Human Attitude toward Reptiles: A Relationship between Fear, Disgust, and Aesthetic Preferences

Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Focused on reptiles
1.2. Research of Human Relationship to Animals
1.3. Negative Relationship to Animals
1.4. Positive Relationship to Animals
1.5. Aims
- What is the human attitude toward a group of reptiles in terms of positive and negative emotions and does our evaluation rely on basic morphotypes, specifically, are legless snakes perceived differently than the rest of reptiles?
- What is the relationship between perceived fear, disgust, and beauty of reptiles? Do these evaluations affect each other?
2. Materials and Methods
2.1. Selection and Preparation of Stimuli
2.2. Testing Emotional Response to Reptiles
2.3. Statistical Analyses
2.4. Ethical Note
3. Results
3.1. Agreement among Respondents
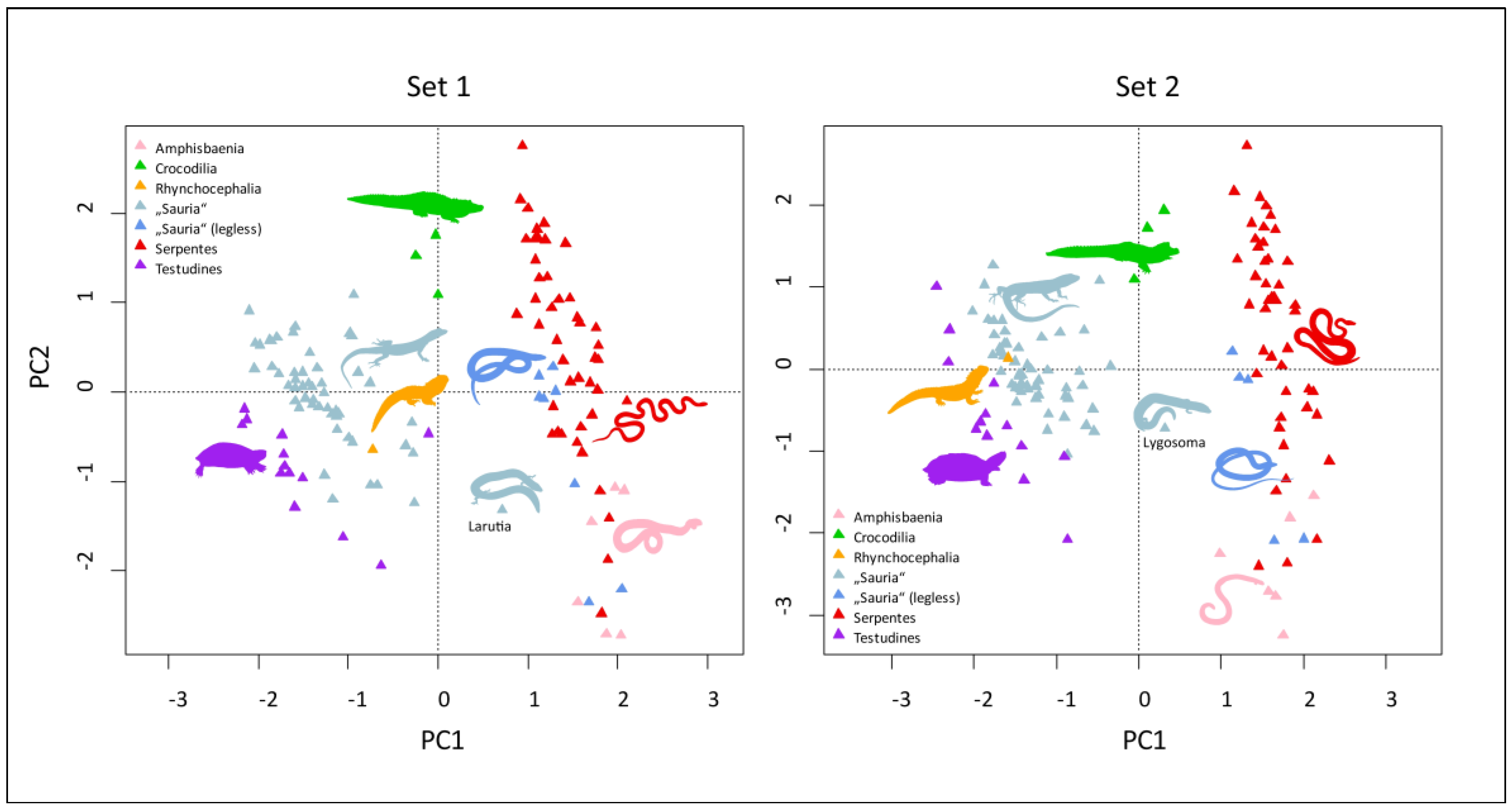
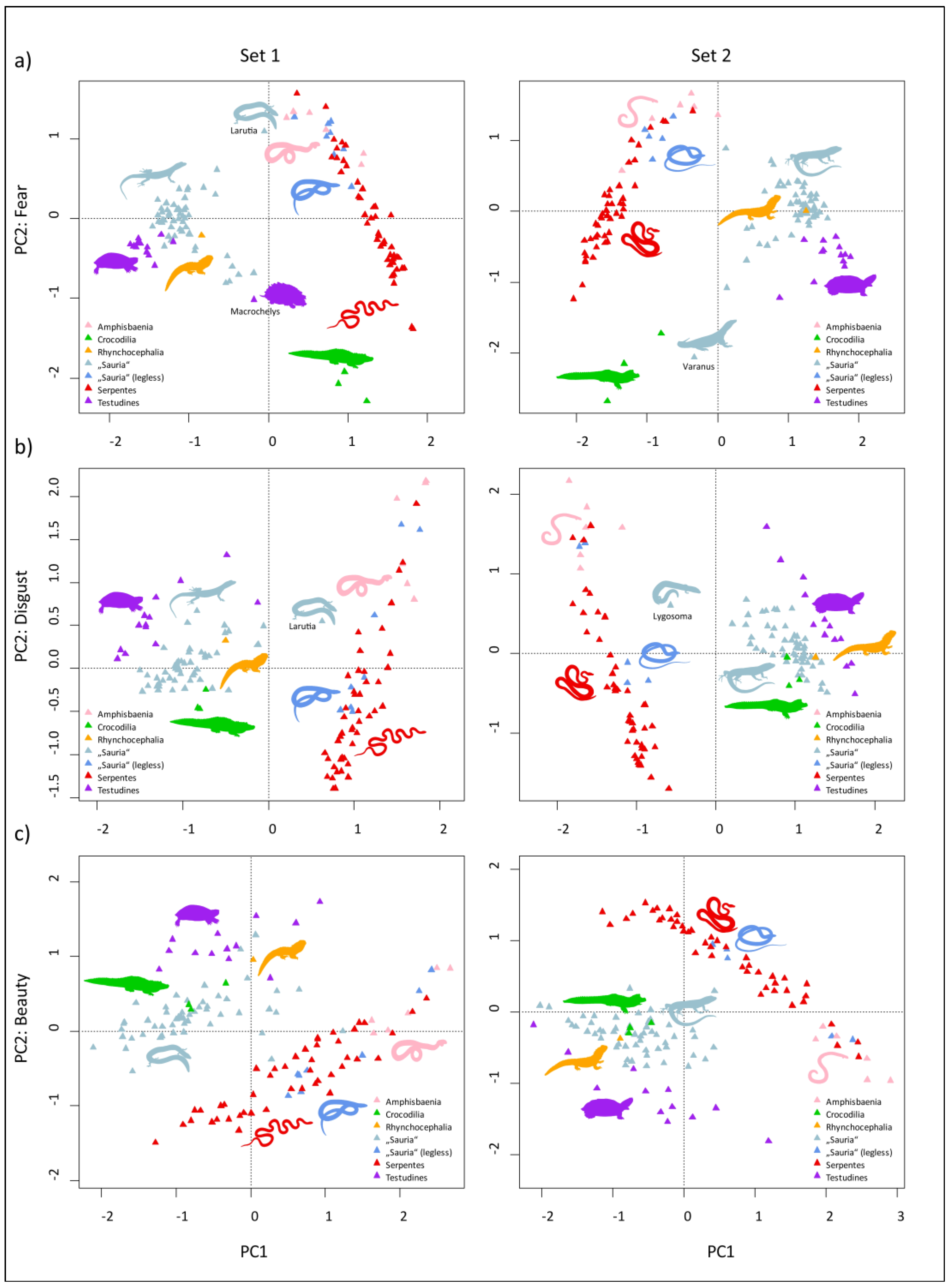
3.2. Multivariate Analyses of Datasets
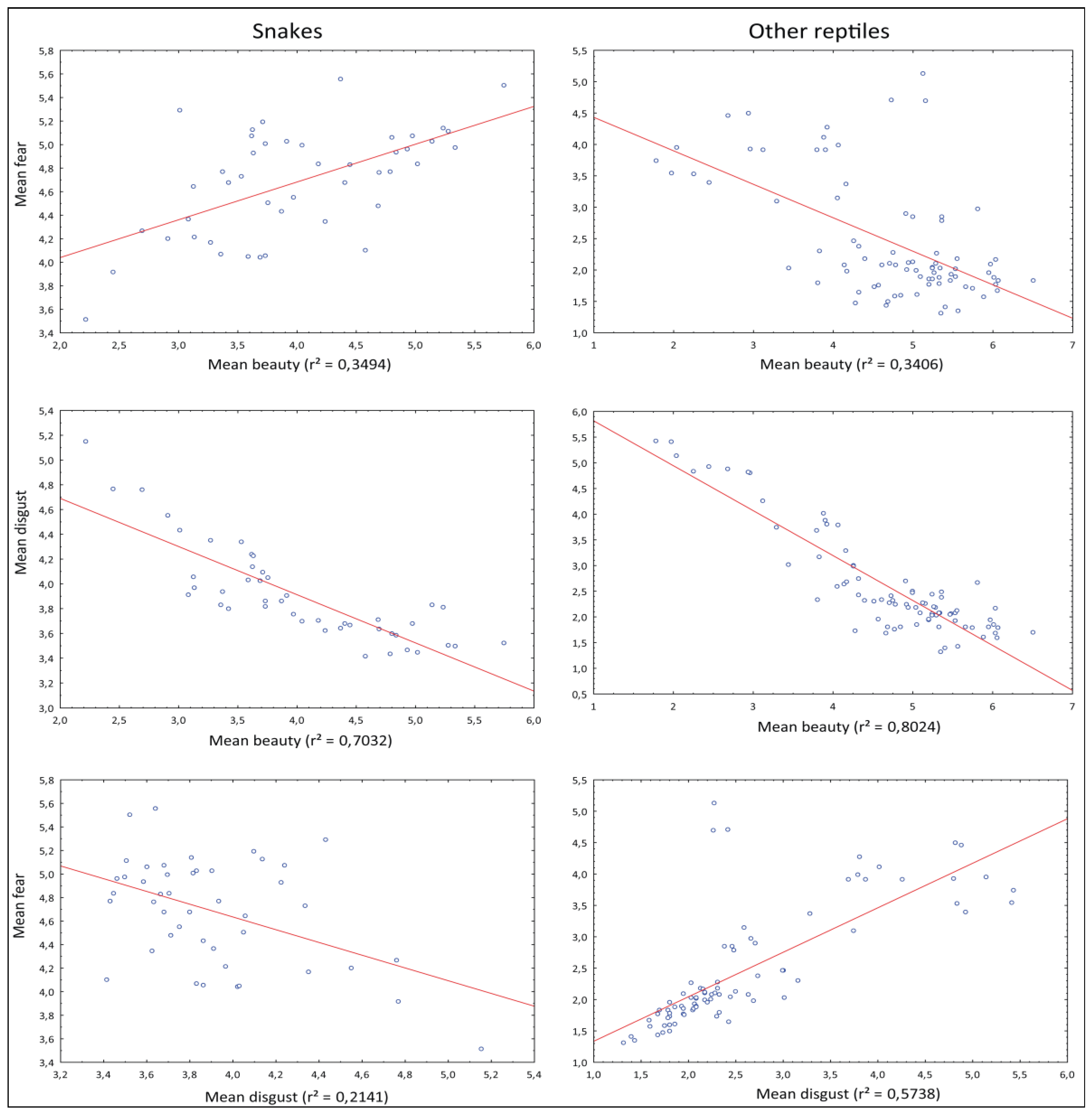
3.3. Mutual Correlation of the Fear, Disgust, and Beauty Ranks
4. Discussion
4.1. Relationship to Reptiles as a Group
4.2. Evaluation of Perceived Fear, Disgust, and Beauty
4.3. Mutual Relationship of the Ranks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rieppel, O. Turtles as diapsid reptiles. Nature 1996, 384, 453. [Google Scholar] [CrossRef]
- Lyson, T.R.; Sperling, E.A.; Heimberg, A.M.; Gauthier, J.A.; King, B.L.; Peterson, K.J. MicroRNAs support a turtle+ lizard clade. Biol. Lett. 2011, 8, 104–107. [Google Scholar] [CrossRef]
- Chiari, Y.; Cahais, V.; Galtier, N.; Delsuc, F. Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria). BMC Biol. 2012, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Linnæus, C. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio Decima, Reformata; Laurentii Salvii, Holmiæ: Stockholm, Sweden, 1758; pp. 1–4. 824p. [Google Scholar]
- Berlin, B. Ethnobiological Classification: Principles of Categorization of Plants and Animals in Traditional Societies; Princeton University Press: New Jersey, NY, USA, 2014; Volume 185. [Google Scholar]
- Uetz, P.; Freed, P.; Hosek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 14 November 2018).
