
Differential Influence of the Dorsal Premotor and Primary Somatosensory Cortex on Corticospinal Excitability during Kinesthetic and Visual Motor Imagery: A Low-Frequency Repetitive Transcranial Magnetic Stimulation Study


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample
2.2. EMG and Single-Pulse TMS
2.3. Procedure
2.4. Data Handling and Statistical Analysis
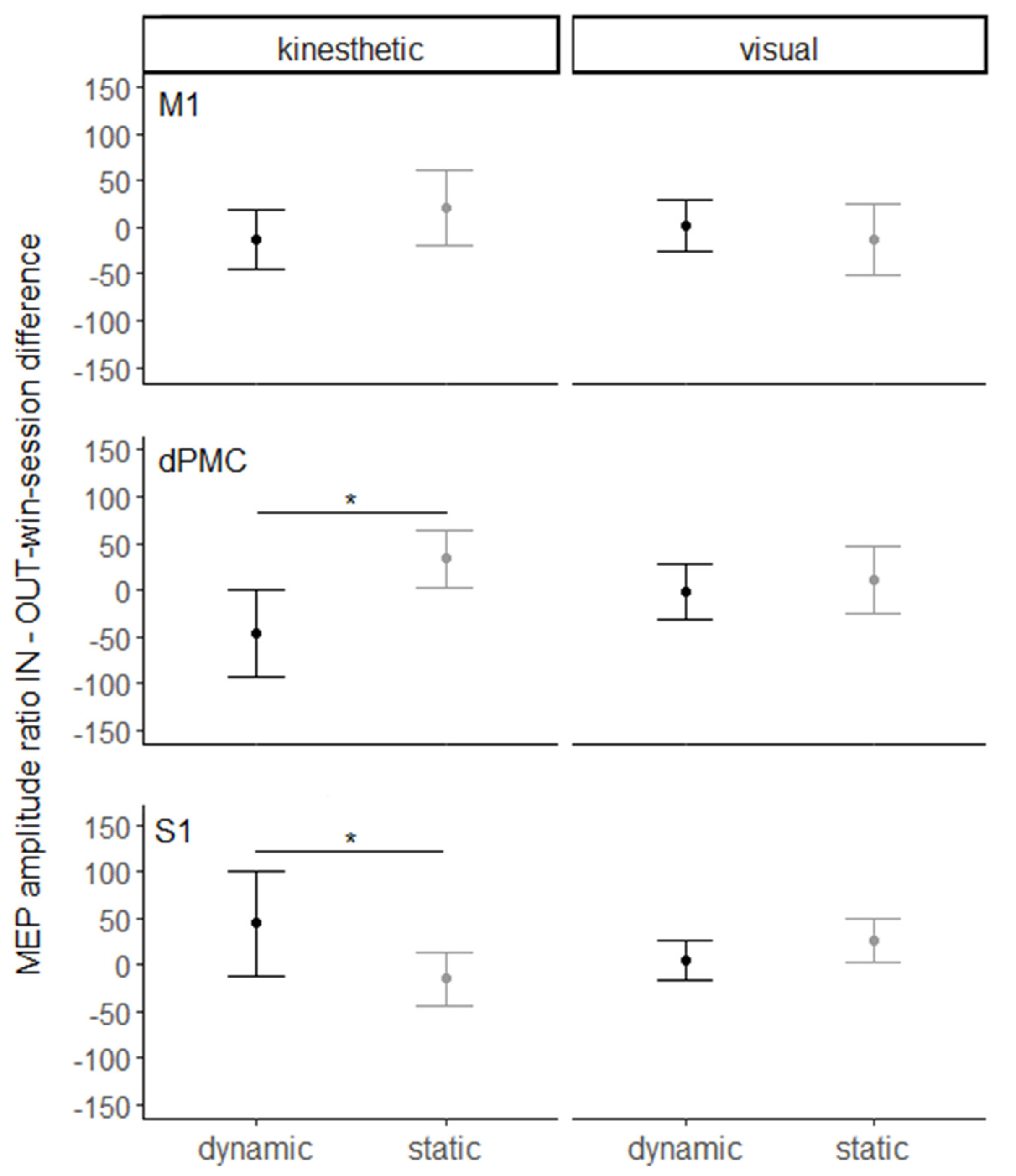
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeannerod, M. The representing brain: Neural correlates of motor intention and imagery. Behav. Brain Sci. 1994, 17, 187–202. [Google Scholar] [CrossRef]
- Wexler, M.; Kosslyn, S.M.; Berthoz, A. Motor processes in mental rotation. Cognition 1998, 68, 77–94. [Google Scholar] [CrossRef]
- Jeannerod, M.; Decety, J. Mental motor imagery: A window into the representational stages of action. Curr. Opin. Neurobiol. 1995, 5, 727–732. [Google Scholar] [CrossRef]
- Grèzes, J.; Decety, J. Does visual perception of object afford action? Evidence from a neuroimaging study. Neuropsychologia 2002, 40, 212–222. [Google Scholar] [CrossRef]
- Lotze, M.; Montoya, P.; Erb, M.; Hülsmann, E.; Flor, H.; Klose, U.; Birbaumer, N.; Grodd, W. Activation of cortical and cerebellar motor areas during executed and imagined hand movements: An fMRI study. J. Cogn. Neurosci. 1999, 11, 491–501. [Google Scholar] [CrossRef]
- Tanji, J.; Shima, K. Supplementary motor cortex in organization of movement. Eur. Neurol. 1996, 36, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.B.; Hughes, L.; Nimmo-Smith, I. Action selection: A race model for selected and non-selected actions distinguishes the contribution of premotor and prefrontal areas. Neuroimage 2010, 51, 888–896. [Google Scholar] [CrossRef]
- Porro, C.A.; Francescato, M.P.; Cettolo, V.; Diamond, M.E.; Baraldi, P.; Zuiani, C.; Bazzocchi, M.; Di Prampero, P.E. Primary motor and sensory cortex activation during motor performance and motor imagery: A functional magnetic resonance imaging study. J. Neurosci. 1996, 16, 7688–7698. [Google Scholar] [CrossRef]
- Yoo, S.S.; Freeman, D.K.; McCarthyIII, J.J.; Jolesz, F.A. Neural substrates of tactile imagery: A functional MRI study. Neuroreport 2003, 14, 581–585. [Google Scholar] [CrossRef]
- Kilteni, K.; Andersson, B.J.; Houborg, C.; Ehrsson, H.H. Motor imagery involves predicting the sensory consequences of the imagined movement. Nat. Commun. 2018, 9, 1617. [Google Scholar] [CrossRef]
- Gerardin, E.; Sirigu, A.; Léhericy, S.; Poline, J.B.; Gaymard, B.; Marsault, C.; Agid, Y.; Le Bihan, D. Partially overlapping neural networks for real and imagined hand movements. Cereb. Cortex 2000, 10, 1093–1104. [Google Scholar] [CrossRef]
- Tian, X.; Poeppel, D. Mental imagery of speech and movement implicates the dynamics of internal forward models. Front. Psychol. 2010, 1, 166. [Google Scholar] [CrossRef]
- Ridderinkhof, K.R.; Brass, M. How kinesthetic motor imagery works: A predictive-processing theory of visualization in sports and motor expertise. J. Physiol. Paris 2015, 109, 53–63. [Google Scholar] [CrossRef]
