Physical and Cognitive Performance of the Least Shrew (Cryptotis parva) on a Calcium-Restricted Diet

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animal Husbandry and Diet

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calcium (mg/g) | Treatment |
|---|---|
| 4.27 ± 0.68 | Base diet |
| 1.10 ± 0.34 | Low |
| 19.47 ± 5.05 | High |
2.2. Performance Assays

2.3. Data Analysis
3. Results
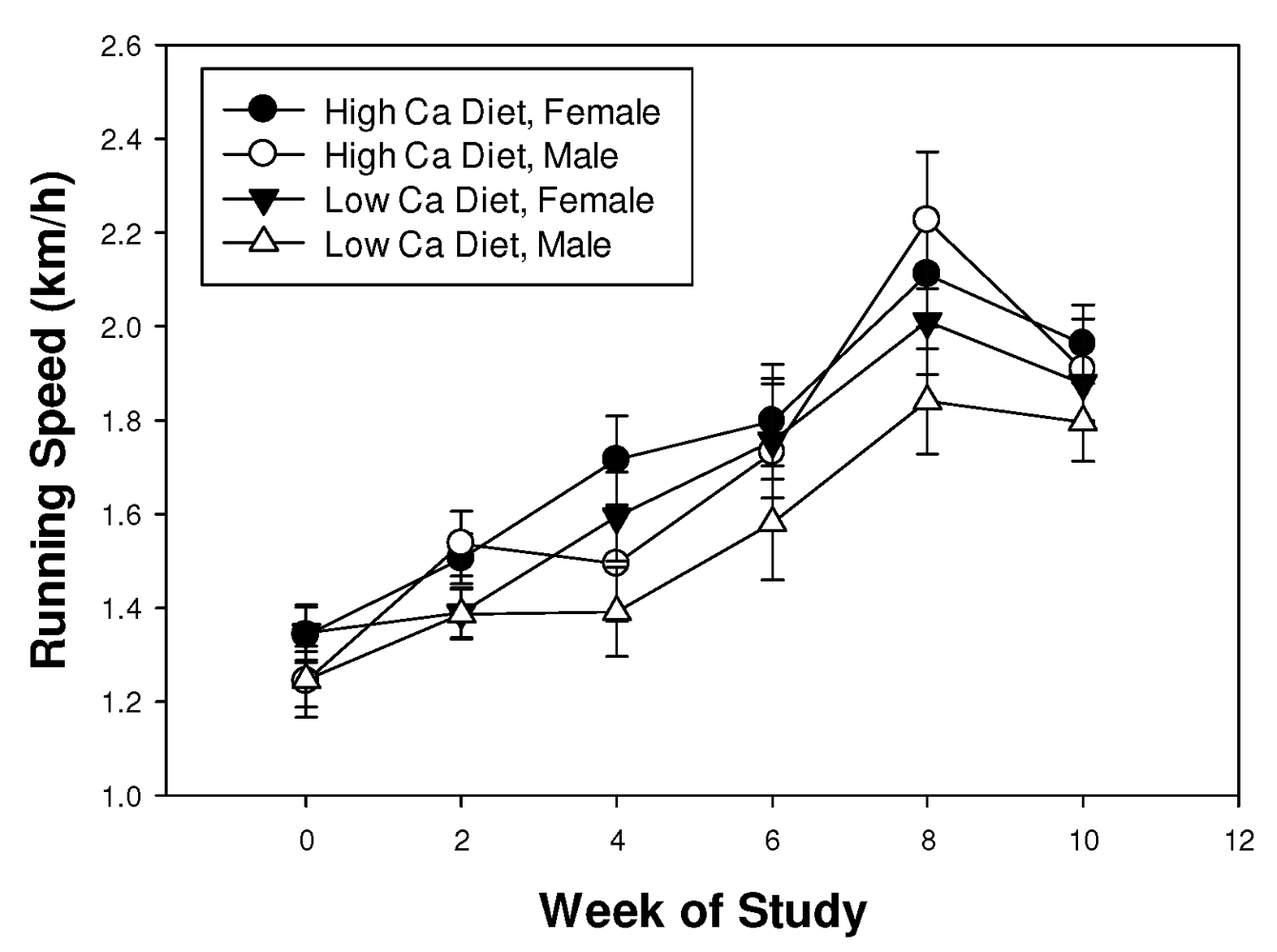
3.1. Running Track Trial
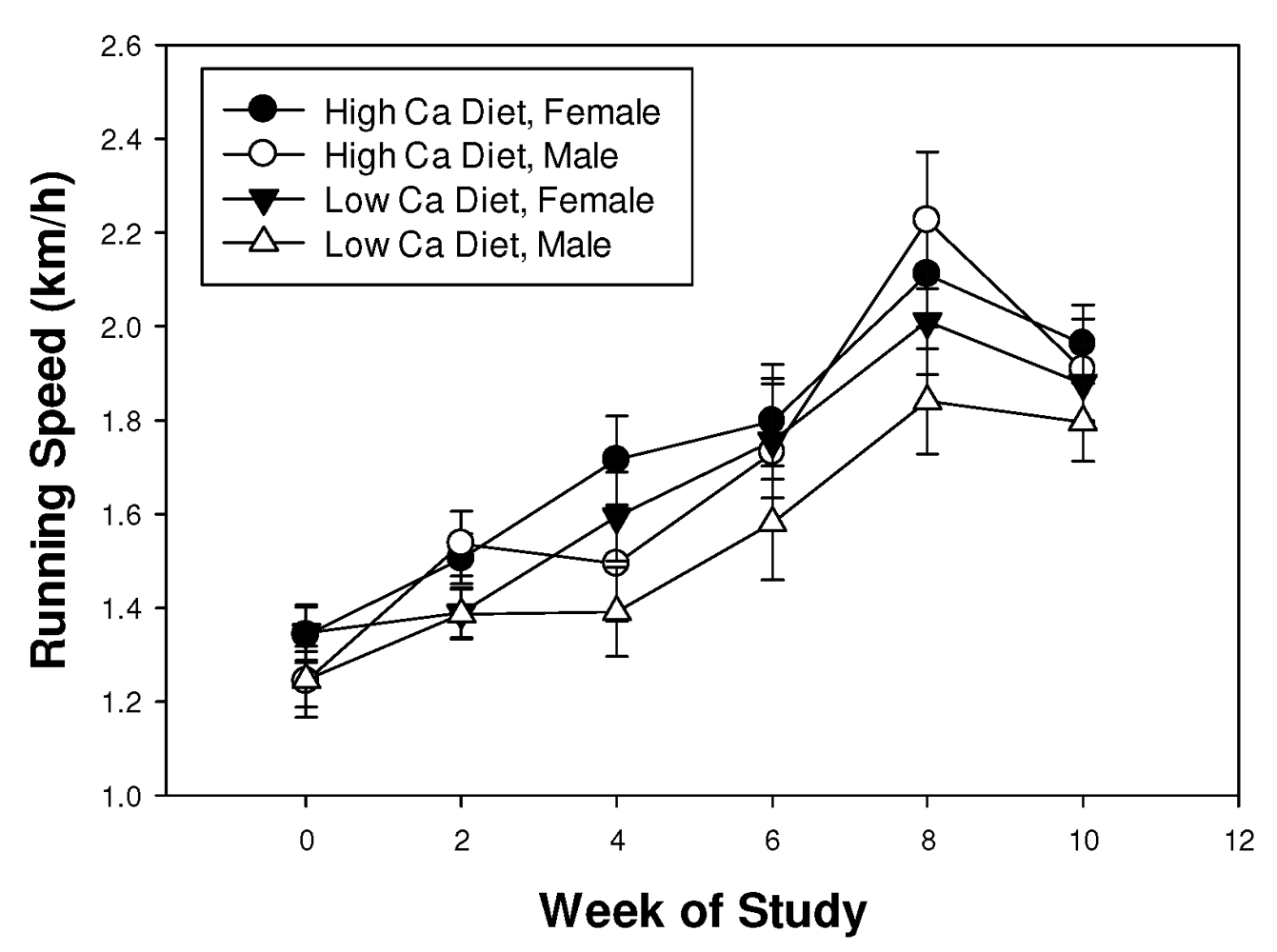
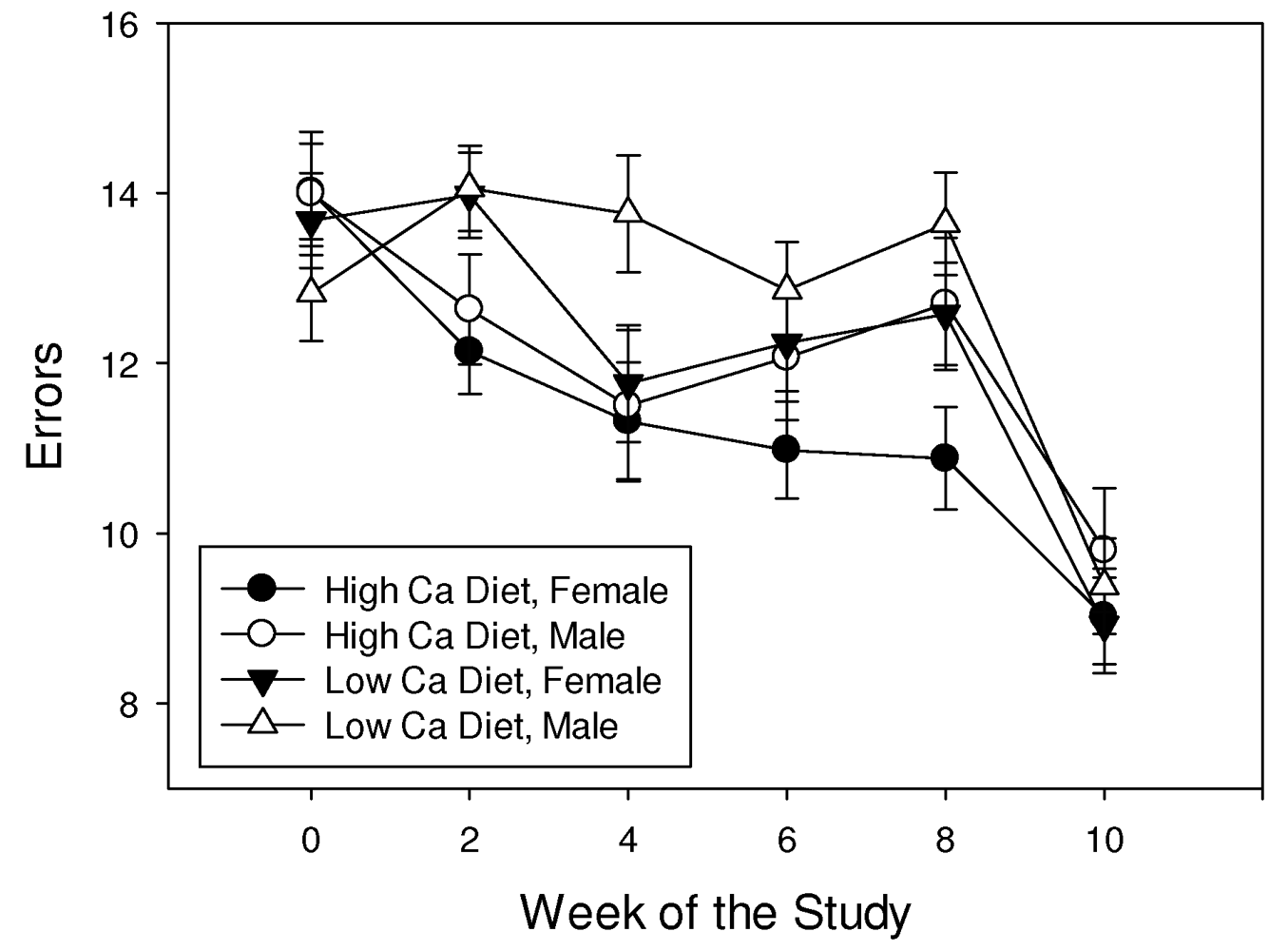
3.2. Complex Maze Trial
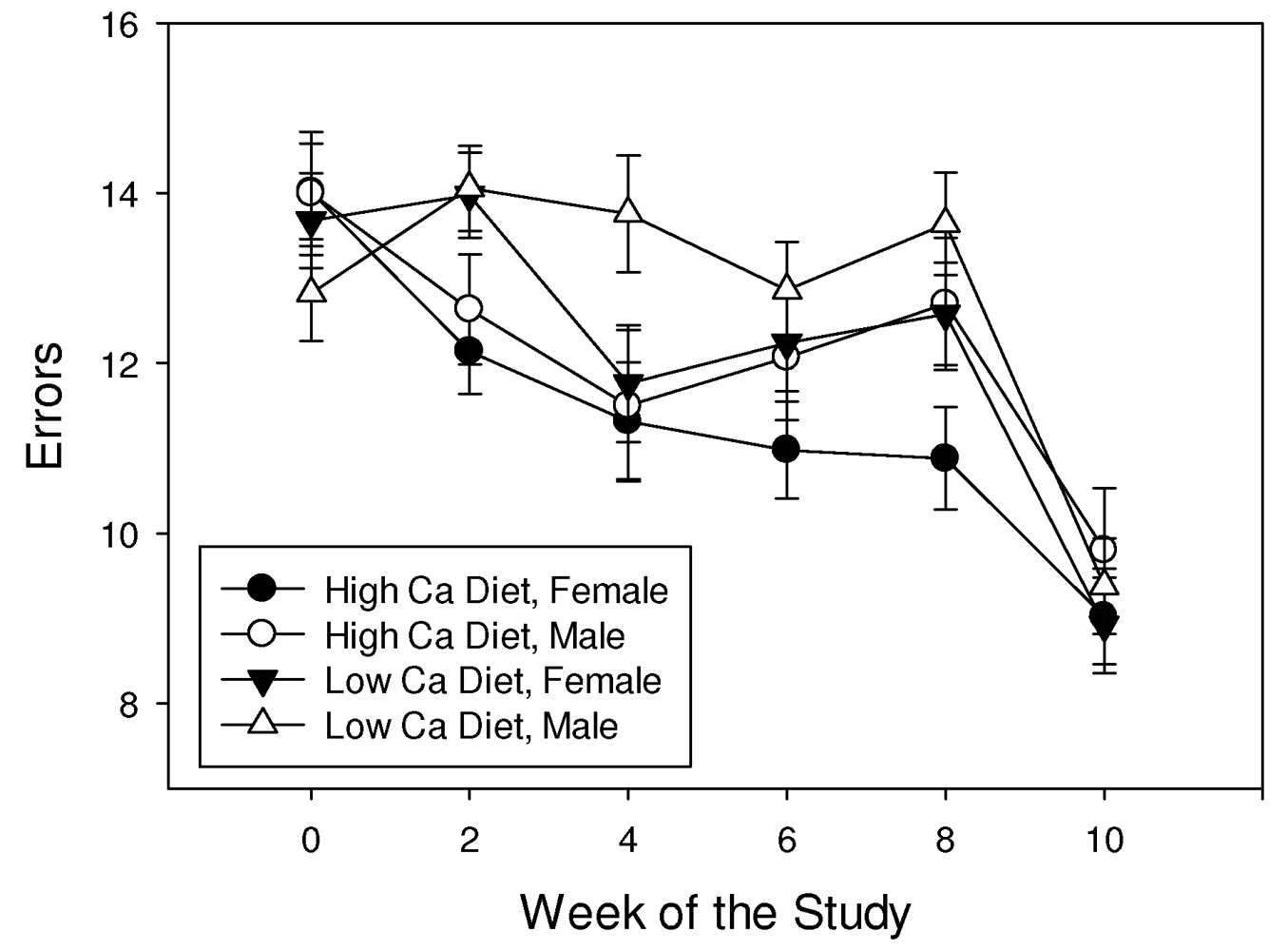
3.3. Shrew Mass Fluctuation
| Treatment | Starting Mass (g) ± SE | Ending Mass (g) ± SE |
|---|---|---|
| Females High Ca | 6.10 ± 0.30 | 6.14 ± 0.30 |
| Males High Ca | 5.65 ± 0.92 | 5.80 ± 0.71 |
| Females Low Ca | 5.84 ± 0.26 | 5.34 ± 0.30 |
| Males Low Ca | 5.20 ± 0.16 | 5.02 ± 0.17 |
4. Discussion
5. Conclusions
Acknowledgments
References
- Driscoll, C.T.; Lawrence, G.B.; Bulger, A.J.; Butler, T.J.; Cronan, C.S.; Eagar, C.; Lambert, K.F.; Likens, G.E.; Stoddard, J.L.; Weathers, K.C. Acidic deposition in the northeastern United States: Sources and inputs, ecosystem effects, and management strategies. Bioscience 2001, 51, 180–198. [Google Scholar] [CrossRef]
- Scheuhammer, A.M. Effects of acidification on the availability of toxic metals and calcium to wild birds and mammals. Environ. Pollut. 1991, 71, 329–375. [Google Scholar] [CrossRef]
- Graveland, J.; Drent, R.H. Calcium availability limits breeding success of passerines on poor soils. J. Anim. Ecol. 1997, 66, 279–288. [Google Scholar] [CrossRef]
- Graveland, J.; Wal, R. Decline in snail abundance due to soil acidification causes eggshell defects in forest passerines. Oecologia 1996, 105, 351–360. [Google Scholar] [CrossRef]
- Hotopp, K.P. Land snails and soil calcium in central Appalachian mountain forest. Southeast. Nat. 2002, 1, 27–44. [Google Scholar] [CrossRef]
- Taliaferro, E.H.; Holmes, R.T.; Blum, J.D. Eggshell characteristics and calcium demands of a migratory songbird breeding in two New England forests. Wilson Bull. 2001, 113, 94–100. [Google Scholar] [CrossRef]
