Early Feed Restriction Programs Metabolic Disorders in Fattening Merino Lambs

 ,
,  , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Liver and Ruminal Parameters
2.2. Blood Sampling, Biochemical Profile Analysis, and Flow Cytometry
2.3. Statistical Analysis
3. Results
3.1. Biochemical Profile
3.2. Immunological Parameters
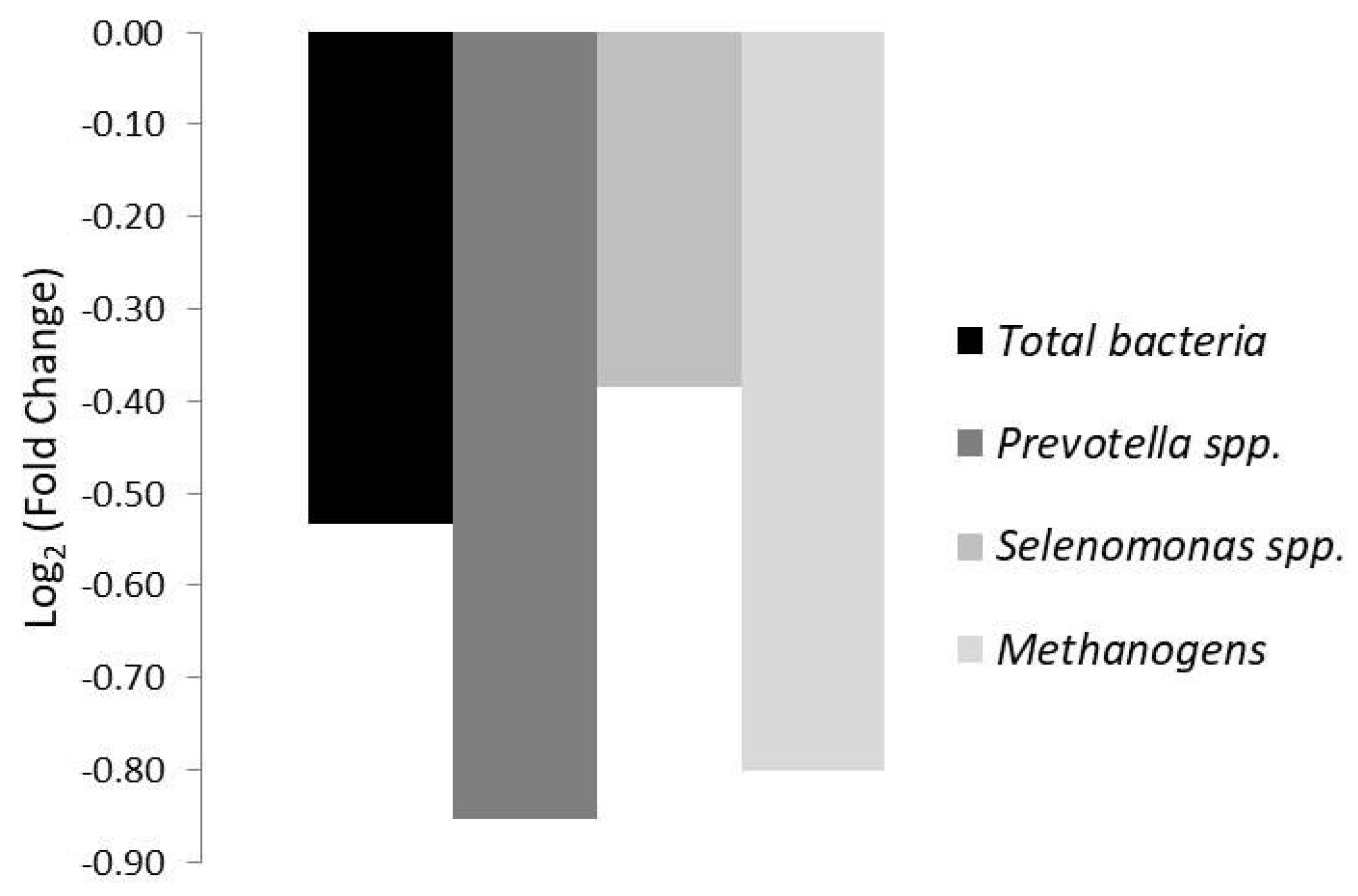
3.3. Liver Antioxidant Status (TBARS), Ruminal Characteristics, and Epimural Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.P.; Martyn, C.N.; Osmond, C.; Hales, C.N.; Fall, C.H.D. Growth in-utero and serum-cholesterol concentrations in adult life. BMJ 1993, 307, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Lizárraga-Mollinedo, E.; Fernández-Millán, E.; García-San Frutos, M.; de Toro-Martín, J.; Fernández-Agulló, T.; Ros, M.; Álvarez, C.; Escrivá, F. Early and long-term undernutrition in female rats exacerbates the metabolic risk associated with nutritional rehabilitation. J. Biol. Chem. 2015, 290, 19353–19366. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.S.; Tingey, K.; Van Bon, B.W.M.; Ozanne, S.E.; Wilson, V.; Dandrea, J.; Keisler, D.H.; Stephenson, T.; Symonds, M.E. Programming of glucose-insulin metabolism in adult sheep after maternal undernutrition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R947–R954. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Li, M.; Fries, P.; Griebel, P.J.; Guan, L.L. Regional and age dependent changes in gene expression of toll-like receptors and key antimicrobial defence molecules throughout the gastrointestinal tract of dairy calves. Vet. Immunol. Immunopathol. 2012, 146, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, E.; Amadori, M.; Riva, F.; Bertoni, G.; Bani, P. Evaluation of innate immune responses in bovine forestomachs. Res. Vet. Sci. 2014, 96, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Changes in the rumen epimural bacterial diversity of beef cattle as affected by diet and induced ruminal acidosis. Appl. Environ. Microbiol. 2013, 79, 3744–3755. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef] [PubMed]