- Bonnet, X.; Shine, R.; Lourdais, O. Taxonomic chauvinism. Trends Ecol. Evol. 2002, 17, 1–3. [Google Scholar] [CrossRef]
- Troudet, J.; Grandcolas, P.; Blin, A.; Vignes-Lebbe, R.; Legendre, F. Taxonomic bias in biodiversity data and societal preferences. Sci. Rep. 2017, 7, 9132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.A.; May, R.M. Taxonomic bias in conservation research. Science 2002, 297, 191–192. [Google Scholar] [CrossRef]
- Böhm, M.; Collen, B.; Baillie, J.E.; Bowles, P.; Chanson, J.; Cox, N.; Rhodin, A.G. The conservation status of the world’s reptiles. Biol. Conserv. 2013, 157, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.; Santana, G.; Almeida, W.; Neto, N.L.; Vieira, W. Reptiles used for medicinal and magic religious purposes in Brazil. Appl. Herpetol. 2009, 6, 257–274. [Google Scholar] [CrossRef]
- Schlegel, J.; Rupf, R. Attitudes towards potential animal flagship species in nature conservation: A survey among students of different educational institutions. J. Nat. Conserv. 2010, 18, 278–290. [Google Scholar] [CrossRef]
- Alves, R.R.N.; Vieira, K.S.; Santana, G.G.; Vieira, W.L.S.; Almeida, W.O.; Souto, W.M.S.; Pezzuti, J.C.B. A review on human attitudes towards reptiles in Brazil. Environ. Monit. Assess. 2012, 184, 6877–6901. [Google Scholar] [CrossRef]
- Ceriaco, L.M. Human attitudes towards herpetofauna: The influence of folklore and negative values on the conservation of amphibians and reptiles in Portugal. J. Ethnobiol. Ethnomed. 2012, 8, 8. [Google Scholar] [CrossRef]
- Alves, R.R. Relationships between fauna and people and the role of ethnozoology in animal conservation. Ethnobiol. Conserv. 2012, 1, 1–69. [Google Scholar] [CrossRef]
- Alves RR, N.; Souto WM, S. Ethnozoology: A brief introduction. Ethnobiol. Conserv. 2015, 4, 1–13. [Google Scholar] [CrossRef]
- Bertrand, H. Contribution à l'étude de l'herpétologie et de l'ethnoherpétologie en Anjou—A study on the herpetology and ethnoherpetology of Anjou province (France). Bull. Soc. Herpétol. Fr. 1997, 82–83, 51–62. [Google Scholar]
- Alves, R.R.N.; Alves, R.R.N.; Pereira Filho, G.A.; Vieira, K.S.; Souto, W.M.S.; Mendonça, L.E.T.; Montenegro, P.F.G.P.; Vieira, W.L.S. A zoological catalogue of hunted reptiles in the semiarid region of Brazil. J. Ethnobiol. Ethnomed. 2012, 8, 27. [Google Scholar] [CrossRef]
