
Impact of Spinal Manipulation on Cortical Drive to Upper and Lower Limb Muscles

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Protocol
2.2. EEG and EMG Acquisition and Analysis
2.3. TMS
2.4. F Waves
2.5. MRCP
2.5.1. Signal Processing
2.5.2. Morphology Extraction (MRCP)
2.6. Interventions
2.6.1. Spinal Manipulation Intervention
2.6.2. Control Intervention
2.7. Statistical Analysis
3. Results
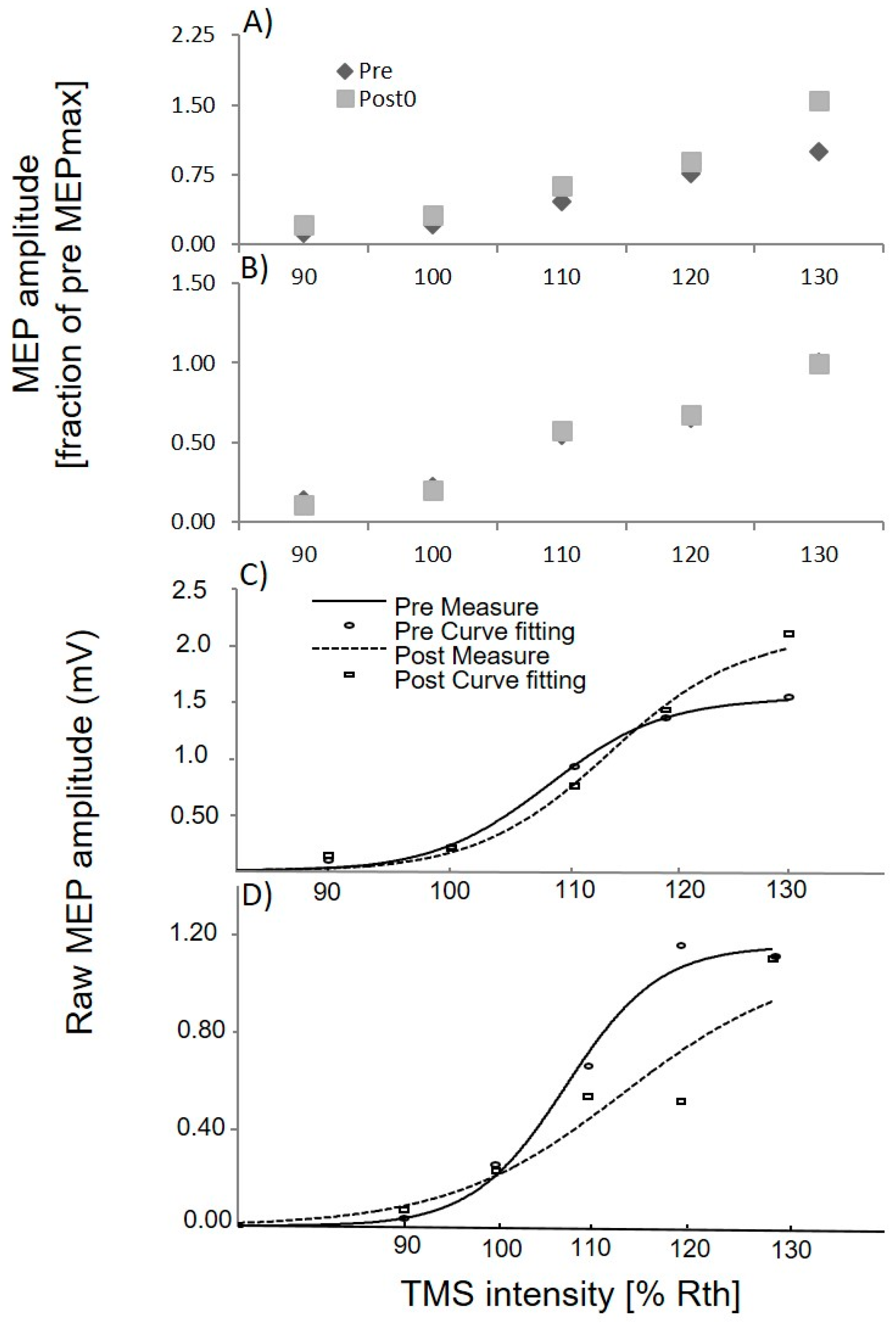
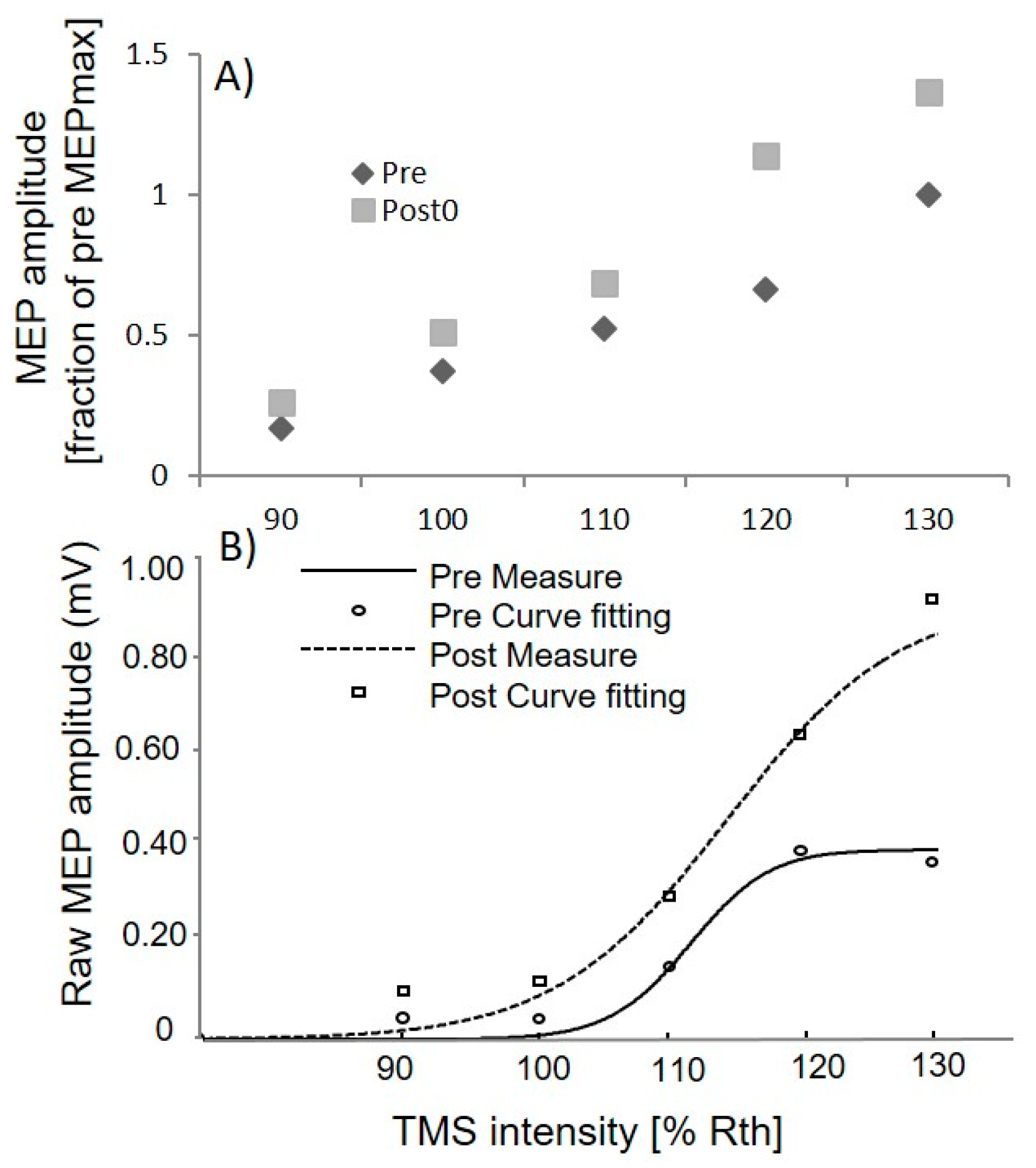
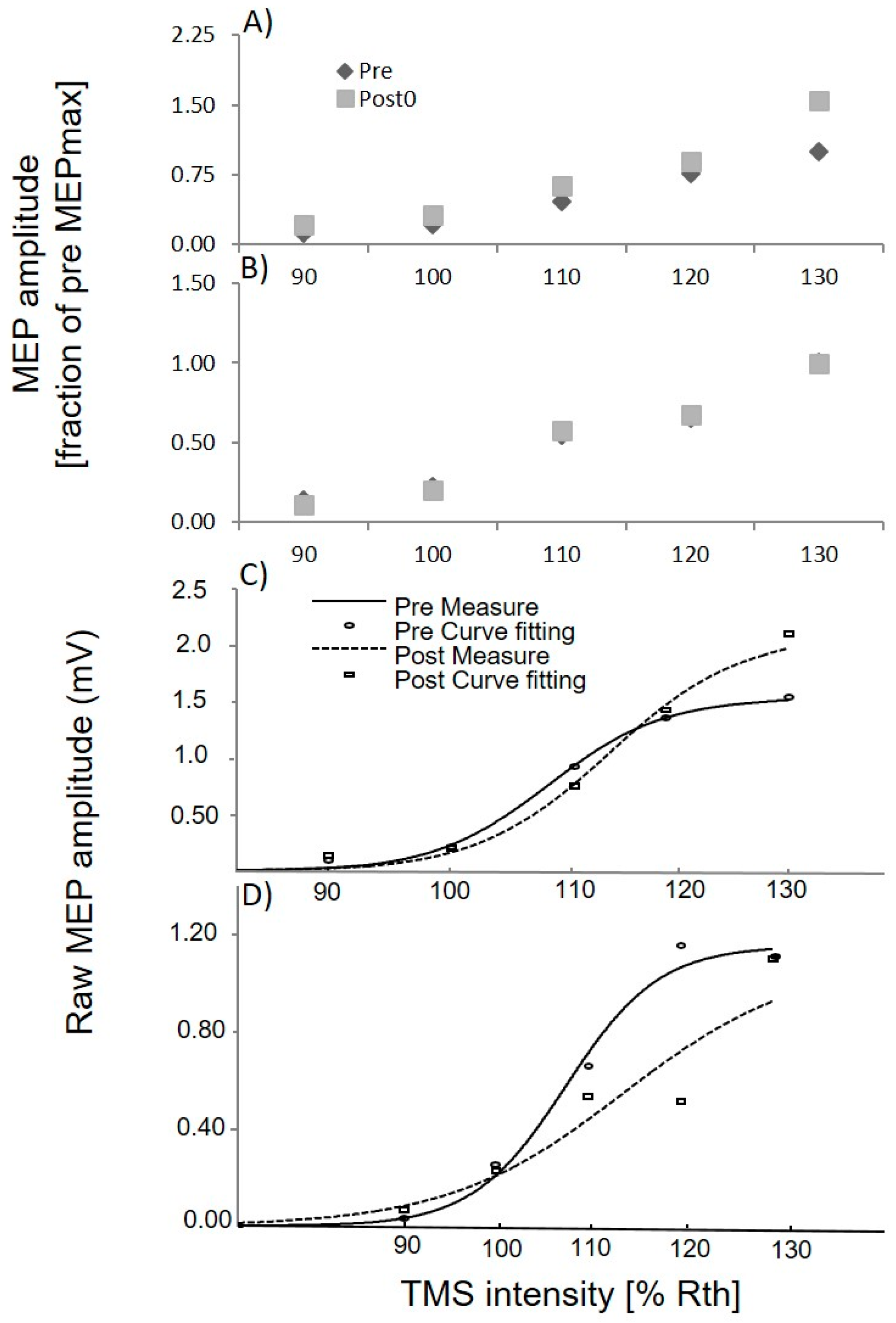
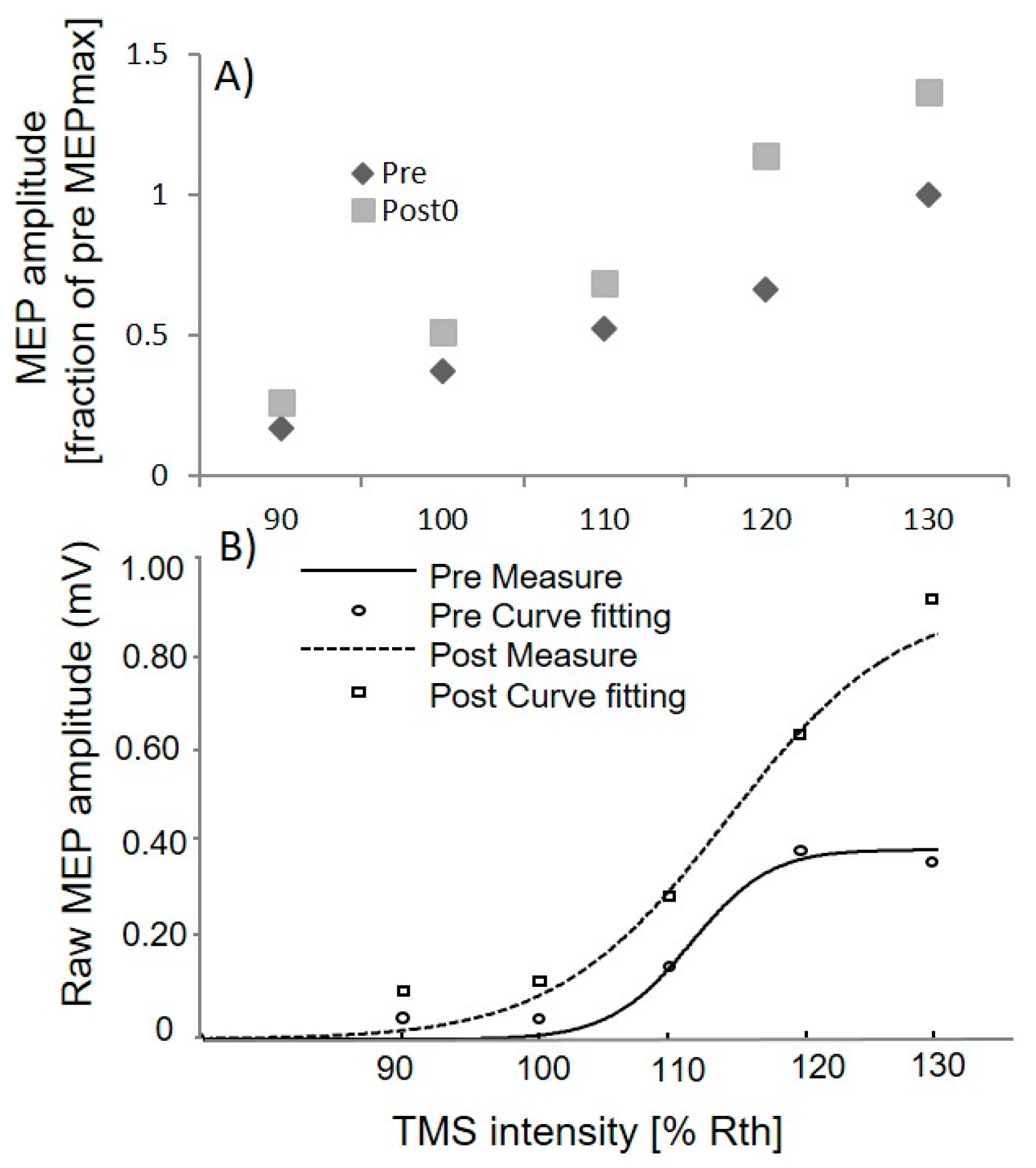
3.1. Upper Limb TMS
3.2. Upper Limb F and M Waves
3.3. Lower Limb TMS
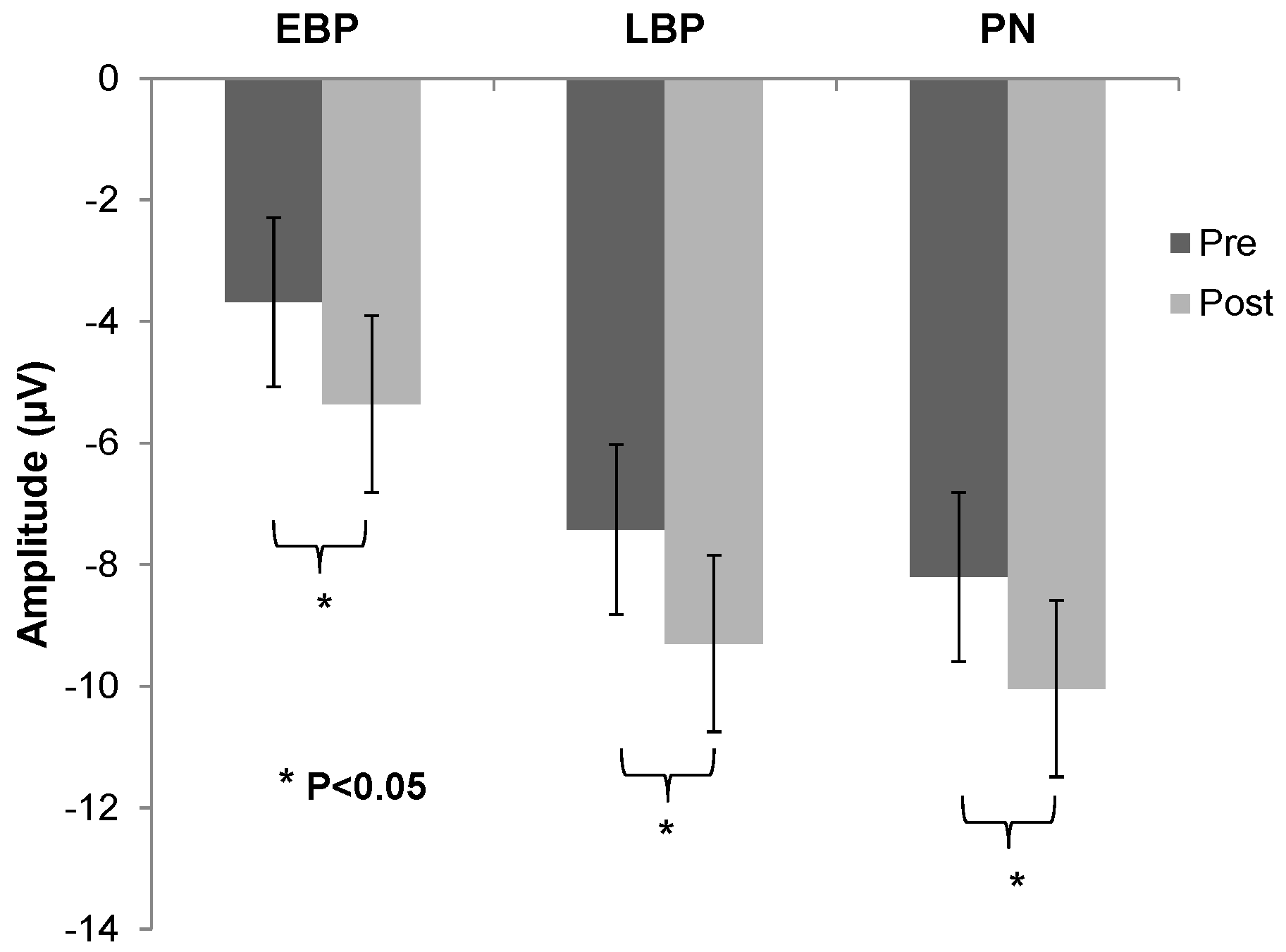
3.4. Lower Limb MRCP Morphology
4. Discussion
4.1. Neuroplastic Changes
4.2. Cortical vs. Spinal Changes
4.3. Limitation
4.4. Functional Implication
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haavik, H.; Murphy, B. The role of spinal manipulation in addressing disordered sensorimotor integration and altered motor control. J. Electromyogr. Kinesiol. 2012, 22, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Bronfort, G.; Evans, R.; Anderson, A.V.; Svendsen, K.H.; Bracha, Y.; Grimm, R.H. Spinal manipulation, medication, or home exercise with advice for acute and subacute neck pain: A randomized trial. Ann. Intern. Med. 2012, 156, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bronfort, G.; Haas, M.; Evans, R.L.; Bouter, L.M. Efficacy of spinal manipulation and mobilization for low back pain and neck pain: A systematic review and best evidence synthesis. Spine J. 2004, 4, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, J.J.; Schwieterman, B.; Columber, K.; Knupp, D.; Shaub, L.; Cook, C.E. Effectiveness of physical therapist administered spinal manipulation for the treatment of low back pain: A systematic review of the literature. Int. J. Sports Phys. Ther. 2012, 7, 647–662. [Google Scholar] [PubMed]
- Gross, A.; Miller, J.; D’Sylva, J.; Burnie, S.J.; Goldsmith, C.H.; Graham, N.; Haines, T.; Bronfort, G.; Hoving, J.L. Manipulation or mobilisation for neck pain: A cochrane review. Man. Ther. 2010, 15, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Marshall, P.; Murphy, B. The effect of sacroiliac joint manipulation on feed-forward activation times of the deep abdominal musculature. J. Manip. Physiol. Ther. 2006, 29, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Haavik-Taylor, H.; Murphy, B. Cervical spine manipulation alters sensorimotor integration: A somatosensory evoked potential study. Clin. Neurophysiol. 