Sublethal Injury Caused to Listeria monocytogenes by Natural Plant Extracts: Case Study on Grape Seed Extract and Garlic Extract


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Preculture Conditions
2.2. Preparation of Bioactive Extracts
2.3. Minimum Inhibitory Concentration (MIC)
2.4. Challenge Testing
2.5. Assessment of Sublethal Injury
2.6. Determination of Propidium Iodide Uptake
2.7. Statistical Analysis
3. Results and Discussion
3.1. Minimum Inhibitory Concentration (MIC)
3.2. Challenge Testing
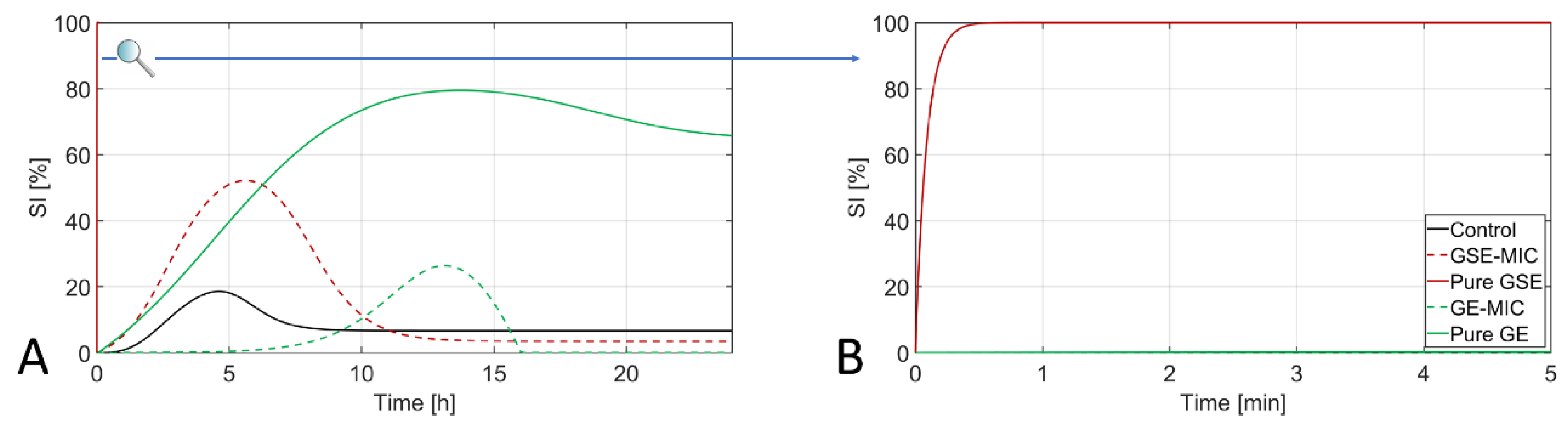
3.3. Assessment of Sublethal Injury
3.4. Determination of Propidium Iodide Uptake
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brashears, M.M.; Amezquita, A.; Stratton, J. Validation of methods used to recover Escherichia coli O157: H7 and Salmonella spp. subjected to stress conditions. J. Food Prot. 2001, 64, 1466–1471. [Google Scholar] [CrossRef] [PubMed]
- Noriega, E.; Velliou, E.G.; Van Derlinden, E.; Mertens, L.; Van Impe, J. Effect of cell immobilization on heat-induced sublethal injury of Escherichia coli, Salmonella Typhimurium and Listeria innocua. Food Microbiol. 2013, 36, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Hurst, A. Bacterial injury: A review. Can. J. Microbiol. 1977, 23, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Devlieghere, F.; Vermeiren, L.; Debevere, J. New preservation technologies: Possibilities and limitations. Int. Dairy J. 2004, 14, 273–285. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The use of plant antimicrobial compounds for food preservation. BioMed. Res. Int. 2015, 2015, 246264. [Google Scholar] [CrossRef] [PubMed]
- Negi, P.S. Plant extracts for the control of bacterial growth: Efficacy, stability and safety issues for food application. Int. J. Food Microbiol. 2012, 156, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Belguith, H.; Kthiri, F.; Chati, A.; Sofah, A.A.; Hamida, J.B.; Landoulsi, A. Study of the effect of aqueous garlic extract (Allium sativum) on some Salmonella serovars isolates. Emir. J. Food Agric. 2010, 22, 189–206. [Google Scholar] [CrossRef]
- Silván, J.M.; Mingo, E.; Hidalgo, M.; de Pascual-Teresa, S.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Antibacterial activity of a grape seed extract and its fractions against Campylobacter spp. Food Control. 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Perumalla, A.V.S.; Hettiarachchy, N.S. Green tea and grape seed extracts—Potential applications in food safety and quality. Food Res. Int. 2011, 44, 827–839. [Google Scholar] [CrossRef]
- Miron, T.; Rabinkov, A.; Mirelman, D.; Wilchek, M.; Weiner, I. The mode of action of allicin: Its ready permeability through phospholipid membranes may contribute to its biological activity. Biochim. Biophys. Acta Biomembr. 2000, 1463, 20–30. [Google Scholar] [CrossRef]
- Baka, M.; Noriega, E.; Tsakali, E.; Van Impe, J.F. Influence of composition and processing of Frankfurter sausages on the growth dynamics of Listeria monocytogenes under vacuum. Food Res. Int. 2015, 70, 94–100. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar] [CrossRef]
- Jones, D. Foodborne listeriosis. Lancet 1990, 336, 1171–1174. [Google Scholar] [CrossRef]