- Fernandes-Ferreira, H.; Mendonca, S.V.; Cruz, R.L.; Borges-Nojosa, D.M.; Nobrega Alves, R.R. Hunting of herpetofauna in montane, coastal, and dryland areas of Northeastern Brazil. Herpetol. Conserv. Biol. 2013, 8, 652–666. [Google Scholar]
- Mendonça, L.E.T.; Vieira, W.L.S.; Alves, R.R.N. Caatinga Ethnoherpetology: Relationships between herpetofauna and people in a semiarid region. Amphib. Rept. Conserv. 2014, 8, 24–32. [Google Scholar] [CrossRef]
- Wagler, R. The association between preservice elementary teacher animal attitude and likelihood of animal incorporation in future science curriculum. Int. J. Environ. Sci. Educ. 2010, 5, 353–375. [Google Scholar]
- Batt, S. Human attitudes towards animals in relation to species similarity to humans: A multivariate approach. Biosci. Horiz. 2009, 2, 180–190. [Google Scholar] [CrossRef]
- IUCN Red list. Available online: https://www.iucnredlist.org/resources/summary-statistics/ (accessed on 21 March 2019).
- Borgi, M.; Cirulli, F. Attitudes toward animals among kindergarten children: Species preferences. Anthrozoös 2015, 28, 45–59. [Google Scholar] [CrossRef]
- Alves, R.R.; Silva, V.N.; Trovão, D.M.; Oliveira, J.V.; Mourão, J.S.; Dias, T.L.; Vieira, W.L. Students’ attitudes toward and knowledge about snakes in the semiarid region of Northeastern Brazil. J. Ethnobiol. Ethnomed. 2014, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Frynta, D.; Šimková, O.; Lišková, S.; Landová, E. Mammalian collection on Noah’s ark: The effects of beauty, brain and body size. PLoS ONE 2013, 8, e63110. [Google Scholar] [CrossRef] [PubMed]
- Landová, E.; Poláková, P.; Rádlová, S.; Janovcová, M.; Bobek, M.; Frynta, D. Beauty ranking of mammalian species kept in the Prague Zoo: Does beauty of animals increase the respondents’ willingness to protect them? Sci. Nat. 2018, 105, 69. [Google Scholar] [CrossRef] [PubMed]
- Lišková, S.; Frynta, D. What determines bird beauty in human eyes? Anthrozoös 2013, 26, 27–41. [Google Scholar] [CrossRef]
- Lišková, S.; Landová, E.; Frynta, D. Human preferences for colorful birds: Vivid colors or pattern? Evol. Psychol. 2015, 13, 147470491501300203. [Google Scholar]