- Meale, S.J.; Li, S.; Azevedo, P.; Derakhshani, H.; Plaizier, J.C.; Khafipour, E.; Steele, M.A. Development of ruminal and fecal microbiomes are affected by weaning but not weaning strategy in dairy calves. Front. Microbiol. 2016, 7, 582. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Giráldez, F.J.; Mateo, J.; Frutos, J.; Andrés, S. Programming merino lambs by early feed restriction reduces growth rates and increases fat accretion during the fattening period with no effect on meat quality traits. Meat Sci. 2018, 135, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Frutos, J.; Andrés, S.; Yáñez-Ruiz, D.R.; Benavides, J.; López, S.; Santos, A.; Martínez-Valladares, M.; Rozada, F.; Giráldez, F.J. Early feed restriction of lambs modifies ileal epimural microbiota and affects immunity parameters during the fattening period. Animal 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Giráldez, F.J.; Trevisi, E.; Lucini, L.; Frutos, J.; Andrés, S. Liver transcriptomic and plasma metabolomic profiles of fattening lambs are modified by feed restriction during the suckling period. J. Anim. Sci. 2018, 96, 1495–1507. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Valdés, C.; Giráldez, F.J.; López, S.; France, J.; Frutos, J.; Fernández, M.; Andrés, S. Feed efficiency and the liver proteome of fattening lambs are modified by feed restriction during the suckling period. Animal 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bodas, R.; Prieto, N.; Jordán, M.J.; López-Campos, O.; Giráldez, F.J.; Morán, L.; Andrés, S. The liver antioxidant status of fattening lambs is improved by naringin dietary supplementation at 0.15% rates but not meat quality. Animal 2012, 6, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Andrés, S.; Bodas, R.; Tejido, M.L.; Giráldez, F.J.; Valdés, C.; López, S. Effects of the inclusion of flaxseed and quercetin in the diet of fattening lambs on ruminal microbiota, in vitro fermentation and biohydrogenation of fatty acids. J. Agric. Sci. 2016, 154, 542–552. [Google Scholar] [CrossRef]
- Vargas, J.E.; Andrés, S.; Snelling, T.J.; López-Ferreras, L.; Yáñez-Ruiz, D.R.; García-Estrada, C.; López, S. Effect of sunflower and marine oils on ruminal microbiota, in vitro fermentation and digesta fatty acid profile. Front. Microbiol. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Sebastiano, S.R.; Sweeney, T.; Hanrahan, J.P.; Glynn, A.; Keane, O.M.; Mukhopadhya, A.; Thornton, K.; Good, B. Breed differences in humoral and cellular responses of lambs to experimental infection with the gastrointestinal nematode teladorsagia circumcincta. Vet. Res. 2015, 46, 8. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rodríguez, J.; Monleón, E.; Sanz, A.; Badiola, J.J.; Joy, M. Rumen fermentation and histology in light lambs as affected by forage supply and lactation length. Res. Vet. Sci. 2012, 92, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Jacometo, C.B.; Osorio, J.S.; Socha, M.; Correa, M.N.; Piccioli-Cappelli, F.; Trevisi, E.; Loor, J.J. Maternal consumption of organic trace minerals alters calf systemic and neutrophil mRNA and microRNA indicators of inflammation and oxidative stress. J. Dairy Sci. 2015, 98, 7717–7729. [Google Scholar] [CrossRef] [PubMed]
- Morán, L.; Andrés, S.; Bodas, R.; Benavides, J.; Prieto, N.; Pérez, V.; Giráldez, F.J. Antioxidants included in the diet of fattening lambs: Effects on immune response, stress, welfare and distal gut microbiota. Anim. Feed Sci. Technol. 2012, 173, 177–185. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Hunt, A.S.; Slepetis, R.M.; Finnerty, K.D.; Alston, C.; Beermann, D.H.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep: III. Regulation of energy metabolism. J. Anim. Sci. 2002, 80, 2850–2861. [Google Scholar] [CrossRef] [PubMed]
- Taschuk, R.; Griebel, P.J. Commensal microbiome effects on mucosal immune system development in the ruminant gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic stress and inflammatory response in high-yielding, periparturient dairy cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Viñoles, C.; De Barbieri, I.; Gil, J.; Olivera, J.; Fierro, S.; Bialade, F.; Montossi, F. Long-term effect of nutrition on the metabolic status and reproductive potential of merino rams under grazing conditions. Anim. Prod. Sci. 2012, 52, 881–889. [Google Scholar] [CrossRef]
- Nurliyana, A.R.; Shariff, Z.M.; Taib, M.N.M.; Gan, W.Y.; Tan, K.A. Early nutrition, growth and cognitive development of infants from birth to 2 years in malaysia: A study protocol. BMC Pediatr. 2016, 16, 160. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dong, H.; Zhang, Y.; Cao, M.; Song, L.; Pan, Q.; Bulmer, A.; Adams, D.B.; Dong, X.; Wang, H. Bilirubin increases insulin sensitivity by regulating cholesterol metabolism, adipokines and PPARγ levels. Sci. Rep. 2015, 5, 9886. [Google Scholar] [CrossRef] [PubMed]
- Jansen, T.; Daiber, A. Direct antioxidant properties of bilirubin and biliverdin. Is there a role for biliverdin reductase? Front. Pharmacol. 2012, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.A.; AlZahal, O.; Hook, S.E.; Croom, J.; McBride, B.W. Ruminal acidosis and the rapid onset of ruminal parakeratosis in a mature dairy cow: A case report. Acta Vet. Scand. 2009, 51, 39. [Google Scholar] [CrossRef] [PubMed]