- Hames, R.S.; Rosenberg, K.V.; Lowe, J.D.; Barker, S.E.; Dhondt, A.A. Adverse effects of acid rain on the distribution of the wood thrush Hylocichla mustelina in North America. Proc. Natl. Acad. Sci. USA 2002, 99, 11235–11240. [Google Scholar]
- Blancher, P.J.; McNicol, D.K. Breeding biology of tree swallows in relation to wetland acidity. Can. J. Zool. 1988, 66, 842–849. [Google Scholar] [CrossRef]
- Graveland, J.; van Gijzen, T. Arthropods and seeds are not sufficient as calcium sources for shell formation and skeletal growth in passerines. Ardea 1994, 82, 299–314. [Google Scholar]
- Levy, S. A plague of deer. BioScience 2006, 56, 718–721. [Google Scholar] [CrossRef]
- McShea, W.J.; Healy, W.M.; Devers, P.; Fearer, T.; Koch, F.H.; Stauffer, D.; Waldon, J. Forestry matters: Decline of oaks will impact wildlife in hardwood forests. J. Wildl. Manag. 2007, 71, 1717–1728. [Google Scholar] [CrossRef]
- Simkiss, K. Calcium in Reproductive Physiology: A Comparative Study of Vertebrates; Chapman and Hall: New York, NY, USA, 1967; pp. 1–278. [Google Scholar]
- Kwiecinski, G.G.; Falzone, M.; Studier, E.H. Milk concentration and postnatal accretion of minerals and nitrogen in two phyllostomid bats. J. Mammal. 2003, 84, 926–936. [Google Scholar] [CrossRef]
- Studier, E.H.; Kunz, T.H. Accretion of nitrogen and minerals in suckling bats, Myotis velifer and Tadarida brasiliensis. J. Mammal. 1995, 76, 32–42. [Google Scholar] [CrossRef]
- Studier, E.H.; Sevick, S.H.; Keeler, J.O.; Schenck, R.A. Nutrient levels in guano from maternity colonies of big brown bats. J. Mammal. 1994, 75, 71–83. [Google Scholar] [CrossRef]
- Ingram, W.M. Snail associates of Blarina brevicauda talpoides (say). J. Mammal. 1942, 23, 255–258. [Google Scholar] [CrossRef]
- Smart, J.L.; Billing, A.E.; Duggan, J.P.; Massey, R.F. Effects of early life undernutrition in artificially-reared rats: 3. Further studies of growth and behaviour. Physiol. Behav. 1989, 45, 1153–1160. [Google Scholar] [CrossRef]
- De Jonge, W.J.; Marescau, B.; D’Hooge, R.; de Deyn, P.P.; Hallemeeschi, M.M.; Deutz, N.E.P.; Ruijter, J.M.; Lamers, W.H. Overexpression of arginase alters circulating and tissue amino acids and guanidino compounds and affects neuromotor behavior in mice. J. Nutr. 2001, 131, 2732–2740. [Google Scholar]