- Frynta, D.; Lišková, S.; Bültmann, S.; Burda, H. Being attractive brings advantages: the case of parrot species in captivity. PLoS ONE 2010, 5, e12568. [Google Scholar] [CrossRef]
- Ekman, P.; Cordaro, D. What is meant by calling emotions basic. Emot. Rev. 2011, 3, 364–370. [Google Scholar] [CrossRef]
- Woody, S.R.; Teachman, B.A. Intersection of disgust and fear: Normative and pathological views. Clin. Psychol. Sci. Pract. 2000, 7, 291–311. [Google Scholar] [CrossRef]
- Oaten, M.; Stevenson, R.J.; Case, T.I. Disgust as a disease-avoidance mechanism. Psychol. Bull. 2009, 135, 303. [Google Scholar] [CrossRef]
- Curtis, V.; De Barra, M.; Aunger, R. Disgust as an adaptive system for disease avoidance behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Rozin, P.; Fallon, A.E. A perspective on disgust. Psychol. Rev. 1987, 94, 23. [Google Scholar] [CrossRef]
- Rozin, P.; Haidt, J.; Fincher, K. From oral to moral. Science 2009, 323, 1179–1180. [Google Scholar] [CrossRef]
- Cisler, J.M.; Olatunji, B.O.; Lohr, J.M. Disgust, fear, and the anxiety disorders: A critical review. Clin. Psychol. Rev. 2009, 29, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Marzillier, S.; Davey, G. Anxiety and disgust: Evidence for a unidirectional relationship. Cogn. Emot. 2005, 19, 729–750. [Google Scholar] [CrossRef]
- Davey, G.C. Disgust: The disease-avoidance emotion and its dysfunctions. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3453–3465. [Google Scholar] [CrossRef]
- Lindner, G. Crocodile management—Kakadu National Park. In Proceedings of the 17th Working Meeting of the IUCN-SSC Crocodile Specialist Group; The International Union for Conservation of Nature and Natural Resources (IUCN): Gland, Switzerland, 2004; pp. 41–51. [Google Scholar]
- Caldicott, D.G.; Croser, D.; Manolis, C.; Webb, G.; Britton, A. Crocodile attack in Australia: An analysis of its incidence and review of the pathology and management of crocodilian attacks in general. Wild. Environ. Med. 2005, 16, 143–159. [Google Scholar] [CrossRef]
- McGregor, J. Crocodile crimes: People versus wildlife and the politics of postcolonial conservation on Lake Kariba, Zimbabwe. Geoforum 2005, 36, 353–369. [Google Scholar] [CrossRef]
- The Snake Database. Available online: http://snakedatabase.org (accessed on 17 March 2019).
