google Scholar] [CrossRef]
- Goldman, R.I.; Stern, J.M.; Engel, J., Jr.; Cohen, M.S. Simultaneous EEG and fMRI of the alpha rhythm. Neuroreport 2002, 13, 2487. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.W.; Crunelli, V. Thalamic Mechanisms of EEG Alpha Rhythms and Their Pathological Implications. Neuroscience 2005, 11, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Aspart, F.; Remme, M.W.H.; Obermayer, K. Differential polarization of cortical pyramidal neuron dendrites through weak extracellular fields. PLoS Comput. Boil. 2018, 14, e1006124. [Google Scholar] [CrossRef] [PubMed]
- Miranda, P.C.; Faria, P.; Hallett, M. What does the ratio of injected current to electrode area tell us about current density in the brain during tDCS? Clin. Neurophysiol. 2009, 120, 1183–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozen, S.; Sirota, A.; Belluscio, M.A.; Anastassiou, C.A.; Stark, E.; Koch, C.; Buzsáki, G. Transcranial electric stimulation entrains cortical neuronal populations in rats. J. Neurosci. 2010, 30, 11476–11485. [Google Scholar] [CrossRef] [PubMed]
- Smit, C.M.; Wright, M.J.; Hansell, N.K.; Geffen, G.M.; Martin, N.G. Genetic variation of individual alpha frequency (IAF) and alpha power in a large adolescent twin sample. Int. J. Psychophysiol. 2006, 61, 235–243. [Google Scholar] [CrossRef]
- Schreckenberger, M.; Lange-Asschenfeld, C.; Lochmann, M.; Mann, K.; Siessmeier, T.; Buchholz, H.-G.; Bartenstein, P.; Gründer, G. The thalamus as the generator and modulator of EEG alpha rhythm: A combined PET/EEG study with lorazepam challenge in humans. NeuroImage 2004, 22, 637–644. [Google Scholar] [CrossRef]
- D’Atri, A.; De Simoni, E.; Gorgoni, M.; Ferrara, M.; Ferlazzo, F.; Rossini, P.; De Gennaro, L. Electrical stimulation of the frontal cortex enhances slow-frequency EEG activity and sleepiness. Neuroscience 2016, 324, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Albouy, G.; Fogel, S.; King, B.R.; LaVenture, S.; Benali, H.; Karni, A.; Carrier, J.; Robertson, E.M.; Doyon, J. Maintaining vs. enhancing motor sequence memories: Respective roles of striatal and hippocampal systems. NeuroImage 2015, 108, 423–434. [Google Scholar] [CrossRef]
- Brashers-Krug, T.; Shadmehr, R.; Bizzi, E. Consolidation in human motor memory. Nat. 1996, 382, 252–255. [Google Scholar] [CrossRef]
- Kuo, H.-I.; Bikson, M.; Datta, A.; Minhas, P.; Paulus, W.; Kuo, M.-F.; Nitsche, M.A. Comparing Cortical Plasticity Induced by Conventional and High-Definition 4 × 1 Ring tDCS: A Neurophysiological Study. Brain Stimul. 2013, 6, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Albert, N.B.; Robertson, E.M.; Miall, R.C. The resting human brain and motor learning. Curr. Boil. 2009, 19, 1023–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyon, J.; Penhune, V.; Ungerleider, L.G. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychology 2003, 41, 252–262. [Google Scholar] [CrossRef]
- Krakauer, J.W.; Shadmehr, R. Consolidation of motor memory. Trends Neurosci. 2006, 29, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, M.D.; Agam, Y.; Selvadurai, C.; Nagy, A.; Vangel, M.; Tucker, M.; Robertson, E.M.; Stickgold, R.; Manoach, D.S. Resting state connectivity immediately following learning correlates with subsequent sleep-dependent enhancement of motor task performance. NeuroImage 2014, 102, 666–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annarumma, L.; D’Atri, A.; Alfonsi, V.; De Gennaro, L. The Efficacy of Transcranial Current Stimulation Techniques to Modulate Resting-State EEG, to Affect Vigilance and to Promote Sleepiness. Brain Sci. 2018, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Ben-Simon, E.; Podlipsky, I.; Arieli, A.; Zhdanov, A.; Hendler, T. Never Resting Brain: Simultaneous Representation of Two Alpha Related Processes in Humans. PLoS ONE 2008, 3, e3984. [Google Scholar] [CrossRef] [Green Version]
- Bönstrup, M.; Iturrate, I.; Thompson, R.; Cruciani, G.; Censor, N.; Cohen, L.G. A Rapid Form of Offline Consolidation in Skill Learning. Curr. Boil. 2019, 29, 1346–1351. [Google Scholar] [CrossRef]
- Humiston, G.B.; Wamsley, E.J. A brief period of eyes-closed rest enhances motor skill consolidation. Neurobiol. Learn. Mem. 2018, 155, 1–6. [Google Scholar] [CrossRef]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Datta, A.; Bikson, M.; Fregni, F. Transcranial direct current stimulation in patients with skull defects and skull plates: High-resolution computational FEM study of factors altering cortical current flow. NeuroImage 2010, 52, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, P.C.; Mölle, M.; Marshall, L.; Moelle, M. Efficacy of slow oscillatory-transcranial direct current stimulation on EEG and memory—Contribution of an inter-individual factor. Eur. J. Neurosci. 2018, 47, 812–823. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koo-Poeggel, P.; Böttger, V.; Marshall, L. Distinct Montages of Slow Oscillatory Transcranial Direct Current Stimulation (so-tDCS) Constitute Different Mechanisms during Quiet Wakefulness. Brain Sci. 2019, 9, 324. https://doi.org/10.3390/brainsci9110324
Koo-Poeggel P, Böttger V, Marshall L. Distinct Montages of Slow Oscillatory Transcranial Direct Current Stimulation (so-tDCS) Constitute Different Mechanisms during Quiet Wakefulness. Brain Sciences. 2019; 9(11):324. https://doi.org/10.3390/brainsci9110324
Chicago/Turabian StyleKoo-Poeggel, Ping, Verena Böttger, and Lisa Marshall. 2019. "Distinct Montages of Slow Oscillatory Transcranial Direct Current Stimulation (so-tDCS) Constitute Different Mechanisms during Quiet Wakefulness" Brain Sciences 9, no. 11: 324. https://doi.org/10.3390/brainsci9110324