- Lotze, M.; Halsband, U. Motor imagery. J. Physiol. Paris 2006, 99, 386–395. [Google Scholar] [CrossRef]
- Saruco, E.; Guillot, A.; Saimpont, A.; Di Rienzo, F.; Durand, A.; Mercier, C.; Malouin, F.; Jackson, P. Motor imagery ability of patients with lower-limb amputation: Exploring the course of rehabilitation effects. Eur. J. Phys. Rehabil. Med. 2019, 55, 634–645. [Google Scholar] [CrossRef]
- Cho, H.Y.; Kim, J.S.; Lee, G.C. Effects of motor imagery training on balance and gait abilities in post-stroke patients: A randomized controlled trial. Clin. Rehabil. 2012, 27, 675–680. [Google Scholar] [CrossRef]
- Callow, N.; Roberts, R.; Hardy, L.; Jiang, D.; Edwards, M.G. Performance improvements from imagery: Evidence that internal visual imagery is superior to external visual imagery for slalom performance. Front. Hum. Neurosci. 2013, 7, 697. [Google Scholar] [CrossRef]
- Battaglia, C.; D’Artibale, E.; Fiorilli, G.; Piazza, M.; Tsopani, D.; Giombini, A.; Calcagno, G.; Di Cagno, A. Use of video observation and motor imagery on jumping performance in national rhythmic gymnastics athletes. Hum. Mov. Sci. 2014, 38, 225–234. [Google Scholar] [CrossRef]
- Theiler, A.M.; Lippman, L.G. Effects of mental practice and modeling on guitar and vocal performance. J. Gen. Psychol. 1995, 122, 329–343. [Google Scholar] [CrossRef]
- Madigan, R.; Frey, R.D.; Matlock, T.S. Cognitive strategies of university athletes. Can. J. Sport Sci. 1992, 17, 135–140. [Google Scholar]
- Féry, Y.A. Differentiating visual and kinesthetic imagery in mental practice. Can. J. Exp. Psychol. 2003, 57, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fourkas, A.D.; Ionta, S.; Aglioti, S.M. Influence of imagined posture and imagery modality on corticospinal excitability. Behav. Brain Res. 2006, 168, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Binkofski, F.; Amunts, K.; Stephan, K.M.; Posse, S.; Schormann, T.; Freund, H.J.; Zilles, K.; Seitz, R.J. Broca’s region subserves imagery of motion: A combined cytoarchitectonic and fMRI study. Hum. Brain Mapp. 2000, 11, 273–285. [Google Scholar] [CrossRef]
- Solodkin, A.; Hlustik, P.; Chen, E.E.; Small, S.L. Fine modulation in network activation during motor execution and motor imagery. Cereb. Cortex 2004, 14, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Collet, C.; Nguyen, V.A.; Malouin, F.; Richards, C.; Doyon, J. Brain activity during visual versus kinesthetic imagery: An fMRI study. Hum. Brain Mapp. 2009, 30, 2157–2172. [Google Scholar] [CrossRef]
- Jiang, D.; Edwards, M.G.; Mullins, P.; Callow, N. The neural substrates for the different modalities of movement imagery. Brain Cogn. 2015, 97, 22–31. [Google Scholar] [CrossRef]
- Seiler, B.D.; Monsma, E.V.; Newman-Norlund, R.D. Biological evidence of imagery abilities: Intraindividual differences. J. Sport Exerc. Psychol. 2015, 37, 421–435. [Google Scholar] [CrossRef]