- Bernard, R.T.F.; Kerley, G.I.H.; Doubell, T.; Davison, A. Reproduction in the round-eared elephant shrew (Macroscelides proboscideus) in the southern Karoo, South Africa. J. Zool. 1996, 240, 233–243. [Google Scholar] [CrossRef]
- Batzli, G.O. Nutritional ecology of the California vole: Effects of food quality on reproduction. Ecology 1986, 67, 406–412. [Google Scholar] [CrossRef]
- Colombo, P.J.; Wetsel, W.C.; Gallagher, M. Spatial memory is related to hippocampal subcellular concentrations of calcium-dependent protein kinase C isoforms in young and aged rats. Proc. Natl. Acad. Sci. USA 1997, 94, 14195–14199. [Google Scholar]
- Punzo, F. Early-life nutritional environment and spatial navigation in the water shrew, Sorex palustris (Insectivora). J. Environ. Biol. 2004, 25, 403–411. [Google Scholar]
- Sato, T.; Teramoto, T.; Tanaka, K.; Ohnishi, Y.; Irifune, M.; Nishikawa, T. Effects of ovariectomy and calcium deficiency on learning and memory of eight-arm radial maze in middle-aged female rats. Behav. Brain Res. 2003, 142, 207–216. [Google Scholar] [CrossRef]
- Stamps, J. Motor learning and the value of familiar space. Am. Nat. 1995, 146, 41–58. [Google Scholar]
- Denisova, N.A.; Shukitt-Hale, B.; Rabin, B.M.; Joseph, J.A. Brain signaling and behavioral responses induced by exposure to 56Fe-particle radiation. Radiat. Res. 2002, 158, 725–734. [Google Scholar] [CrossRef]
- Whitaker, J.O. Cryptotis parva. Mamm. Species 1974, 43, 1–8. [Google Scholar] [CrossRef]
- Mock, O.B. The Care and Use of Laboratory Least Shrews, Cryptotis parva. In Advances in the Biology of Shrews II; Merritt, J.F., Churchfield, S., Hutterer, R., Sheftel, B.I., Eds.; International Society of Shrew Biologists: New York, NY, USA, 2005; pp. 439–448, no. 1. [Google Scholar]
- Armitage, J.M.; Gobas, F.A.P.C. A terrestrial food-chain bioaccumulation model for POPs. Environ. Sci. Technol. 2007, 41, 4019–4025. [Google Scholar] [CrossRef]
- Hughes, L.; Mackay, D. Model of the fate of chemicals in sludge-amended soils with uptake in vegetation and soil-dwelling organisms. Soil Sediment Contam. 2011, 20, 938–960. [Google Scholar]