2007, 118, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Haavik-Taylor, H.; Murphy, B. Transient modulation of intracortical inhibition following spinal manipulation. Chiropr. J. Aust. 2007, 37, 106–116. [Google Scholar]
- Taylor, H.H.; Murphy, B. Altered sensorimotor integration with cervical spine manipulation. J. Manip. Physiol. Ther. 2008, 31, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.H.; Murphy, B. Altered central integration of dual somatosensory input after cervical spine manipulation. J. Manip. Physiol. Ther. 2010, 33, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Haavik Taylor, H.; Murphy, B. The Effects of spinal manipulation on central integration of dual somatosensory input observed after motor training: A crossover study. J. Manip. Physiol. Ther. 2010, 33, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Haavik, H.; Murphy, B. Subclinical neck pain and the effects of cervical manipulation on elbow joint position sense. J. Manip. Physiol. Ther. 2011, 34, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Allison, T.; McCarthy, G.; Wood, C.C.; Williamson, P.D.; Spencer, D.D. Human Cortical potentials evoked by stimulation of the median nerve. II. Cytoarchitectonic areas generating long-latency activity. J. Neurophysiol. 1989, 62, 711–722. [Google Scholar] [PubMed]
- Allison, T.; McCarthy, G.; Wood, C.C.; Darcey, T.M.; Spencer, D.D.; Williamson, P.D.; Spencer, D.D. Human cortical potentials evoked by stimulation of the median nerve. II. Cytoarchitectonic areas generating short-latency activity. J. Neurophysiol. 1989, 62, 694–710. [Google Scholar] [PubMed]
- Allison, T.; Wood, C.C.; McCarthy, G.; Spencer, D.D. Cortical somatosensory evoked potentials. II. Effects of excision of somatosensory or motor cortex in humans and monkeys. J. Neurophysiol. 1991, 66, 64–82. [Google Scholar] [PubMed]
- Kanovsky, P.; Bares, M.; Rektor, I. The selective gating of the N30 cortical component of the somatosensory evoked potentials of median nerve is different in the mesial and dorsolateral frontal cortex: Evidence from intracerebral recordings. Clin. Neurophysiol. 2003, 114, 981–991. [Google Scholar] [CrossRef]