- Hétu, S.; Grégoire, M.; Saimpont, A.; Coll, M.P.; Eugène, F.; Michon, P.E.; Jackson, P.L. The neural network of motor imagery: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 930–949. [Google Scholar] [CrossRef]
- Lee, W.H.; Kim, E.; Seo, H.G.; Oh, B.; Nam, H.S.; Kim, Y.J.; Lee, H.H.; Kang, M.; Kim, S.; Bang, M.S. Target-oriented motor imagery for grasping action: Different characteristics of brain activation between kinesthetic and visual imagery. Sci. Rep. 2019, 9, 12770. [Google Scholar] [CrossRef]
- Yang, Y.J.; Jeon, E.J.; Kim, J.S.; Chung, C.K. Characterization of kinesthetic motor imagery compared with visual motor imageries. Sci. Rep. 2021, 11, 3751. [Google Scholar] [CrossRef]
- Kasess, C.H.; Windischberger, C.; Cunnington, R.; Lanzenberger, R.; Pezawas, L.; Moser, E. The suppressive influence of SMA on M1 in motor imagery revealed by fMRI and dynamic causal modeling. Neuroimage 2008, 40, 828–837. [Google Scholar] [CrossRef]
- Waldvogel, D.; Van Gelderen, P.; Muellbacher, W.; Ziemann, U.; Immisch, I.; Hallett, M. The relative metabolic demand of inhibition and excitation. Nature 2000, 406, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Fadiga, L.; Buccino, G.; Craighero, L.; Fogassi, L.; Gallese, V.; Pavesi, G. Corticospinal excitability is specifically modulated by motor imagery: A magnetic stimulation study. Neuropsychologia 1999, 37, 147–158. [Google Scholar] [CrossRef]
- Bruno, V.; Fossataro, C.; Garbarini, F. Inhibition or facilitation? Modulation of corticospinal excitability during motor imagery. Neuropsychologia 2018, 111, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Facchini, S.; Muellbacher, W.; Battaglia, F.; Boroojerdi, B.; Hallett, M. Focal enhancement of motor cortex excitability during motor imagery: A transcranial magnetic stimulation study. Acta Neurol. Scand. 2002, 105, 146–151. [Google Scholar] [CrossRef]
- Fourkas, A.; Bonavolontà, V.; Avenanti, A.; Aglioti, S. Kinesthetic imagery and tool-specific modulation of corticospinal representations in expert tennis players. Cereb. Cortex 2008, 18, 2382–2390. [Google Scholar] [CrossRef] [PubMed]
- Stinear, C.M.; Byblow, W.D.; Steyvers, M.; Levin, O.; Swinnen, S.P. Kinesthetic, but not visual, motor imagery modulates corticomotor excitability. Exp. Brain Res. 2006, 168, 157–164. [Google Scholar] [CrossRef]
- Roosink, M.; Zijdewind, I. Corticospinal excitability during observation and imagery of simple and complex hand tasks: Implications for motor rehabilitation. Behav. Brain Res. 2010, 213, 35–41. [Google Scholar] [CrossRef]
- Guillot, A.; Collet, C.; Dittmar, A. Relationship between visual and kinesthetic imagery, field dependence-independence, and complex motors skills. J. Psychophysiol. 2004, 18, 190–198. [Google Scholar] [CrossRef]
- Filgueiras, A.; Quintas Conde, E.F.; Hall, C.R. The neural basis of kinesthetic and visual imagery in sports: An ALE meta—Analysis. Brain Imaging Behav. 2018, 12, 1513–1523. [Google Scholar] [CrossRef]
- Avenanti, A.; Bolognini, N.; Maravita, A.; Aglioti, S.M. Somatic and Motor Components of Action Simulation. Curr. Biol. 2007, 17, 2129–2135. [Google Scholar] [CrossRef]