- Gadang, V.P.; Hettiarachchy, N.S.; Johnson, M.G.; Owens, C. Evaluation of antibacterial activity of whey protein isolate coating incorporated with nisin, grape seed extract, malic acid, and EDTA on a turkey frankfurter system. J. Food Sci. 2008, 73, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Kakaei, S.; Shahbazi, Y. Effect of chitosan-gelatin film incorporated with ethanolic red grape seed extract and Ziziphora clinopodioides essential oil on survival of Listeria monocytogenes and chemical, microbial and sensory properties of minced trout fillet. Food Sci. Technol. 2016, 72, 432–438. [Google Scholar] [CrossRef]
- Moradi, M.; Tajik, H.; Rohani, S.M.R.; Oromiehie, A.R. Effectiveness of Zataria multiflora Boiss essential oil and grape seed extract impregnated chitosan film on ready-to-eat mortadella-type sausages during refrigerated storage. J. Sci. Food Agric. 2011, 91, 2850–2857. [Google Scholar] [CrossRef] [PubMed]
- Theivendran, S.; Hettiarachchy, N.S.; Johnson, M.G. Inhibition of Listeria monocytogenes by nisin combined with grape seed extract or green tea extract in soy protein film coated on turkey frankfurters. J. Food Sci. 2006, 71, 39–44. [Google Scholar] [CrossRef]
- Perumalla, A.V.S.; Hettiarachchy, N.S.; Over, K.; Ricke, S.C.; Slavik, M.F.; Gbur, E.; Davis, B.; Acosta, S. Effect of partial replacement of potassium lactate and sodium diacetate by natural green tea and grape seed extracts and postpackaging thermal treatment on the growth of Listeria monocytogenes in hotdog model system. Int. J. Food Sci. Technol. 2013, 48, 918–926. [Google Scholar] [CrossRef]
- Tajik, H.; Aminzare, M.; Raad, T.M.; Hashemi, M.; Azar, H.H.; Raeisi, M.; Naghili, H. Effect of Zataria Multiflora Boiss essential oil and grape seed extract on the shelf life of raw buffalo patty and fate of inoculated Listeria monocytogenes. J. Food Process Preserv. 2015, 39, 3005–3013. [Google Scholar] [CrossRef]
- Pinilla, C.M.B.; Noreña, C.P.Z.; Brandelli, A. Antimicrobial activity of nanoliposomes co-encapsulating nisin and garlic extract against Gram-positive and Gram-negative bacteria in milk. Innov. Food Sci. Emerg. Technol. 2016, 36, 287–293. [Google Scholar] [CrossRef]
- Pinilla, C.M.B.; Brandelli, A. Development and characterization of phosphatidylcholine nanovesicles, containing garlic extract, with antilisterial activity in milk. Food Chem. 2017, 220, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Moreira, R.G.; Castell-Perez, E. Microencapsulated antimicrobial compounds as a means to enhance electron beam irradiation treatment for inactivation of pathogens on fresh spinach leaves. J. Food Sci. 2011, 76, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Laxdal, V.A.; Yu, M.; Raney, B.L. Antioxidant activity of allicin, an active principle in garlic. Mol. Cell. Biochem. 1995, 148, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Geeraerd, A.H.; Herremans, C.H.; Van Impe, J.F. Structural model requirements to describe microbial inactivation during a mild heat treatment. Int. J. Food Microbiol. 2000, 59, 185–209. [Google Scholar] [CrossRef]
- Busch, S.V.; Donnelly, C.W. Development of a repair-enrichment broth for resuscitation of heat-injured Listeria monocytogenes and Listeria innocua. Appl. Environ. Microbiol. 1992, 58, 14–20. [Google Scholar]