- Mauguiere, F.; Desmedt, J.E.; Courjon, J. Astereognosis and dissociated loss of frontal or parietal components of somatosensory evoked potentials in hemispheric lesions. detailed correlations with clinical signs and computerized tomographic scanning. Brain 1983, 106, 271–311. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Gigli, G.L.; Marciani, M.G.; Zarola, F.; Caramia, M. Non-invasive evaluation of input-output characteristics of sensorimotor cerebral areas in healthy humans. Electroencephalogr. Clin. Neurophysiol. 1987, 68, 88–100. [Google Scholar] [CrossRef]
- Rossini, P.M.; Babiloni, F.; Bernardi, G.; Cecchi, L.; Johnson, P.B.; Malentacca, A.; Stanzione, P.; Urbano, A. Abnormalities of Short-latency somatosensory evoked potentials in parkinsonian patients. Electroencephalogr. Clin. Neurophysiol. 1989, 74, 277–289. [Google Scholar] [CrossRef]
- Waberski, T.D.; Buchner, H.; Perkuhn, M.; Gobbele, R.; Wagner, M.; Kucker, W.; Silny, J. N30 and the effect of explorative finger movements: A model of the contribution of the motor cortex to early somatosensory potentials. Clin. Neurophysiol. 1999, 110, 1589–1600. [Google Scholar] [CrossRef]
- Rossi, S.; della Volpe, R.; Ginanneschi, F.; Ulivelli, M.; Bartalini, S.; Spidalieri, R.; Rossi, A. Early somatosensory processing during tonic muscle pain in humans: Relation to loss of proprioception and motor ‘defensive’ strategies. Clin. Neurophysiol. 2003, 114, 1351–1358. [Google Scholar] [CrossRef]
- Lelic, D.; Niazi, I.K.; Holt, K.; Jochumsen, M.; Dremstrup, K.; Yielder, P.; Murphy, B.; Drewes, A.M.; Haavik, H. Manipulation of dysfunctional spinal joints affects sensorimotor integration in the prefrontal cortex: A brain source localization study. Neural Plast. 2016, 2016, 3704964. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.R.; Haavik, H.; Lee, A.C.L.; Murphy, B.; Elley, C.R. Effectiveness of chiropractic care to improve sensorimotor function associated with falls risk in older people: A randomized controlled trial. J. Manip. Physiol. Ther. 2016, 39, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Niazi, I.K.; Türker, K.S.; Flavel, S.; Kinget, M.; Duehr, J.; Haavik, H. Changes in H-reflex and V-waves following spinal manipulation. Exp. Brain Res. 2015, 233, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Schlaug, G.; Renga, V.; Nair, D. Transcranial direct current stimulation in stroke recovery. Arch. Neurol. 