- Avenanti, A.; Annella, L.; Candidi, M.; Urgesi, C.; Aglioti, S. Compensatory plasticity in the action observation network: Virtual lesions of STS enhance anticipatory simulation of seen actions. Cereb. Cortex 2013, 23, 570–580. [Google Scholar] [CrossRef]
- Chen, R.; Classen, J.; Gerloff, C.; Celnik, P.; Wassermann, E.M.; Hallett, M.; Cohen, L.G. Depression of motor cortex excitability by low-frequency transcranial magnetic stimulation. Neurology 1997, 48, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Gerschlager, W.; Siebner, H.R.; Rothwell, J.C. Decreased corticospinal excitability after subthreshold 1 Hz rTMS over lateral premotor cortex. Neurology 2001, 57, 449–455. [Google Scholar] [CrossRef]
- O’Shea, J.; Johansen-Berg, H.; Trief, D.; Göbel, S.; Rushworth, M.F.S. Functionally Specific Reorganization in Human Premotor Cortex. Neuron 2007, 54, 479–490. [Google Scholar] [CrossRef]
- Beck, M.M.; Grandjean, M.U.; Hartmand, S.; Spedden, M.E.; Christiansen, L.; Roig, M.; Lundbye-Jensen, J. Acute Exercise Protects Newly Formed Motor Memories Against rTMS-induced Interference Targeting Primary Motor Cortex. Neuroscience 2020, 436, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Knecht, S.; Ellger, T.; Breitenstein, C.; Ringelstein, E.B.; Henningsen, H. Changing cortical excitability with low-frequency transcranial magnetic stimulation can induce sustained disruption of tactile perception. Biol. Psychiatry 2003, 53, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Satow, T.; Mima, T.; Yamamoto, J.; Oga, T.; Begum, T.; Aso, T.; Hashimoto, N.; Rothwell, J.C.; Shibasaki, H. Short-lasting impairment of tactile perception by 0.9Hz-rTMS of the sensorimotor cortex. Neurology 2003, 60, 1045–1047. [Google Scholar] [CrossRef] [PubMed]
- Balslev, D.; Christensen, L.O.D.; Lee, J.-H.; Law, I.; Paulson, O.B.; Miall, R.C. Enhanced Accuracy in Novel Mirror Drawing after Repetitive Transcranial Magnetic Stimulation-Induced Proprioceptive Deafferentation. J. Neurosci. 2004, 24, 9698. [Google Scholar] [CrossRef] [PubMed]
- Münchau, A.; Bloem, B.R.; Irlbacher, K.; Trimble, M.R.; Rothwell, J.C. Functional connectivity of human premotor and motor cortex explored with repetitive transcranial magnetic stimulation. J. Neurosci. 2002, 22, 554–561. [Google Scholar] [CrossRef]
- Chouinard, P.A.; Van Der Werf, Y.D.; Leonard, G.; Paus, T. Modulating neural networks with transcranial magnetic stimulation applied over the dorsal premotor and primary motor cortices. J. Neurophysiol. 2003, 90, 1071–1083. [Google Scholar] [CrossRef]
- Jacobs, M.; Tsang, P.; Lee, K.; Asmussen, M.; Zapallow, C.; Nelson, A. 30 Hz theta-burst stimulation over primary somatosensory cortex modulates corticospinal output to the hand. Brain Stimul. 2014, 7, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Sauner, D.; Bestmann, S.; Siebner, H.; Rothwell, J. No evidence for a substantial involvement of primary motor hand area in handedness judgements: A transcranial magnetic stimulation study. Eur. J. Neurosci. 2006, 23, 2215–2224. [Google Scholar] [CrossRef]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef] [PubMed]