- Headland, T.N.; Greene, H.W. Hunter—Gatherers and other primates as prey, predators, and competitors of snakes. Proc. Natl. Acad. Sci. USA 2011, 108, E1470–E1474. [Google Scholar] [CrossRef]
- Murphy, J.C.; Henderson, R.W. Tales of Giant Snakes: A Historical Natural History of Anacondas and Pythons; Krieger Publishing Company: Malabar, FL, USA, 1997. [Google Scholar]
- Öhman, A.; Mineka, S. Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning. Psychol. Rev. 2001, 108, 483. [Google Scholar] [CrossRef]
- Isbell, L.A. The Fruit, the Tree, and the Serpent; Harvard University Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Öhman, A.; Mineka, S. The malicious serpent: Snakes as a prototypical stimulus for an evolved module of fear. Curr. Direct. Psychol. Sci. 2003, 12, 5–9. [Google Scholar] [CrossRef]
- Maddison, D.R. African origin of human mitochondrial DNA reexamined. Syst. Zool. 1991, 40, 355–363. [Google Scholar] [CrossRef]
- Ke, Y.; Su, B.; Song, X.; Lu, D.; Chen, L.; Li, H.; Xiao, C. African origin of modern humans in East Asia: A tale of 12,000 Y chromosomes. Science 2001, 292, 1151–1153. [Google Scholar] [CrossRef]
- Öhman, A.; Flykt, A.; Esteves, F. Emotion drives attention: detecting the snake in the grass. J. Exp. Psychol. Gen. 2001, 130, 466. [Google Scholar] [CrossRef] [PubMed]
- Flykt, A.; Caldara, R. Tracking fear in snake and spider fearful participants during visual search: A multi-response domain study. Cogn. Emot. 2006, 20, 1075–1091. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.C.; Lindström, B.; Esteves, F.; Öhman, A. The hidden snake in the grass: Superior detection of snakes in challenging attentional conditions. PLoS ONE 2014, 9, e114724. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.C.; Esteves, F.; Lundqvist, D.; Öhman, A. Some animal specific fears are more specific than others: Evidence from attention and emotion measures. Behav. Res. Ther. 2009, 47, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.C. The lurking snake in the grass: Interference of snake stimuli in visually taxing conditions. Evol. Psychol. 2012, 10. [Google Scholar] [CrossRef]
- Van Strien, J.W.; Eijlers, R.; Franken IH, A.; Huijding, J. Snake pictures draw more early attention than spider pictures in non-phobic women: Evidence from event-related brain potentials. Biol. Psychol. 2014, 96, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbell, L.A. Snakes as agents of evolutionary change in primate brains. J. Hum. Evol. 2006, 51, 1–35. [Google Scholar] [CrossRef] [Green Version]
- LoBue, V.; DeLoache, J.S. Detecting the snake in the grass: Attention to fear-relevant stimuli by adults and young children. Psychol. Sci. 2008, 19, 284–289. [Google Scholar] [CrossRef]
- Öhman, A.; Soares, S.C.; Juth, P.; Lindström, B.; Esteves, F. Evolutionary derived modulations of attention to two common fear stimuli: Serpents and hostile humans. J. Cogn. Psychol. 2012, 24, 17–32. [Google Scholar] [CrossRef]
- Van Le, Q.; Isbell, L.A.; Matsumoto, J.; Nguyen, M.; Hori, E.; Maior, R.S.; Nishijo, H. Pulvinar neurons reveal neurobiological evidence of past selection for rapid detection of snakes. Proc. Natl. Acad. Sci. USA 2013, 110, 19000–19005. [Google Scholar] [CrossRef] [Green Version]
- Baynes-Rock, M. Human perceptual and phobic biases for snakes: A review of the experimental evidence. Anthrozoös 2017, 30, 5–18. [Google Scholar] [CrossRef]
- Shibasaki, M.; Kawai, N. Rapid detection of snakes by Japanese monkeys (Macaca fuscata): An evolutionarily predisposed visual system. J. Comp. Psychol. 2009, 123, 131–135. [Google Scholar] [CrossRef]
- Kawai, N.; Koda, H. Japanese monkeys (Macaca fuscata) quickly detect snakes but not spiders: Evolutionary origins of fear-relevant animals. J. Comp. Psychol. 2016, 130, 299–303. [Google Scholar] [CrossRef]