2008, 65, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Mansur, C.G.; Fregni, F.; Boggio, P.S.; Riberto, M.; Gallucci-Neto, J.; Santos, C.M.; Wagner, T.; Rigonatti, S.P.; Marcolin, M.A.; Pascual-Leone, A. A Sham stimulation-controlled trial of rTMS of the unaffected hemisphere in stroke patients. Neurology 2005, 64, 1802–1804. [Google Scholar] [CrossRef] [PubMed]
- Hummel, F.; Celnik, P.; Giraux, P.; Floel, A.; Wu, W.-H.; Gerloff, C.; Cohen, L.G. Effects of non-invasive cortical stimulation on skilled motor function in chronic stroke. Brain 2005, 128, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M.; Jahanshahi, M. The Bereitschaftspotential: Movement-Related Cortical Potentials; Springer Science & Business Media: Berlin, Germany, 2003. [Google Scholar]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety of TMS consensus group. safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Nicholson, L.L.; Adams, R.D. Cervical range of motion associations with subclinical neck pain. Spine 2004, 29, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Nicholson, L.L.; Adams, R.D.; Bae, S.-S. Proprioception and rotation range sensitization associated with subclinical neck pain. Spine 2005, 30, E60–E67. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Wang, J.-D.; Yao, G.; Wang, S.-F. Association between cervicocephalic kinesthetic sensibility and frequency of subclinical neck pain. Man. Ther. 2008, 13, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.J.; Riek, S.; Carson, R.G. Reliability of the input-output properties of the cortico-spinal pathway obtained from transcranial magnetic and electrical stimulation. J. Neurosci. Methods 2001, 112, 193–202. [Google Scholar] [CrossRef]
- Malcolm, M.P.; Triggs, W.J.; Light, K.E.; Shechtman, O.; Khandekar, G.; Gonzalez Rothi, L.J. Reliability of motor cortex transcranial magnetic stimulation in four muscle representations. Clin. Neurophysiol. 2006, 117, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Kamen, G. Reliability of motor-evoked potentials during resting and active contraction conditions. Med. Sci. Sports Exerc. 2004, 36, 1574–1579. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.A. AAEM Minimonograph #13: H reflexes and F waves: Physiology and clinical indications. Muscle Nerve 1992, 15, 1223–1233. [Google Scholar] [PubMed]