- Hoffmann, A.A.; Daborn, P.J. Towards genetic markers in animal populations as biomonitors for human-induced environmental change. Ecol. Lett. 2007, 10, 63–76. [Google Scholar] [CrossRef]
- Formanowicz, D.R., Jr; Bradley, P.J.; Brodie, E.D., Jr. Food hoarding by the least shrew (Cryptotis parva): Intersexual and prey type effects. Am. Midl. Nat. 1989, 122, 26–33. [Google Scholar]
- Mock, O.B. The least shrew (Cryptotis parva) as a laboratory animal. Lab Anim. Sci. 1982, 32, 177–179. [Google Scholar]
- Klasing, K.C.; Thacker, P.; Lopez, M.A.; Calvert, C.C. Increasing the calcium content of mealworms (Tenebrio molitor) to improve their nutritional value for bone mineralization of growing chicks. J. Zoo Wildl. Med. 2000, 31, 512–517. [Google Scholar]
- Studier, E.H.; Sevick, S.H. Live mass, water content, nitrogen and mineral levels in some insects from south-central lower Michigan. Comp. Biochem. A 1992, 103, 579–595. [Google Scholar] [CrossRef]
- Punzo, F.; Chavez, S. Effect of aging on spatial learning and running speed in the shrew (Cryptotis parva). J. Mamm. 2003, 84, 1112–1120. [Google Scholar] [CrossRef]
- Milliken, G.A.; Johnson, D.E. Analysis of Messy Data, Volume 1: Designed Experiments; Wadsworth: Belmont, CA, USA, 1994; p. 490. [Google Scholar]
- Erdfelder, E.; Faul, F.; Buchner, A. GPOWER: A general power analysis program. Behav. Res. Methods Instrum. Comput. 1996, 28, 1–11. [Google Scholar] [CrossRef]
- Glade, M.J. A 21st century evaluation of the safety of oral vitamin D. Nutrition 2012, 28, 344–356. [Google Scholar] [CrossRef]
- White, J.A.; McKinney, B.C.; John, M.C.; Powers, P.A.; Kamp, J.; Murphy, G.G. Conditional forebrain deletion of the L-type calcium channel Ca V 1.2 disrupts remote spatial memories in mice. Learn Memory 2008, 15, 1–5. [Google Scholar] [CrossRef]
- Whittenberg, G.M.; Tsien, J.Z. An emerging molecular and cellular framework for memory processing by the hippocampus. Trends Neurosci. 2002, 25, 501–505. [Google Scholar] [CrossRef]
- Swanson, L. The Hippocampus and the Concept of the Limbic System. In Neurobiology of Hippocampus; Seffert, W., Ed.; Academic Press: London, UK, 1983; pp. 3–19. [Google Scholar]