- Khiaosa-ard, R.; Pourazad, P.; Aditya, S.; Humer, E.; Zebeli, Q. Factors related to variation in the susceptibility to subacute ruminal acidosis in early lactating simmental cows fed the same grain-rich diet. Anim. Feed Sci. Technol. 2018, 238, 111–122. [Google Scholar] [CrossRef]


{kind=link}
| Treatment | Growth Stage | SED1 | SED2 | p-Values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ADL | RES | SCK | PWN | FTT | T | GS | T*GS | |||
| Haptoglobin (g/L) | 0.22 | 0.23 | 0.21 A | 0.20 A | 0.27 B | 0.022 | 0.025 | 0.886 | 0.016 | 0.470 |
| Ceruloplasmin (µmol/L) | 2.34 A | 3.10 B | 2.81 | 2.80 | 2.55 | 0.262 | 0.222 | 0.009 | 0.427 | 0.308 |
| β-hydroxybutyrate (mmol/L) | 0.29 | 0.31 | 0.30 | 0.27 | 0.32 | 0.027 | 0.033 | 0.495 | 0.393 | 0.316 |
| Paraoxonase (U/L) | 117 | 125 | 88 A | 150 C | 125 B | 15.0 | 7.63 | 0.624 | <0.001 | 0.018 |
| ROM (mg/100 mL) | 16.5 | 17.6 | 17.0 | 17.9 | 16.2 | 1.22 | 1.04 | 0.382 | 0.276 | 0.020 |
| Myeloperoxidase (U/L) | 434 | 466 | 400 A | 417 A | 533 B | 52.1 | 49.8 | 0.554 | 0.021 | 0.848 |
| FRAP (µmol/L) | 154 | 154 | 147 | 152 | 164 | 10.3 | 12.6 | 0.945 | 0.400 | 0.207 |
| SOD (U/L) | 0.69 A | 1.04 B | 0.64 A | 0.89 B | 1.07 B | 0.077 | 0.094 | <0.001 | <0.001 | 0.043 |
| Ca (mmol/L) | 3.02 | 2.96 | 3.16 B | 3.09 B | 2.73 A | 0.044 | 0.043 | 0.174 | <0.001 | 0.200 |
| Mg (mmol/L) | 0.92 | 0.90 | 0.86 A | 0.87 A | 0.99 B | 0.022 | 0.016 | 0.557 | <0.001 | 0.461 |
| Zn (mmol/L) | 15.8 | 16.5 | 18.9 C | 16.0 B | 13.6 A | 0.959 | 1.174 | 0.481 | <0.001 | 0.196 |
| Retinol (µg/100 mL) | 40.2 | 39.9 | 3.43 | 0.935 | ||||||
| Tocopherol (µg/mL) | 0.23 | 0.23 | 0.016 | 0.867 | ||||||
| AST (U/L) | 146 | 162 | 73 A | 87 A | 301 B | 36.13 | 44.25 | 0.656 | <0.001 | 0.763 |
| GGT (U/L) | 126 | 102 | 210 B | 69 A | 63 A | 22.91 | 27.12 | 0.298 | <0.001 | 0.271 |
| Total bilirubin (µmol/L) | 2.10 B | 0.93 A | 2.73 B | 1.06 A | 0.76 A | 0.314 | 0.3770 | <0.001 | <0.001 | 0.661 |
| Creatinine (µmol/L) | 63.5 | 63.7 | 57.5 A | 65.2 B | 68.1 C | 1.73 | 1.113 | 0.937 | <0.001 | 0.385 |
| Glucose (mmol/L) | 6.66 | 6.31 | 7.47 B | 6.76 B | 5.22 A | 0.212 | 0.164 | 0.109 | <0.001 | 0.675 |
| Insulin (ng/L) | 497 B | 289 A | 418 B | 499 B | 262 A | 92.2 | 106.8 | 0.034 | 0.008 | 0.025 |
| Urea (mmol/L) | 6.89 | 7.45 | 4.59 A | 6.09 B | 10.84 C | 0.37 | 0.36 | 0.143 | <0.001 | 0.009 |
