Bythocythere solisdeus n. sp. and Cytheropteron eleonorae n. sp. (Crustacea, Ostracoda) from the Early Pleistocene Bathyal Sediments of Cape Milazzo (NE, Sicily)

Abstract
:1. Introduction
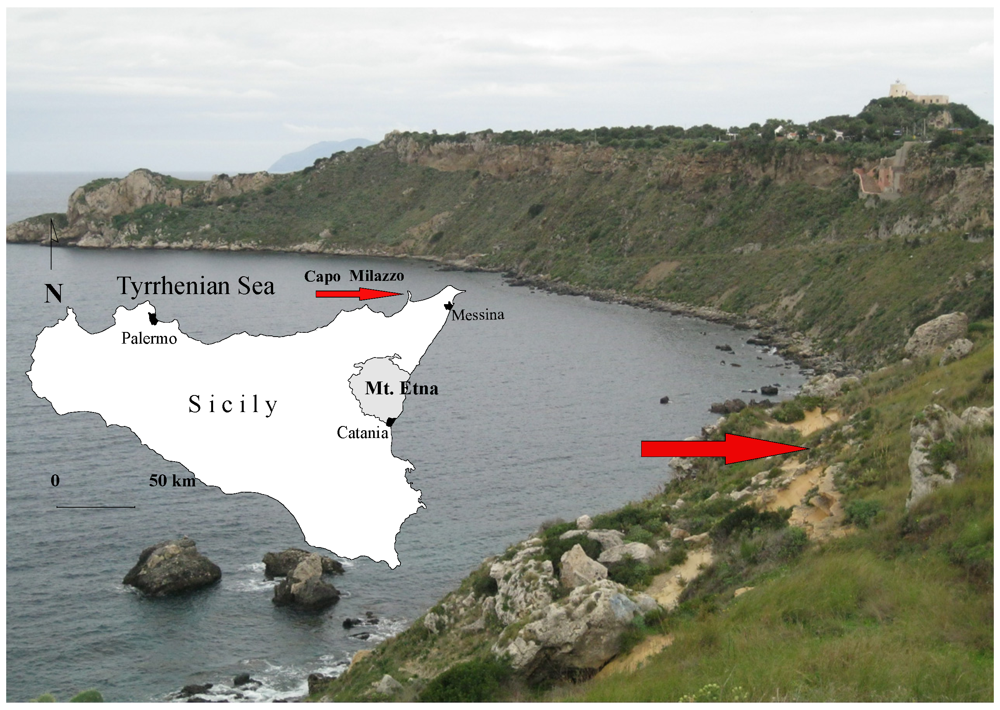


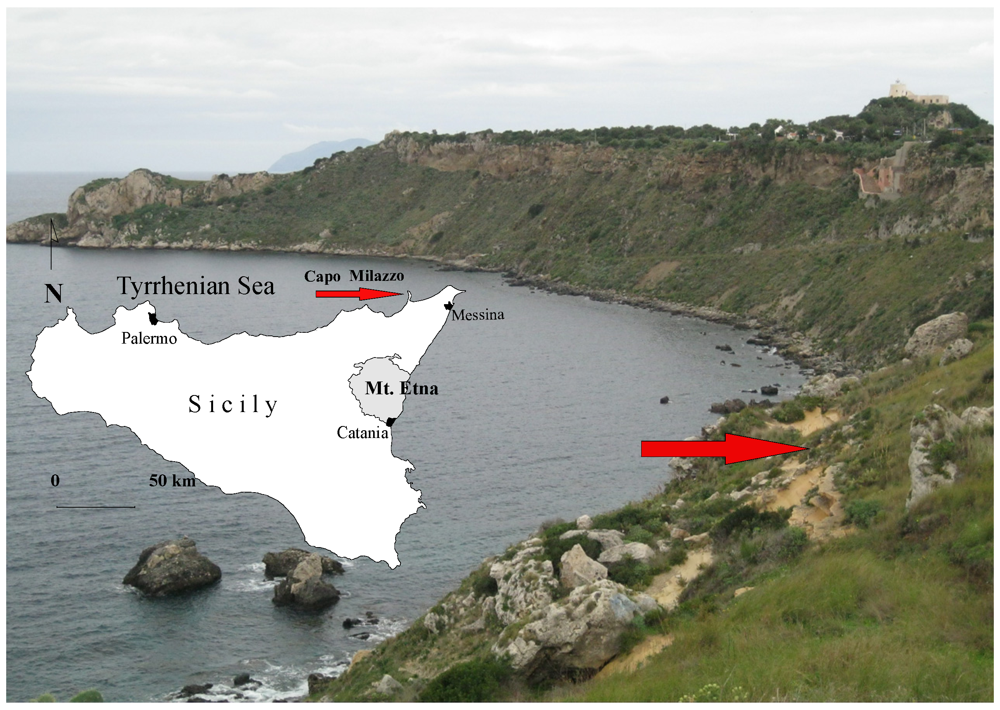{kind=link}
{kind=link}
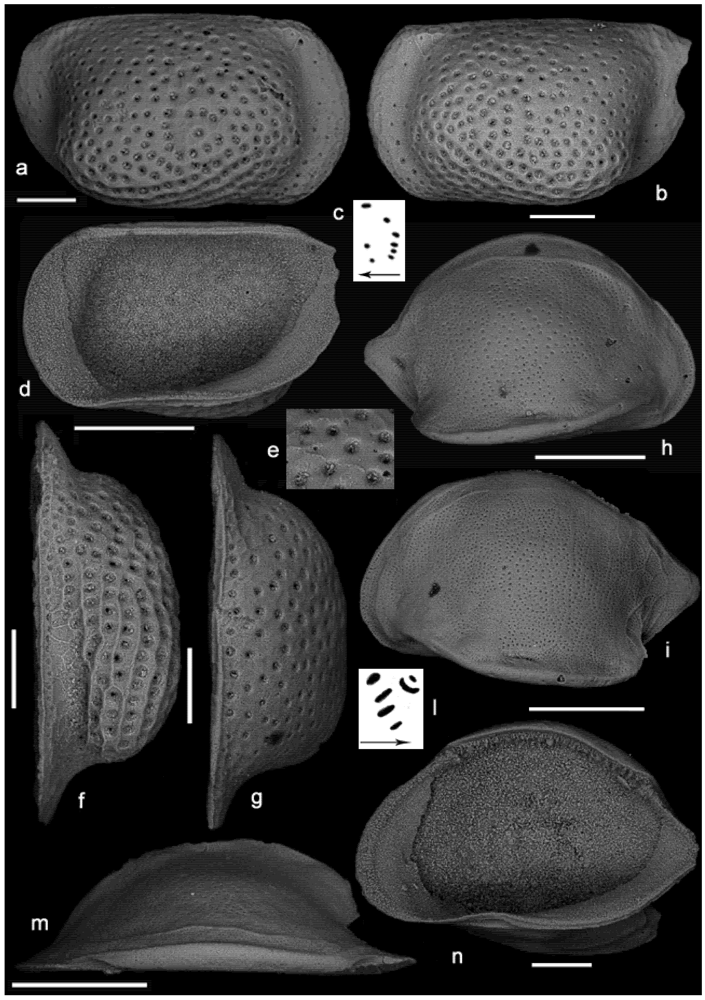{kind=link}
| OSTRACOD SPECIES | n. spec. | % | OSTRACOD SPECIES | n. spec. | % |
|---|---|---|---|---|---|
| Bythocypris obtusata (SARS) | 45 | 19.31 | Eucythere curta RUGGIERI | 2 | 0.86 |
| Bairdia conformis (TERQUEM) | 25 | 10.73 | Paracytherois spp. | 2 | 0.86 |
| Cytherella vulgatella AIELLO et al. | 20 | 8.58 | Paradoxostoma spp. | 2 | 0.86 |
| Henryhowella sarsii profunda BONADUCE et al. | 20 | 8.58 | Pontocypris acuminata (MUELLER) | 2 | 0.86 |
| Cytheropteron eleonare n. sp. | 17 | 7.30 | Pseudocythere armata BONADUCE et al. | 2 | 0.86 |
| Bythocythere solisdeus n. sp. | 8 | 3.43 | Argilloecia acuminata MUELLER | 1 | 0.43 |
| Pseudocythere caudata SARS | 7 | 3.00 | Argilloecia micra BONADUCE et al. | 1 | 0.43 |
| Krithe compressa (SEGUENZA) | 7 | 3.00 | Argilloecia robusta BONADUCE et al. | 1 | 0.43 |
| Bythocypris bosquetiana (BRADY) | 6 | 2.58 | Aurila lanceaeformis* ULICZNY | 1 | 0.43 |
| Cytheropteron lancei CARBONEL | 6 | 2.58 | Cytheropteron bifidum COLALONGO & PASINI | 1 | 0.43 |
| Krithe spp. | 6 | 2.58 | Cytheropteron pinarense gillesi AIELLO et al. | 1 | 0.43 |
| Anchistrocheles interrupta AIELLO et al. | 5 | 2.15 | Cytheropteron pseudoalatum COLALONGO & PASINI | 1 | 0.43 |
| Cytheropteron ionicum COLALONGO & PASINI | 5 | 2.15 | Cytheropteron testudo SARS | 1 | 0.43 |
| Sclerochilus gr.contortus (NORMAN) | 5 | 2.15 | Eucythere cf. E. triangula WHATLEY & COLES | 1 | 0.43 |
| Aurila impressa* RUGGIERI | 4 | 1.72 | Eucytherura sp. | 1 | 0.43 |
| Cytheropteron pinnatum COLALONGO & PASINI | 3 | 1.29 | Heterocythereis albomaculata* (BAIRD) | 1 | 0.43 |
| Sclerochilus spp. | 3 | 1.29 | Macrocypris sp. | 1 | 0.43 |
| Krithe frutex ABATE et al. | 3 | 1.29 | Paijenborchella malaiensis cymbula RUGGIERI | 1 | 0.43 |
| Aurila spp.* | 2 | 0.86 | Pedicythere phryne BONADUCE et al. | 1 | 0.43 |