- Talbott, S.M.; Rothkopf, M.M.; Shapses, S.A. Dietary restriction of energy and calcium alters bone turnover and density in younger and older female rats. J. Nutr. 1998, 128, 640–645. [Google Scholar]
- Guessous, I.; Bochud, M.; Bonny, O.; Burnier, M. Calcium, Vitamin D and cardiovascular disease. Kidney Blood Press. Res. 2011, 34, 404–417. [Google Scholar] [CrossRef]
- Wilkins, C.H.; Birge, S.J.; Sheline, Y.I.; Morris, J.C. Vitamin D deficiency is associated with worse cognitive performance and lower bone density in older African Americans. J. Natl. Med. Assoc. 2009, 101, 349–354. [Google Scholar]
- Bischoff, H.A.; Stahelin, H.B.; Dick, W.; Akos, R.; Knecht, M.; Salis, C.; Nebiker, M.; Theiler, R.; Pfeifer, M.; Begerow, B.; et al. Effects of vitamin D and calcium supplementation on falls: A randomized controlled trial. J. Bone Miner. Res. 2003, 18, 343–351. [Google Scholar] [CrossRef]
- Swift, S.N.; Baek, K.; Swift, J.M.; Bloomfield, S.A. Restriction of dietary energy intake has a greater impact on bone integrity than does restriction of calcium in exercising female rats. J. Nutr. 2012, 142, 1038–1045. [Google Scholar] [CrossRef]
- Welch, J.M.; Turner, C.H.; Devareddy, L.; Arjmandi, B.H.; Weaver, C.M. High impact exercise is more beneficial than dietary calcium for building bone strength in the growing rat skeleton. Bone 2008, 42, 660–668. [Google Scholar] [CrossRef]
- Alaei, H.; Moloudi, R.; Sarkaki, A.R. Effects of treadmill running on mid-term memory and swim speed in the rat with Morris water maze test. J. Bodyw. Mov. Ther. 2008, 12, 72–75. [Google Scholar] [CrossRef]
- Siddiqui, S.M.K.; Chang, E.; Li, J.; Burlage, C.; Zou, M.; Buhman, K.K.; Koser, S.; Donkin, S.S.; Teegarden, D. Dietary intervention with vitamin D, calcium, and whey protein reduced fat mass and increased lean mass in rats. Nutr. Res. 2008, 28, 783–790. [Google Scholar] [CrossRef]
- Zemel, M.B. Proposed role of calcium and dairy food components in weight management and metabolic health. Phys. Sportsmed. 2009, 37, 29–39. [Google Scholar] [CrossRef]
- Davey, G. Ecological Learning Theory; Taylor & Frances/Routledge: Oxfordshire, UK, 1989; p. 392. [Google Scholar]
- Rado, R.; Shanas, U.; Zuri, I.; Terkel, J. Seasonal activity in the blind mole rat (Spalax ehrenbergi). Can. J. Zool. 1993, 71, 1733–1737. [Google Scholar] [CrossRef]