- Panayiotopoulos, C.P.; Chroni, E. F-Waves in clinical neurophysiology: A review, methodological issues and overall value in peripheral neuropathies. Electroencephalogr. Clin. Neurophysiol. 1996, 101, 365–374. [Google Scholar] [CrossRef]
- Taniguchi, M.H.; Hayes, J.; Rodriguez, A.A. Reliability determination of F mean response latency. Arch. Phys. Med. Rehabil. 1993, 74, 1139–1143. [Google Scholar] [PubMed]
- Slobounov, S.M.; Ray, W.J. Movement-related potentials with reference to isometric force output in discrete and repetitive tasks. Exp. Brain Res. 1998, 123, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, O.F.; Nielsen, K.D.; Voigt, M. Relationship between plantar-flexor torque generation and the magnitude of the movement-related potentials. Exp. Brain Res. 2005, 160, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Hestbaek, L.; Leboeuf-Yde, C. Are Chiropractic Tests for the lumbo-pelvic spine reliable and valid? A systematic critical literature review. J. Manip. Physiol. Ther. 2000, 23, 258–275. [Google Scholar] [CrossRef]
- Fryer, G.; Morris, T.; Gibbons, P. Paraspinal muscles and intervertebral dysfunction: Part one. J. Manip. Physiol. Ther. 2004, 27, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Jull, G.; Bogduk, N.; Marsland, A. The Accuracy of manual diagnosis for cervical zygapophysial joint pain syndromes. Med. J. Aust. 1988, 148, 233–236. [Google Scholar] [PubMed]
- Hubka, M.J.; Phelan, S.P. Interexaminer reliability of palpation for cervical spine tenderness. J. Manip. Physiol. Ther. 1994, 17, 591–595. [Google Scholar]
- Rheault, W.; Albright, B.; Beyers, C.; Franta, M.; Johnson, A.; Skowronek, M.; Dougherty, J. Intertester reliability of the cervical range of motion device. J. Orthop. Sports Phys. Ther. 1992, 15, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Youdas, J.W.; Garrett, T.R.; Suman, V.J.; Bogard, C.L.; Hallman, H.O.; Carey, J.R. Normal range of motion of the cervical spine: An initial goniometric study. Phys. Ther. 1992, 72, 770–780. [Google Scholar] [PubMed]
- Strender, L.E.; Sjoblom, A.; Sundell, K.; Ludwig, R.; Taube, A. Interexaminer reliability in physical examination of patients with low back pain. Spine 1997, 22, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W.; Read, L.J.; Conway, P.J.; Shaw, L.D.; McEwen, M.C. Reliability of motion palpation procedures to detect sacroiliac joint fixations. J. Manip. Physiol. Ther. 1989, 12, 86–92. [Google Scholar]