- Bjerke, T.; Ødegårdstuen, T.S.; Kaltenborn, B.P. Attitudes toward animals among Norwegian children and adolescents: Species preferences. Anthrozoös 1998, 11, 227–235. [Google Scholar] [CrossRef]
- Prokop, P.; Tunnicliffe, S.D. Disgusting" Animals: Primary School Children’s Attitudes and Myths of Bats and Spiders. Euras. J. Math. Sci. Technol. Educ. 2008, 4. [Google Scholar] [CrossRef]
- Randler, C.; Hummel, E.; Prokop, P. Practical work at school reduces disgust and fear of unpopular animals. Soc. Anim. 2012, 20, 61–74. [Google Scholar] [CrossRef]
- Polák, J.; Rádlová, S.; Janovcová, M.; Flegr, J.; Landová, E.; Frynta, D. Scary and nasty beasts: A factor analysis and the role of fear and disgust in the perception of common phobic animals. Brit. J. Psychol. Under Review.
- Reese, A.M. Reptiles as food. Sci. Month. 1917, 5, 545–550. [Google Scholar]
- Klemens, M.W.; Thorbjarnarson, J.B. Reptiles as a food resource. Biodivers. Conserv. 1995, 4, 281–298. [Google Scholar] [CrossRef]
- Teachman, B.A.; Gregg, A.P.; Woody, S.R. Implicit associations for fear-relevant stimuli among individuals with snake and spider fears. J. Abnormal Psychol. 2001, 110, 226. [Google Scholar] [CrossRef]
- Wikström, J.; Lundh, L.G.; Westerlund, J.; Högman, L. Preattentive bias for snake words in snake phobia? Behav. Res. Ther. 2004, 42, 949–970. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R. Darwinian aesthetics informs traditional aesthetics. In Evolutionary Aesthetics; Voland, E., Grammar, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 9–35. [Google Scholar]
- Geldart, S.; Maurer, D.; Carney, K. Effects of eye size on adults’ aesthetic ratings of faces and 5-month-olds’ looking times. Perception 1999, 28, 361–374. [Google Scholar] [CrossRef]
- Richards, R. A new aesthetic for environmental awareness: Chaos theory, the beauty of nature, and our broader humanistic identity. J. Hum. Psychol. 2001, 41, 59–95. [Google Scholar] [CrossRef]
- Jacobsen, T.; Schubotz, R.I.; Höfel, L.; Cramon DY, V. Brain correlates of aesthetic judgment of beauty. Neuroimage 2006, 29, 276–285. [Google Scholar] [CrossRef]
- Woods, B. Beauty and the beast: Preferences for animals in Australia. J. Tour. Stud. 2000, 11, 25. [Google Scholar]
- Kühn, S.; Gallinat, J. The neural correlates of subjective pleasantness. Neuroimage 2012, 61, 289–294. [Google Scholar] [CrossRef]
- Frynta, D.; Marešová, J.; Řeháková-Petrů, M.; Šklíba, J.; Šumbera, R.; Krása, A. Cross-cultural agreement in perception of animal beauty: Boid snakes viewed by people from five continents. Hum. Ecol. 2011, 39, 829–834. [Google Scholar] [CrossRef]
- Landová, E.; Bakhshaliyeva, N.; Janovcová, M.; Peléšková, Š.; Suleymanova, M.; Polák, J.; Frynta, D. Association between fear and beauty evaluation of snakes: Cross-cultural findings. Front. Psychol. 2018, 9, 333. [Google Scholar] [CrossRef]
- Bitgood, S.; Patterson, D. Principles of exhibit design. Visit. Behav. 1987, 2, 4–6. [Google Scholar]
- Balmford, A. Separating fact from artifact in analyses of zoo visitor preferences. Conserv. Biol. 2000, 14, 1193–1195. [Google Scholar] [CrossRef]
- Sommer, R. Semantic profiles of zoos and their animals. Anthrozoös 2008, 21, 237–244. [Google Scholar] [CrossRef]
- Moss, A.; Esson, M. Visitor interest in zoo animals and the implications for collection planning and zoo education programmes. Zoo Biol. 2010, 29, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Czech, B.; Krausman, P.R.; Borkhataria, R. Social construction, political power, and the allocation of benefits to endangered species. Conserv. Biol. 1998, 12, 1103–1112. [Google Scholar] [CrossRef]
- Stokes, D.L. Things we like: Human preferences among similar organisms and implications for conservation. Hum. Ecol. 2007, 35, 361–369. [Google Scholar] [CrossRef]
- Martín-López, B.; Montes, C.; Benayas, J. The non-economic motives behind the willingness to pay for biodiversity conservation. Biol. Conserv. 2007, 139, 67–82. [Google Scholar] [CrossRef]