| Bythoceratina scaberrima BENSON & SILV.- BRADLEY | 2 | 0.86 | Profundobythere sp. | 1 | 0.43 |
| Cytheropteron cf. C. venustum BONADUCE et al. | 2 | 0.86 | Quasibuntonia radiatopora (SEGUENZA) | 1 | 0.43 |
| Cytheropteron circumactum COLALONGO & PASINI | 2 | 0.86 | Tenedocythere prava* (BAIRD) | 1 | 0.43 |
| Cytheropteron triangulum COLALONGO & PASINI | 2 | 0.86 | Typhloeucytherura calabra COLALONGO & PASINI | 1 | 0.43 |
| Total | 233 | 100.00 |
2. Systematics

| Bythocythere solisdeus n. sp. | Cytheropteron eleonorae n. sp. | ||||||
|---|---|---|---|---|---|---|---|
| L | H | L/H | L | H | L/H | ||
| LV | 560 | 355 | 1.577465 | LV | 623.27 | 396.47 | 1.572048 |
| LV | 538.31 | 298.39 | 1.804048 | LV | 662.66 | 405.77 | 1.633093 |
| LV | 575.31 | 316.71 | 1.81652 | LV | 627.89 | 396.27 | 1.5845 |
| LV | 547.81 | 308.24 | 1.777219 | RV | 661.84 | 396.62 | 1.668701 |
| RV | 577.33 | 327.14 | 1.76478 | RV | 600 | 405 | 1.481481 |
| RV | 580.47 | 332.14 | 1.747667 | RV | 658.92 | 428.57 | 1.537485 |
| RV | 575.24 | 336.02 | 1.711922 | RV | 668.72 | 410.18 | 1.630309 |
| RV | 584.29 | 343.01 | 1.70342 | ||||
| Mean LV | 555.3575 | 319.585 | 637.94 | 399.5033 | |||
| Mean RV | 579.3325 | 334.5775 | 647.37 | 410.0925 | |||
| LV | 15.99205 | 24.76843 | 21.53242 | 5.428014 | |||
| RV | 3.942473 | 6.694417 | 31.8461 | 13.52612 | |||
3. Discussion and Conclusions
Acknowledgments
References
- Violanti, D.I. Foraminiferi plio-pleistocenici di Capo Milazzo. Boll. Mus. Reg. Sc. Nat. T. 1988, 6, 359–392. [Google Scholar]
- Fois, E. La successione neogenica di Capo Milazzo (Sicilia nord-orientale). Riv. Ital. Paleontol. S. 1990, 95, 397–440. [Google Scholar]
- Fois, E. Stratigraphy and palaeogeography of the Capo Milazzo area (NE Sicily, Italy): Clues to the evolution of the southern margin of the Tyrrhenian Basin during the Neogene. Palaeogeogr Palaeoclimatol. Palaeoecol. 1990, 78, 87–108. [Google Scholar] [CrossRef]
- Sciuto, F. Dati preliminari sulla ostracofauna pliocenica di Capo Milazzo (Sicilia NE). Boll. Soc. Paleontol. I. 2003, 42, 179–184. [Google Scholar]
- Sciuto, F. Ostracodi batiali pleistocenici di Capo Milazzo (Sicilia NE) ed implicazioni paleoambientali. Rend. Soc. Paleontolol. I. 2005, 2, 219–227. [Google Scholar]