- Potter, N.A.; Rothstein, J.M. Intertester reliability for selected clinical tests of the sacroiliac joint. Phys. Ther. 1985, 65, 1671–1675. [Google Scholar] [CrossRef] [PubMed]
- Flynn, T.; Fritz, J.; Whitman, J.; Wainner, R.; Magel, J.; Rendeiro, D.; Butler, B.; Garber, M.; Allison, S. A clinical prediction rule for classifying patients with low back pain who demonstrate short-term improvement with spinal manipulation. Spine 2002, 27, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Hessell, B.W.; Herzog, W.; Conway, P.J.; McEwen, M.C. Experimental measurement of the force exerted during spinal manipulation using the thompson technique. J. Manip. Physiol. Ther. 1990, 13, 448–453. [Google Scholar]
- Herzog, W. Mechanical, Physiologic, and Neuromuscular Considerations of Chiropractic Treatment; Lawrence, D., Ed.; Mosby-Year Book: New York, NY, USA, 1996. [Google Scholar]
- Herzog, W.; Conway, P.J.; Zhang, Y.T.; Gal, J.; Guimaraes, A.C. Reflex responses associated with manipulative treatments on the thoracic spine: A pilot study. J. Manip. Physiol. Ther. 1995, 18, 233–236. [Google Scholar]
- Pickar, J.G.; Wheeler, J.D. Response of Muscle proprioceptors to spinal manipulative-like loads in the anesthetized cat. J. Manip. Physiol. Ther. 2001, 24, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Devanne, H.; Lavoie, B.A.; Capaday, C. Input-output properties and gain changes in the human corticospinal pathway. Exp. Brain Res. 1997, 114, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Groppa, S.; Oliviero, A.; Eisen, A.; Quartarone, A.; Cohen, L.G.; Mall, V.; Kaelin-Lang, A.; Mima, T.; Rossi, S.; Thickbroom, G.W.; et al. A practical guide to diagnostic transcranial magnetic stimulation: Report of an IFCN committee. Clin. Neurophysiol. 2012, 123, 858–882. [Google Scholar] [CrossRef] [PubMed]
- Deecke, L.; Kornhuber, H.H. An electrical sign of participation of the mesial ‘supplementary’ motor cortex in human voluntary finger movement. Brain Res. 1978, 159, 473–476. [Google Scholar] [CrossRef]
- Deecke, L. Bereitschaftspotential as an indicator of movement preparation in supplementary motor area and motor cortex. Ciba Found. Symp. 1987, 132, 231–250. [Google Scholar] [PubMed]