- Frynta, D.; Marešová, J.; Landová, E.; Lišková, S.; Šimková, O.; Tichá, I.; Fuchs, R. Are Animals in Zoos Rather Conspicuous than Endangered? Nova Science Publishers: New York, NY, USA, 2010. [Google Scholar]
- Marešová, J.; Frynta, D. Noah’s Ark is full of common species attractive to humans: The case of boid snakes in zoos. Ecol. Econ. 2008, 64, 554–558. [Google Scholar] [CrossRef]
- Dietz, J.M.; Dietz, L.A.; Nagagata, E.Y. The effective use of flagship species for conservation of biodiversity: The example of lion tamarins in Brazil. In Creative Conservation; Springer: Dordrecht, The Netherlands, 1994; pp. 32–49. [Google Scholar]
- Walpole, M.J.; Leader-Williams, N. Tourism and flagship species in conservation. Biodiv. Conserv. 2002, 11, 543–547. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef]
- Likert, R.A. A technique for the development of attitudes. Arch. Psychol. 1932, 140, 5–55. [Google Scholar]
- McGraw, K.O.; Wong, S.P. Forming inferences about some intraclass correlation coefficients. Psychol. Methods 1996, 1, 30. [Google Scholar] [CrossRef]
- Hallgren, K.A. Computing inter-rater reliability for observational data: An overview and tutorial. Tutor. Quant. Methods Psychol. 2002, 8, 23. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- StatSoft Inc. Statistica (Data Analysis Software System), Version 9.1. Available online: http://www.statsoft.com/ (accessed on 17 March 2019).
- Shrout, P.E.; Fleiss, J.L. Intraclass correlations: Uses in assessing rater reliability. Psychol. Bull. 1979, 86, 420. [Google Scholar] [CrossRef] [PubMed]
- Öhman, A. Of snakes and faces: An evolutionary perspective on the psychology of fear. Scand. J. Psychol. 2009, 50, 543–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A. Dangerous snakes, deadly snakes and medically important snakes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 26. [Google Scholar] [CrossRef]
- Ducey, S.D.; Cooper, J.S.; Wadman, M.C. Bitten by a Dragon. Wild. Environ. Med. 2016, 27, 291–293. [Google Scholar] [CrossRef]
- Pittenger, J.B. Body proportions as information for age and cuteness: Animals in illustrated children’s books. Attent. Perc. Psychophys. 1990, 48, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Masataka, N.; Hayakawa, S.; Kawai, N. Human young children as well as adults demonstrate superior rapid snake detection when typical striking posture is displayed by the snake. PLoS ONE 2010, 5, e15122. [Google Scholar] [CrossRef]
- Bar, M.; Neta, M. Humans prefer curved visual objects. Psychol. Sci. 2006, 17, 645–648. [Google Scholar] [CrossRef]
- Bar, M.; Neta, M. Visual elements of subjective preference modulate amygdala activation. Neuropsychologia 2007, 45, 2191–2200. [Google Scholar] [CrossRef] [Green Version]
- Wegener, J.E.; Swoboda, S.; Hawlitschek, O.; Franzen, M.; Wallach, V.; Vences, M.; Glaw, F. Morphological variation and taxonomic reassessment of the endemic Malagasy blind snake family Xenotyphlopidae. Spixiana 2013, 36, 269–282. [Google Scholar]
- Prokop, P.; Fančovičová, J. The association between disgust, danger and fear of macroparasites and human behaviour. Acta Ethol. 2010, 13, 57–62. [Google Scholar] [CrossRef]
- Kuplen, M. Disgust and ugliness: A kantian perspective. Cont. Aesthet. 2011, 9, 10. [Google Scholar]
- Korsmeyer, C. Disgust and Aesthetics. Philos. Compass 2012, 7, 753–761. [Google Scholar] [CrossRef]
- Rádlová, S.; Janovcová, M.; Polák, J.; Landová, E.; Frynta, D. Emoce vyvolané zvířaty I: krásaa estetické preference (Emotion awared animals I: beauty and aesthetic preference). E-Psychologie 2018, 12, 35–50. [Google Scholar] [CrossRef]
- Landová, E.; Marešová, J.; Šimková, O.; Cikánová, V.; Frynta, D. Human responses to live snakes and their photographs: Evaluation of beauty and fear of the king snakes. J. Environ. Psychol. 2012, 32, 69–77. [Google Scholar] [CrossRef]