- Sciuto, F. Bythocythere mylaensis n. sp. (Crustacea, Ostracoda) from Early Pleistocene of Capo Milazzo (NE Sicily). Boll. Soc. Paleontol. I. 2009, 48, 183–188. [Google Scholar]
- Gaetani, M.; Saccà, D. Brachiopodi batiali del Pliocene e del Pleistocene di Sicilia e Calabria. Riv. Ital. Paleontol. S. 1984, 90, 407–458. [Google Scholar]
- Rosso, A. Terataulopocella borealis gen. et sp. nov., a deep-water Pliocene Lekythoporid (Briozoa) from the Mediterranean Area. Mem. Sc. Geol. 2002, 54, 65–72. [Google Scholar]
- Sciuto, F. New ostracod species from Lower Pleistocene Bathyal sediments of Capo Milazzo (NE, Sicily). Boll. Soc. Paleontol. I. 2012. in submit. [Google Scholar]
- Benson, R.H. Ostracodes as indicators of threshold depth in the Mediterranean during the Pliocene. In The Mediterranean Sea: A Natural Sedimentation Laboratory; Stanley, D.J., Ed.; Stroudsburg, PA, USA, 1972; pp. 63–73. [Google Scholar]
- Benson, R.H. Estimating greater paleodepths with ostracodes, especially in past thermospheric oceans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984, 48, 107–141. [Google Scholar] [CrossRef]
- Dingle, R.V.; Lord, A.R. Benthic ostracods and deep water-masses in the Atlantic Ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 80, 213–235. [Google Scholar] [CrossRef]
- Catalano, S.; Di Stefano, A. Sollevamenti e tettogenisu pleistocenica lungo il margine tirrenico dei Monti Peloritani: Integrazione dei dati geomorfologici, strutturali e biostratigrafici. Il Quaternario 1997, 10, 337–342. [Google Scholar]
- Antonioli, F.; Kershaw, S.; Renda, P.; Rust, D.; Belluomini, G.; Cerasoli, M.; Radtke, U.; Silenzi, S. Elevation of the last interglacial highstand in Sicily (Italy): A benchmark of Coastal tectonic. Quarter. Int. 2006, 145-146, 3–18. [Google Scholar] [CrossRef]
- Stepanova, A.; Taldenkova, E.; Bauch, H.A. Recent ostracoda from Laptev Sea (Arctic Siberia): species assemblages and some environmental relationships. Mar. Micropaleontol. 2003, 48, 23–48. [Google Scholar] [CrossRef]
- Swanson, K.M.; Ayress, M.A. Cytheropteron testudo and related species from SW Pacific with analyses of their soft anatomics, relationships and distribution. Senck. Biol. 1999, 79, 151–193. [Google Scholar]