- Fourkas, A.D.; Avenanti, A.; Urgesi, C.; Aglioti, S.M. Corticospinal facilitation during first and third person imagery. Exp. Brain Res. 2006, 168, 143–151. [Google Scholar] [CrossRef]
- Bufalari, I.; Sforza, A.; Cesari, P.; Aglioti, S.M.; Fourkas, A.D. Motor imagery beyond the joint limits: A transcranial magnetic stimulation study. Biol. Psychol. 2010, 85, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Yahagi, S.; Kasai, T. Motor evoked potentials induced by motor imagery reveal a functional asymmetry of cortical motor control in left- and right-handed human subjects. Neurosci. Lett. 1999, 276, 185–188. [Google Scholar] [CrossRef]
- Willems, R.M.; Toni, I.; Hagoort, P.; Casasanto, D. Body-specific motor imagery of hand actions: Neural evidence from right- and left-handers. Front. Hum. Neurosci. 2009, 3, 39. [Google Scholar] [CrossRef] [PubMed]
- Sugishita, M.; Takayama, Y. Paraesthesia elicited by repetitive magnetic stimulation of the postcentral gyrus. Neuroreport 1993, 4, 569–570. [Google Scholar] [CrossRef]
- Tegenthoff, M.; Ragert, P.; Pleger, B.; Schwenkreis, P.; Förster, A.F.; Nicolas, V.; Dinse, H.R. Improvement of tactile discrimination performance and enlargement of cortical somatosensory maps after 5 Hz rTMS. PLoS Biol. 2005, 3, e362. [Google Scholar] [CrossRef]
- Hall, C.R.; Martin, K.A. Measuring movement imagery abilities A revision of the movement imagery questionnaire. J. Ment. Imag. 1997, 21, 143–154. [Google Scholar]
- Catmur, C.; Mars, R.B.; Rushworth, M.F.; Heyes, C. Making mirrors: Premotor cortex stimulation enhances mirror and counter-mirror motor facilitation. J. Cogn. Neurosci. 2011, 23, 2352–2362. [Google Scholar] [CrossRef]
- Cavallo, A.; Bucchioni, G.; Castiello, U.; Becchio, C. Goal or movement? Action representation within the primary motor cortex. Eur. J. Neurosci. 2013, 38, 3507–3512. [Google Scholar] [CrossRef]
- Urgesi, C.; Candidi, M.; Fabbro, F.; Romani, M.; Aglioti, S.M. Motor facilitation during action observation: Topographic mapping of the target muscle and influence of the onlooker’s posture. Eur. J. Neurosci. 2006, 23, 2522–2530. [Google Scholar] [CrossRef]
- Meteyard, L.; Davies, R. Best practice guidance for linear mixed-effects models in psychological science. J. Mem. Lang. 2019, 112, 104092. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Avenanti, A.; Candidi, M.; Urgesi, C. Vicarious motor activation during action perception: Beyond correlational evidence. Front. Hum. Neurosci. 2013, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, R.M.; Caspers, S.; Eickhoff, S.B.; Swinnen, S.P. Neural correlates of action: Comparing meta-analyses of imagery, observation, and execution. Neurosci. Biobehav. Rev. 2018, 94, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kuhtz-Buschbeck, J.P.; Mahnkopf, C.; Holzknecht, C.; Siebner, H.; Ulmer, S.; Jansen, O. Effector-independent representations of simple and complex imagined finger movements: A combined fMRI and TMS study. Eur. J. Neurosci. 2003, 18, 3375–3387. [Google Scholar] [CrossRef] [PubMed]