- Vleck, D. The energy cost of burrowing by the pocket gopher Thomomys bottae. Physiol. Zool. 1979, 52, 122–136. [Google Scholar]
- Churchfield, S. The Natural History of Shrews; Cornell University Press: Ithaca, NY, USA, 1990; p. 178. [Google Scholar]
- Korpimaki, E. Predation causing synchronous decline phases in microtine and shrew populations in western Finland. Oikos 1986, 46, 124–127. [Google Scholar] [CrossRef]
- Barkley, C.L.; Jacobs, L.F. Sex and species differences in spatial memory in food-storing kangaroo rats. Anim. Behav. 2007, 73, 321–329. [Google Scholar] [CrossRef]
- Squire, L.R.; Knowlton, B.J. Memory, Hippocampus, and Brain Systems. In The Cognitive Neurosciences; Gazzinga, M., Ed.; MIT Press: Cambridge, MA, USA, 1994; pp. 825–837. [Google Scholar]
- Lynch, M.A. Long-term potentiation and memory. Physiol. Rev. 2003, 84, 87–136. [Google Scholar] [CrossRef]
- Silva, A.J.; Paylor, R.; Wehner, J.M.; Tonegawa, S. Impaired spatial learning in alpha-calcium-calmodulin kinase II mutant mice. Science 1992, 257, 206–211. [Google Scholar]
- Silva, A.J.; Rosahl, T.W.; Chapman, P.F.; Marowitz, Z.; Friedman, E.; Frankland, P.W.; Cestari, V.; Cioffi, D.; Sudhof, T.C.; Bourtchuladze, R. Impaired learning in mice with abnormal short-lived plasticity. Curr. Biol. 1996, 6, 1509–1517. [Google Scholar] [CrossRef]
- Johansen, J.P.; Cain, C.K.; Ostroff, L.E.; le Doux, J.E. Molecular mechanisms of fear and learning. Cell 2011, 147, 509–524. [Google Scholar] [CrossRef]
- Kurta, A.; Bell, G.P.; Nagy, K.A.; Kunz, T.H. Energetics of pregnancy and lactation in free ranging little brown bats (Myotis lucifugus). Physiol. Zool. 1989, 62, 804–818. [Google Scholar]
- Speakman, J.R. The physiological costs of reproduction in small mammals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 375–398. [Google Scholar] [CrossRef]
- Gaulin, S.J.C.; Fitzgerald, R.W. Sexual selection for spatial-learning ability. Anim. Behav. 1989, 37, 322–331. [Google Scholar] [CrossRef]
- Gresack, J.E.; Frick, K.M. Male mice exhibit better spatial working and reference memory than females in a water-escape radial arm maze task. Brain Res. 2003, 982, 98–107. [Google Scholar] [CrossRef]