- Miller, F.A.; Brandão, T.R.S.; Teixeira, P.; Silva, C.L.M. Recovery of heat-injured Listeria innocua. Int. J. Food Microbiol. 2006, 112, 261–265. [Google Scholar] [CrossRef]
- Coder, D.M. Current Protocols in Cytometry; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1997; pp. 9.2.1–9.2.14. [Google Scholar]
- Anastasiadi, M.; Chorianopoulos, N.G.; Nychas, G.J.E.; Haroutounian, S.A. Antilisterial activities of polyphenolic-rich extracts of grapes and vinification byproducts. J. Agric. Food Chem. 2009, 57, 457–463. [Google Scholar] [CrossRef]
- Baydar, N.G.; Sagdic, O.; Ozkan, G.; Cetin, S. Determination of antibacterial effects and total phenolic contents of grape (Vitis vinifera L.) seed extracts. Int. J. Food Sci. Technol. 2006, 41, 799–804. [Google Scholar] [CrossRef]
- Yigit, D.; Yigit, N.; Mavi, A.; Yildirim, A.; Güleryüz, M. Antioxidant and antimicrobial activities of methanol and water extracts of fruits, leaves and seeds of Vitis vinifera L. cv. Karaerik. Asian J. Chem. 2009, 21, 183–194. [Google Scholar]
- Bakri, I.M.; Douglas, C.W.I. Inhibitory effect of garlic extract on oral bacteria. Arch. Oral Biol. 2005, 50, 645–651. [Google Scholar] [CrossRef]
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999, 2, 125–129. [Google Scholar] [CrossRef]
- Rohani, S.M.R.; Moradi, M.; Mehdizadeh, T.; Saei-Dehkordi, S.S.; Griffiths, M.W. The effect of nisin and garlic (Allium sativum L.) essential oil separately and in combination on the growth of Listeria monocytogenes. Food Sci. Technol. 2011, 44, 2260–2265. [Google Scholar] [CrossRef]
- Perni, S.; Chalise, P.R.; Shama, G.; Kong, M.G. Bacterial cells exposed to nanosecond pulsed electric fields show lethal and sublethal effects. Int. J. Food Microbiol. 2007, 120, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Bisha, B.; Weinsetel, N.; Brehm-Stecher, B.F.; Mendonca, A. Antilisterial effects of gravinol-S grape seed extract at low levels in aqueous media and its potential application as a produce wash. J. Food Prot. 2010, 73, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Kapadnis, B.P. Heat stable antimicrobial activity of Allium ascalonicum against bacteria and fungi. Indian. J. Exp. Biol. 2005, 43, 751–754. [Google Scholar] [PubMed]
- Hsieh, Y.-H.; Yan, M.; Liu, J.-G.; Hwang, J.C. The synergistic effect of nisin and garlic shoot juice against Listeria spp. in soymilk. J. Taiwan Inst. Chem. Eng. 2011, 42, 576–579. [Google Scholar] [CrossRef]
- Kim, E.L.; Choi, N.H.; Bajpai, V.K.; Kang, S.C. Synergistic effect of nisin and garlic shoot juice against Listeria monocytogenes in milk. Food Chem. 2008, 110, 375–382. [Google Scholar] [CrossRef]
- Leuschner, R.G.K.; Ielsch, V. Antimicrobial effects of garlic, clove and red hot chilli on Listeria monocytogenes in broth model systems and soft cheese. Int. J. Food Sci. Nutr. 2003, 54, 127–133. [Google Scholar] [CrossRef]
- Singh, B.; Falahee, M.B.; Adams, M.R. Synergistic inhibition of Listeria monocytogenes by nisin and garlic extract. Food Microbiol. 2001, 18, 133–139. [Google Scholar] [CrossRef]