- Szameitat, A.J.; Shen, S.; Sterr, A. Effector-dependent activity in the left dorsal premotor cortex in motor imagery. Eur. J. Neurosci. 2007, 26, 3303–3308. [Google Scholar] [CrossRef]
- Guillot, A.; Di Rienzo, F.; Macintyre, T.; Moran, A.; Collet, C. Imagining is not doing but involves specific motor commands: A review of experimental data related to motor inhibition. Front. Hum. Neurosci. 2012, 6, 247. [Google Scholar] [CrossRef]
- Sirigu, A.; Duhamel, J.R.; Cohen, L.; Pillon, B.; Dubois, B.; Agid, Y. The mental representation of hand movements after parietal cortex damage. Science 1996, 273, 1564–1566. [Google Scholar] [CrossRef] [PubMed]
- Schwoebel, J.; Boronat, C.B.; Branch Coslett, H. The man who executed “imagined” movements: Evidence for dissociable components of the body schema. Brain Cogn. 2002, 50, 1–16. [Google Scholar] [CrossRef]
- Blakemore, S.J.; Sirigu, A. Action prediction in the cerebellum and in the parietal lobe. Exp. Brain Res. 2003, 153, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.H.; Sprehn, G.; Saykin, A.J. Intact motor imagery in chronic upper limb hemiplegics: Evidence for activity-independent action representations. J. Cogn. Neurosci. 2002, 14, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Case, L.K.; Pineda, J.; Ramachandran, V.S. Common coding and dynamic interactions between observed, imagined, and experienced motor and somatosensory activity. Neuropsychologia 2015, 79, 233–245. [Google Scholar] [CrossRef]
- Silva, S.; Loubinoux, I.; Olivier, M.; Bataille, B.; Fourcade, O.; Samii, K.; Jeannerod, M.; Démonet, J.F. Impaired visual hand recognition in preoperative patients during brachial plexus anesthesia: Importance of peripheral neural input for mental representation of the hand. Anesthesiology 2011, 114, 126–134. [Google Scholar] [CrossRef]
- Hashimoto, R.; Rothwell, J.C. Dynamic changes in corticospinal excitability during motor imagery. Exp. Brain Res. 1999, 125, 75–81. [Google Scholar] [CrossRef]
- Hyde, C.; Fuelscher, I.; Lum, J.A.G.; Williams, J.; He, J.; Enticott, P.G. Primary Motor Cortex Excitability Is Modulated during the Mental Simulation of Hand Movement. J. Int. Neuropsychol. Soc. 2017, 23, 185–193. [Google Scholar] [CrossRef][Green Version]
- Fadiga, L.; Craighero, L.; Olivier, E. Human motor cortex excitability during the perception of others’ action. Curr. Opin. Neurobiol. 2005, 15, 213–218. [Google Scholar] [CrossRef]
- Goldsworthy, M.R.; Hordacre, B.; Rothwell, J.C.; Ridding, M.C. Effects of rTMS on the brain: Is there value in variability? Cortex 2021, 139, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.R.; Anschel, D.; Sparing, R.; Gangitano, M.; Pascual-Leone, A. Subthreshold low frequency repetitive transcranial magnetic stimulation selectively decreases facilitation in the motor cortex. Clin. Neurophysiol. 2002, 113, 101–107. [Google Scholar] [CrossRef]