- Deecke, L.; Lang, W. Generation of movement-related potentials and fields in the supplementary sensorimotor area and the primary motor area. Adv. Neurol. 1996, 70, 127–146. [Google Scholar] [PubMed]
- Praamstra, P.; Stegeman, D.F.; Horstink, M.W.; Cools, A.R. Dipole source analysis suggests selective modulation of the supplementary motor area contribution to the readiness potential. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 468–477. [Google Scholar] [CrossRef]
- Shibasaki, H.; Hallett, M. What is the bereitschaftspotential? Clin. Neurophysiol. 2006, 117, 2341–2356. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; MacKinnon, C.D. The effect of temporal accuracy constraints on movement-related potentials. Exp. Brain Res. 2009, 194, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Deecke, L.; Scheid, P.; Kornhuber, H.H. Distribution of readiness potential, pre-motion positivity, and motor potential of the human cerebral cortex preceding voluntary finger movements. Exp. Brain Res. 1969, 7, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Deecke, L.; Grozinger, B.; Kornhuber, H.H. Voluntary finger movement in man: cerebral potentials and theory. Biol. Cybern. 1976, 23, 99–119. [Google Scholar] [CrossRef] [PubMed]
- Kristeva, R.; Keller, E.; Deecke, L.; Kornhuber, H.H. Cerebral potentials preceding unilateral and simultaneous bilateral finger movements. Electroencephalogr. Clin. Neurophysiol. 1979, 47, 229–238. [Google Scholar] [CrossRef]
- Kutas, M.; Donchin, E. Preparation to respond as manifested by movement-related brain potentials. Brain Res. 1980, 202, 95–115. [Google Scholar] [CrossRef]
- Shibasaki, H.; Barrett, G.; Halliday, E.; Halliday, A.M. Components of the movement-related cortical potential and their scalp topography. Electroencephalogr. Clin. Neurophysiol. 1980, 49, 213–226. [Google Scholar] [CrossRef]
- Barrett, G.; Shibasaki, H.; Neshige, R. A Computer-assisted method for averaging movement-related cortical potentials with respect to EMG onset. Electroencephalogr. Clin. Neurophysiol. 1985, 60, 276–281. [Google Scholar] [CrossRef]