- Wu, V.C.H. A review of microbial injury and recovery methods in food. Food Microbiol. 2008, 25, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Pereira, A.M.; Pereira, M.C.; Melo, L.F.; Simões, M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. J. Antimicrob. Chemother. 2011, 66, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Lobete, M.M.; Noriega, E.; Batalha, M.A.; De Beurme, S.; Van de Voorde, I.; Van Impe, J.F. Effect of tagatose on growth dynamics of Salmonella Typhimurium and Listeria monocytogenes in media with different levels of structural complexity and in UHT skimmed milk. Food Control. 2017, 73, 31–42. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, F.; Xu, H.; Zoraida, P.A.; Niu, R.; Yuan, Y.; Sun, J.; You, X.; Lai, W.; Xiong, Y.; et al. Magnetic nano-beads based separation combined with propidiummonoazide treatment and multiplex PCR assay for simultaneous detection of viable Salmonella Typhimurium, Escherichia coli O157: H7 and Listeria monocytogenes in food products. Food Microbiol. 2013, 34, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Drazic, A.; Kutzner, E.; Winter, J.; Eisenreich, W. Metabolic response of Escherichia coli upon treatment with Hypchlorite at sub-lethal concentrations. PLoS ONE 2015, 10, e0125823. [Google Scholar] [CrossRef] [PubMed]
- Feldberg, R.; Chang, S. In vitro mechanism of inhibition of bacterial cell growth by allicin. Antimicrob. Agents Chemother. 1988, 32, 1763–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Growth Parameter | Control | GSE-MIC | GE-MIC | |||
|---|---|---|---|---|---|---|
| TSAYE | Oxford | TSAYE | Oxford | TSAYE | Oxford | |
| λ [h] | 1.7 ± 0.2 | 1.7 ± 0.2 | 2.5 ± 0.3 | 3.2 ± 0.2 | 12.3 ± 0.4 | 13.1 ± 0.5 |
| A,a | A,a | B,a | B,b | C,a | C,a | |
| µmax [1/h] | 1.46 ± 0.09 | 1.40 ± 0.08 | 1.04 ± 0.10 | 0.959 ± 0.065 | 0.725 ± 0.068 | 0.907 ± 0.122 |
| C,a | B,a | B,a | A,a | A,a | A,a | |
| Nmax [(CFU/mL)] | 21.6 ± 0.0 | 21.6 ± 0.0 | 21.2 ± 0.1 | 21.2 ± 0.1 | 21.3 ± 0.2 | 21.6 ± 0.3 |
| B,a | B,a | A,a | A,a | A,a | AB,a | |
| Inactivation Parameter | Pure GSE | Pure GE | ||
|---|---|---|---|---|
| TSAYE | Oxford | TSAYE | Oxford | |
| SL [h] | 0.0 ± 0.1 | 0 ± 0.0 | 11.4 ± 0.7 | 5.2 ± 1.7 |
| A,a | A,a | B,b | B,a | |
| kmax [1/h] | 11.4 ± 2.8 | 698 ± 247 | 0.420 ± 0.055 | 0.341 ± 0.061 |
| B,a | B,b | A,a | A,a | |
| Nres [log(CFU/mL)] | 2.7 ± 0.2 | 2.2 ± 0.3 | 5.7 ± 0.1 | 5.2 ± 0.2 |
| A,a | A,a | B,b | B,a | |
| Treatment Condition | TICC [–] |
|---|---|
| Control | 7.8 |
| GSE-MIC | 15.1 |
| GE-MIC | 5.4 |
| Pure GSE | 98.3 |
| Pure GE | 59.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verheyen, D.; Baka, M.; Van Impe, J.F.M. Sublethal Injury Caused to Listeria monocytogenes by Natural Plant Extracts: Case Study on Grape Seed Extract and Garlic Extract. Appl. Sci. 2019, 9, 2731. https://doi.org/10.3390/app9132731
Verheyen D, Baka M, Van Impe JFM. Sublethal Injury Caused to Listeria monocytogenes by Natural Plant Extracts: Case Study on Grape Seed Extract and Garlic Extract. Applied Sciences. 2019; 9(13):2731. https://doi.org/10.3390/app9132731
Chicago/Turabian StyleVerheyen, Davy, Maria Baka, and Jan F. M. Van Impe. 2019. "Sublethal Injury Caused to Listeria monocytogenes by Natural Plant Extracts: Case Study on Grape Seed Extract and Garlic Extract" Applied Sciences 9, no. 13: 2731. https://doi.org/10.3390/app9132731