- Whatley, R.; Coles, G. The late Miocene to Quaternary Ostracoda of Leg 94, Deep Sea Drilling Project. Rev. Esp. Micropaleont. 1987, 19, 33–97. [Google Scholar]
- Aiello, G.; Barra, D.; Bonaduce, G. The genus Cytheropteron Sars, 1866 (Crustacea: Ostracoda) in the Pliocene-Early Pleistocene of the Mount San Nicola Section (Gela, Sicily. Micropaleontology 1996, 42, 167–178. [Google Scholar] [CrossRef]
- Moncharmont-Zei, M.; Russo, B.; Sgarrella, F.; Bonaduce, G.; Mascellaro, P. Paleoclimatic record from 4 cores (Gulf of Taranto, Ionian Sea). Evidence from Foraminifera and Ostracoda. Boll. Soc. Paleontol. I. 1985, 23, 21–51. [Google Scholar]
- Sars, G.O. An account of the Crustacea of Norway; Bergen Museum: Oslo, Norway, 1928; Volume IX. [Google Scholar]
- Puri, H.S.; Bonaduce, G.; Gervasio, A.M. Distribution of Ostracoda in the Mediterranean. In The Taxonomy, Morphology and Ecology of Recent Ostracoda; Neale, J.W., Ed.; Lubrecht & Cramer Ltd.: Edinburgh, UK, 1969; pp. 358–411. [Google Scholar]
- Malinverno, E.; Taviani, M.; Rosso, A.; Violanti, D.; Villa, I.; Savini, A.; Vertino, A.; Remia, A; Corselli, C. Stratigraphic framework of the Apulian deep-water coral province, Ionian Sea. Deep Sea Res. Pt. II Trop. Stud. Oceanogr. 2010, 57, 345–359. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sciuto, F. Bythocythere solisdeus n. sp. and Cytheropteron eleonorae n. sp. (Crustacea, Ostracoda) from the Early Pleistocene Bathyal Sediments of Cape Milazzo (NE, Sicily). Geosciences 2012, 2, 147-156. https://doi.org/10.3390/geosciences2030147
Sciuto F. Bythocythere solisdeus n. sp. and Cytheropteron eleonorae n. sp. (Crustacea, Ostracoda) from the Early Pleistocene Bathyal Sediments of Cape Milazzo (NE, Sicily). Geosciences. 2012; 2(3):147-156. https://doi.org/10.3390/geosciences2030147
Chicago/Turabian StyleSciuto, Francesco. 2012. "Bythocythere solisdeus n. sp. and Cytheropteron eleonorae n. sp. (Crustacea, Ostracoda) from the Early Pleistocene Bathyal Sediments of Cape Milazzo (NE, Sicily)" Geosciences 2, no. 3: 147-156. https://doi.org/10.3390/geosciences2030147