- LaBuda, C.J.; Mellgren, R.L.; Hale, R.L. Sex differences in the acquisition of a radial maze task in the CD-1 mouse. Physiol. Behav. 2002, 76, 213–217. [Google Scholar] [CrossRef]
- Root, R.L. Neonatal exogenous testosterone modifies sex differences in radial arm and Morris water maze performance in prepubescent and adult rats. Behav. Brain Res. 1993, 53, 1–10. [Google Scholar] [CrossRef]
- Williams, C.; Meck, W.H. The organizational effects of gonadal steroid on sexually dimorphic spatial abilty. Pseudoneuroendocrinology 1991, 16, 155–176. [Google Scholar] [CrossRef]
- Jonasson, Z. Meta-analysis of sex differences in rodent models of learning and memory: A review of behavioral and biological data. Neurosci. Biobehav. Rev. 2005, 28, 811–825. [Google Scholar] [CrossRef]
- Galea, L.A.; Kavaliers, M.; Ossenkopp, K.P. Sexually dimorphic spatial learning in meadow voles Microtus pennsylvanicus and deer mice Peromyscus maniculatus. J. Exp. Biol. 1996, 199, 195–200. [Google Scholar]
- Perrot-Sinal, T.S.; Kostenuik, M.A.; Ossenkopp, K.P.; Kavaliers, M. Sex differences in performance in the Morris water maze and the effects of initial nonstationary hidden platform training. Behav. Neurosci. 1996, 110, 1309–1320. [Google Scholar] [CrossRef]
- Gaulin, S.J.C.; Fitzgerald, R.W.; Wartell, M.S. Sex differences in spatial ability: An evolutionary hypothesis and test. Am. Nat. 1990, 127, 72–88. [Google Scholar]
- Beatty, W.W. Gonadal hormones and sex differences in non-reproductive behaviors in rodents: Organizational and activational influences. Horm. Behav. 1979, 12, 112–163. [Google Scholar] [CrossRef]
- Sandstrom, N.L.; Kaufman, J.; Huettel, S.A. Males and females use different distal cues in a virtual environment navigation task. Cogn. Brain Res. 1998, 6, 351–360. [Google Scholar] [CrossRef]
- Bimonte, H.A.; Hyde, L.A.; Hoplight, V.H.; Denenberg, V.H. In two species, females exhibit superior working memory and inferior reference memory on the water radial-arm maze. Physiol. Behav. 2000, 70, 311–317. [Google Scholar] [CrossRef]
- Rendeiro, C.; Spencer, J.P.E.; Vauzour, D.; Butler, L.T.; Ellis, J.A.; Williams, C. The impact of flavinoids on spatial memory in rodents: From behaviour to underlying hippocampal mechanisms. Genes Nutr. 2009, 4, 251–270. [Google Scholar] [CrossRef]