- Bäumer, T.; Lange, R.; Liepert, J.; Weiller, C.; Siebner, H.R.; Rothwell, J.C.; Münchau, A. Repeated premotor rTMS leads to cumulative plastic changes of motor cortex excitability in humans. Neuroimage 2003, 20, 550–560. [Google Scholar] [CrossRef]
- Kim, J.S.; Oh, D.W.; Kim, S.Y.; Choi, J.D. Visual and kinesthetic locomotor imagery training integrated with auditory step rhythm for walking performance of patients with chronic stroke. Clin. Rehabil. 2011, 25, 134–145. [Google Scholar] [CrossRef]
- Lim, V.K.; Polych, M.A.; Holländer, A.; Byblow, W.D.; Kirk, I.J.; Hamm, J.P. Kinesthetic but not visual imagery assists in normalizing the CNV in Parkinson’s disease. Clin. Neurophysiol. 2006, 117, 2308–2314. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Out-Win Session | ||||
| Kinesthetic MI | Visual MI | |||
| Dynamic | Static | Dynamic | Static | |
| M1 | 130.7 (14.0) | 125.2 (14.2) | 129.3 (14.2) | 151.7 (14.5) |
| dPMC | 158.4 (14.0) | 98.3 (14.0) | 132.1 (14.0) | 137.3 (14.0) |
| S1 | 125.8 (14.1) | 115.9 (14.0) | 98.3 (14.1) | 96.1 (14.0) |
| In-Win Session | ||||
| Kinesthetic MI | Visual MI | |||
| Dynamic | Static | Dynamic | Static | |
| M1 | 120.3 (14.2) | 145.5 (14.1) | 130.9 (14.0) | 129.7 (14.1) |
| dPMC | 114.0 (14.1) | 132.0 (14.1) | 130.5 (14.1) | 148.0 (14.0) |
| S1 | 170.0 (14.0) | 100.7 (14.1) | 103.5 (14.1) | 123.5 (14.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oldrati, V.; Finisguerra, A.; Avenanti, A.; Aglioti, S.M.; Urgesi, C. Differential Influence of the Dorsal Premotor and Primary Somatosensory Cortex on Corticospinal Excitability during Kinesthetic and Visual Motor Imagery: A Low-Frequency Repetitive Transcranial Magnetic Stimulation Study. Brain Sci. 2021, 11, 1196. https://doi.org/10.3390/brainsci11091196
Oldrati V, Finisguerra A, Avenanti A, Aglioti SM, Urgesi C. Differential Influence of the Dorsal Premotor and Primary Somatosensory Cortex on Corticospinal Excitability during Kinesthetic and Visual Motor Imagery: A Low-Frequency Repetitive Transcranial Magnetic Stimulation Study. Brain Sciences. 2021; 11(9):1196. https://doi.org/10.3390/brainsci11091196
Chicago/Turabian StyleOldrati, Viola, Alessandra Finisguerra, Alessio Avenanti, Salvatore Maria Aglioti, and Cosimo Urgesi. 2021. "Differential Influence of the Dorsal Premotor and Primary Somatosensory Cortex on Corticospinal Excitability during Kinesthetic and Visual Motor Imagery: A Low-Frequency Repetitive Transcranial Magnetic Stimulation Study" Brain Sciences 11, no. 9: 1196. https://doi.org/10.3390/brainsci11091196
APA StyleOldrati, V., Finisguerra, A., Avenanti, A., Aglioti, S. M., & Urgesi, C. (2021). Differential Influence of the Dorsal Premotor and Primary Somatosensory Cortex on Corticospinal Excitability during Kinesthetic and Visual Motor Imagery: A Low-Frequency Repetitive Transcranial Magnetic Stimulation Study. Brain Sciences, 11(9), 1196. https://doi.org/10.3390/brainsci11091196