- Davey, G.C. Self-reported fears to common indigenous animals in an adult UK population: The role of disgust sensitivity. Brit. J. Psychol. 1994, 85, 541–554. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eigenvalue | Proportion Explained by | |||||||
|---|---|---|---|---|---|---|---|---|
| Stimuli Subsets | No. of Axes | Eigenvalues > 1 | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 |
| Set1: fear | 121 | 27 | 267.18 | 26.73 | 10.40 | 69.68 % | 6.97 % | 2.71 % |
| Set1: disgust | 125 | 27 | 192.71 | 34.22 | 9.63 | 61.28 % | 10.88 % | 3.06 % |
| Set1: beauty | 126 | 49 | 182.98 | 27.70 | 15.93 | 47.86 % | 7.25 % | 4.17 % |
| Set1: pooled | 126 | 87 | 548.17 | 156.82 | 43.12 | 50.74 % | 14.52 % | 3.99 % |
| Set2: fear | 126 | 38 | 388.81 | 33.04 | 13.01 | 70.94 % | 6.03 % | 2.37 % |
| Set2: disgust | 126 | 33 | 229.10 | 41.94 | 10.32 | 61.19 % | 11.20 % | 2.76 % |
| Set2: beauty | 126 | 56 | 188.36 | 44.20 | 19.55 | 42.53 % | 9.98 % | 4.41 % |
| Set2: pooled | 126 | 103 | 696.29 | 193.68 | 52.10 | 51.00 % | 14.18 % | 3.82 % |
| (a) Set 1 | mean disgust | mean fear | mean beauty | PC1 fear | PC2 fear | PC1 disgust | PC2 disgust | PC1 beauty | PC2 beauty |
| mean disgust | 1 | 0.773 | −0.836 | 0.786 | 0.369 | 0.995 | 0.002 | 0.859 | −0.423 |
| mean fear | 0.773 | 1 | −0.458 | 0.995 | −0.136 | 0.796 | −0.529 | 0.494 | −0.682 |
| mean beauty | −0.836 | −0.458 | 1 | −0.468 | −0.471 | −0.807 | −0.333 | −0.998 | 0.030 |
| PC1 fear | 0.786 | 0.995 | −0.468 | 1 | −0.104 | 0.808 | −0.537 | 0.505 | −0.711 |
| PC2 fear | 0.369 | −0.136 | −0.471 | −0.104 | 1 | 0.354 | 0.408 | 0.475 | −0.031 |
| PC1 disgust | 0.995 | 0.796 | −0.807 | 0.808 | 0.354 | 1 | −0.043 | 0.831 | −0.454 |
| PC2 disgust | 0.002 | −0.529 | −0.333 | −0.537 | 0.408 | −0.043 | 1 | 0.296 | 0.719 |
| PC1 beauty | 0.859 | 0.494 | −0.998 | 0.505 | 0.475 | 0.831 | 0.296 | 1 | −0.082 |
| PC2 beauty | −0.423 | −0.682 | 0.030 | −0.711 | −0.031 | −0.454 | 0.719 | −0.082 | 1 |
| (b) Set 2 | mean disgust | mean fear | mean beauty | PC1 fear | PC2 fear | PC1 disgust | PC2 disgust | PC1 beauty | PC2 beauty |
| mean disgust | 1 | 0.774 | −0.797 | −0.791 | 0.419 | −0.988 | 0.154 | 0.853 | 0.423 |
| mean fear | 0.774 | 1 | −0.421 | −0.996 | −0.055 | −0.800 | −0.346 | 0.504 | 0.724 |
| mean beauty | −0.797 | −0.421 | 1 | 0.428 | −0.416 | 0.748 | −0.451 | −0.989 | 0.041 |
| PC1 fear | −0.791 | −0.996 | 0.428 | 1 | 0.025 | 0.817 | 0.353 | −0.514 | −0.744 |
| PC2 fear | 0.419 | −0.055 | −0.416 | 0.025 | 1 | −0.421 | 0.402 | 0.425 | −0.064 |
| PC1 disgust | −0.988 | −0.800 | 0.748 | 0.817 | −0.421 | 1 | −0.095 | −0.811 | −0.476 |
| PC2 disgust | 0.154 | −0.346 | −0.451 | 0.353 | 0.402 | −0.095 | 1 | 0.374 | −0.695 |
| PC1 beauty | 0.853 | 0.504 | −0.989 | −0.514 | 0.425 | −0.811 | 0.374 | 1 | 0.071 |
| PC2 beauty | 0.423 | 0.724 | 0.041 | −0.744 | −0.064 | −0.476 | −0.695 | 0.071 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janovcová, M.; Rádlová, S.; Polák, J.; Sedláčková, K.; Peléšková, Š.; Žampachová, B.; Frynta, D.; Landová, E. Human Attitude toward Reptiles: A Relationship between Fear, Disgust, and Aesthetic Preferences. Animals 2019, 9, 238. https://doi.org/10.3390/ani9050238
Janovcová M, Rádlová S, Polák J, Sedláčková K, Peléšková Š, Žampachová B, Frynta D, Landová E. Human Attitude toward Reptiles: A Relationship between Fear, Disgust, and Aesthetic Preferences. Animals. 2019; 9(5):238. https://doi.org/10.3390/ani9050238
Chicago/Turabian StyleJanovcová, Markéta, Silvie Rádlová, Jakub Polák, Kristýna Sedláčková, Šárka Peléšková, Barbora Žampachová, Daniel Frynta, and Eva Landová. 2019. "Human Attitude toward Reptiles: A Relationship between Fear, Disgust, and Aesthetic Preferences" Animals 9, no. 5: 238. https://doi.org/10.3390/ani9050238