- Beck, K.D.; Luine, V. Sex differences in behavioral and neurochemical profiles after chronic stress: Role of housing conditions. Physiol. Behav. 2002, 75, 661–673. [Google Scholar] [CrossRef]
- Gibbs, R.B. Estrogen replacement enhances acquisition of a spatial memory task and reduces deficits associated with hippocampal muscarinic receptor inhibition. Horm. Behav. 1999, 36, 222–233. [Google Scholar] [CrossRef]
- Friedland, A.J.; Miller, E.K. Major-element cycling in a high-elevation Adirondack forest: Patterns and changes, 1986-1996. Ecol. Appl. 1999, 9, 958–967. [Google Scholar]
- Yanai, R.D.; Siccama, T.G.; Arthur, M.A.; Federer, C.A.; Friedland, A.J. Accumulation and depletion of base cations in forest floors in the northeastern United States. Ecology 1999, 80, 2774–2787. [Google Scholar] [CrossRef]
- Tilgar, V.; Mänd, R.; Mägi, M. Calcium shortage as a constraint on reproduction in great tits Parus major: A field experiment. J. Avian Biol. 2002, 33, 407–413. [Google Scholar] [CrossRef]
- James, F.C.; McCulloch, C.E.; Wiedenfeld, D.A. New approaches to the analysis of population trends in land birds. Ecology 1996, 77, 13–27. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Czajka, J.L.; McCay, T.S.; Garneau, D.E. Physical and Cognitive Performance of the Least Shrew (Cryptotis parva) on a Calcium-Restricted Diet. Behav. Sci. 2012, 2, 172-185. https://doi.org/10.3390/bs2030172
Czajka JL, McCay TS, Garneau DE. Physical and Cognitive Performance of the Least Shrew (Cryptotis parva) on a Calcium-Restricted Diet. Behavioral Sciences. 2012; 2(3):172-185. https://doi.org/10.3390/bs2030172
Chicago/Turabian StyleCzajka, Jessica L., Timothy S. McCay, and Danielle E. Garneau. 2012. "Physical and Cognitive Performance of the Least Shrew (Cryptotis parva) on a Calcium-Restricted Diet" Behavioral Sciences 2, no. 3: 172-185. https://doi.org/10.3390/bs2030172
APA StyleCzajka, J. L., McCay, T. S., & Garneau, D. E. (2012). Physical and Cognitive Performance of the Least Shrew (Cryptotis parva) on a Calcium-Restricted Diet. Behavioral Sciences, 2(3), 172-185. https://doi.org/10.3390/bs2030172