- Kornhuber, H.H.; Deecke, L. Changes in the brain potential in voluntary movements and passive movements in man: Readiness potential and reafferent potentials. Pflugers Arch. Gesamte Physiol. Menschen Tiere 1965, 284, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, H.G.J.; Costa, L.D.; Ritter, W. Topography of the human motor potential. Electroencephalogr. Clin. Neurophysiol. 1968, 25, 1–10. [Google Scholar] [CrossRef]
- Colebatch, J.G. Bereitschaftspotential and movement-related potentials: origin, significance, and application in disorders of human movement. Mov. Disord. 2007, 22, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Daligadu, J.; Haavik, H.; Yielder, P.C.; Baarbe, J.; Murphy, B. Alterations in cortical and cerebellar motor processing in subclinical neck pain patients following spinal manipulation. J. Manip. Physiol. Ther. 2013, 36, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.E. Basal Ganglia-thalamocortical circuits: Their role in control of movements. J. Clin. Neurophysiol. 1994, 11, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Rektor, I.; Kanovsky, P.; Bares, M.; Louvel, J.; Lamarche, M. Event-related potentials, CNV, readiness potential, and movement accompanying potential recorded from posterior thalamus in human subjects. A SEEG study. Neurophysiol. Clin. 2001, 31, 253–261. [Google Scholar] [CrossRef]
- Murphy, B.A.; Dawson, N.J.; Slack, J.R. sacroiliac joint manipulation decreases the H-reflex. Electromyogr. Clin. Neurophysiol. 1995, 35, 87–94. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haavik, H.; Niazi, I.K.; Jochumsen, M.; Sherwin, D.; Flavel, S.; Türker, K.S. Impact of Spinal Manipulation on Cortical Drive to Upper and Lower Limb Muscles. Brain Sci. 2017, 7, 2. https://doi.org/10.3390/brainsci7010002
Haavik H, Niazi IK, Jochumsen M, Sherwin D, Flavel S, Türker KS. Impact of Spinal Manipulation on Cortical Drive to Upper and Lower Limb Muscles. Brain Sciences. 2017; 7(1):2. https://doi.org/10.3390/brainsci7010002
Chicago/Turabian StyleHaavik, Heidi, Imran Khan Niazi, Mads Jochumsen, Diane Sherwin, Stanley Flavel, and Kemal S. Türker. 2017. "Impact of Spinal Manipulation on Cortical Drive to Upper and Lower Limb Muscles" Brain Sciences 7, no. 1: 2. https://doi.org/10.3390/brainsci7010002
APA StyleHaavik, H., Niazi, I. K., Jochumsen, M., Sherwin, D., Flavel, S., & Türker, K. S. (2017). Impact of Spinal Manipulation on Cortical Drive to Upper and Lower Limb Muscles. Brain Sciences, 7(1), 2. https://doi.org/10.3390/brainsci7010002