| Protein (g/L) | 63.3 | 63.3 | 62.9 | 63.3 | 63.7 | 1.29 | 1.21 | 0.978 | 0.816 | 0.423 |
| Albumin (g/L) | 34.8 | 34.7 | 33.4 A | 35.5 B | 35.3 B | 0.41 | 0.42 | 0.947 | <0.001 | 0.706 |
| Globulin (g/L) | 28.6 | 28.6 | 29.5 | 27.8 | 28.4 | 1.23 | 1.37 | 0.995 | 0.440 | 0.420 |
| Cholesterol (mmol/L) | 2.06 A | 2.50 B | 3.08 B | 2.72 B | 1.05 A | 0.173 | 0.193 | 0.020 | <0.001 | 0.497 |
| LDL (mmol/L) | 0.56 A | 0.72 B | 0.88 C | 0.73 B | 0.32 A | 0.076 | 0.750 | 0.046 | <0.001 | 0.746 |
| HDL (mmol/L) | 1.10 A | 1.33 B | 1.67 B | 1.50 B | 0.48 A | 0.103 | 0.107 | 0.039 | <0.001 | 0.392 |
| NEFA (g/L) | 0.43 | 0.44 | 0.63 C | 0.53 B | 0.14 A | 0.042 | 0.44 | 0.755 | <0.001 | 0.639 |
| Triglycerides (mmol/L) | 0.53 | 0.54 | 0.75 B | 0.68 B | 0.18 A | 0.060 | 0.072 | 0.872 | <0.001 | 0.977 |
| ADL | RES | RSD | p-Value | |
|---|---|---|---|---|
| Lymphocytes (%) 1 | ||||
| CD4+ | 9.29 | 12.2 | 7.4 | 0.351 |
| CD8+ | 4.83 | 5.26 | 3.69 | 0.779 |
| CD45+ | 17.5 | 19.5 | 10.4 | 0.649 |
| CD21+ | 21.8 | 21.9 | 14.0 | 0.719 |
| CD4+/CD8+ | 1.79 | 2.32 | 1.51 | 0.403 |
| Ruminal parameters | ||||
| Mean gray value 2 | 130 | 126 | 8.5 | 0.297 |
| Papillae length AV (µm) | 3985 | 3649 | 856 | 0.359 |
| Papillae length PD (µm) | 3878 | 3874 | 893 | 0.993 |
| Papillae width AV (µm) | 1244 | 1178 | 177 | 0.383 |
| Papillae width PD (µm) | 1289 | 1318 | 220 | 0.771 |
| Stratum corneum thickness AV (µm) | 61.3 | 59.1 | 13.6 | 0.690 |
| Stratum corneum thickness PD (µm) | 60.2 | 70.6 | 19.8 | 0.208 |
| IgA | ||||
| Blood (pg/mL) | 7921 | 8981 | 2595 | 0.373 |
| Rumen (pg/µg total protein) | 5.87 | 4.51 | 3.52 | 0.413 |
| ADL | RES | RSD | p-Value | |
|---|---|---|---|---|
| Cytokines (ΔCq) 1 | ||||
| IFN-γ | 11.3 | 11.2 | 1.6 | 0.927 |
| TGF-β | 6.32 | 6.52 | 0.65 | 0.558 |
| TLR (ΔCq) 1 | ||||
| TLR1 | 11.3 | 11.9 | 0.8 | 0.072 |
| TLR2 | 23.6 | 23.5 | 1.9 | 0.903 |
| TLR3 | 14.8 | 15.4 | 0.8 | 0.056 |
| TLR4 | 12.2 | 12.4 | 0.7 | 0.521 |
| TLR5 | 23.7 | 23.6 | 1.7 | 0.870 |
| TLR6 | 11.7 | 12.1 | 1.0 | 0.364 |
| TLR7 | 15.2 | 15.7 | 0.9 | 0.212 |
| TLR8 | 18.0 | 17.8 | 0.8 | 0.577 |
| TLR9 | 16.9 | 17.0 | 1.4 | 0.957 |
| TLR10 | 14.0 | 13.9 | 1.3 | 0.938 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frutos, J.; Andrés, S.; Trevisi, E.; Yáñez-Ruiz, D.R.; López, S.; Santos, A.; Giráldez, F.J. Early Feed Restriction Programs Metabolic Disorders in Fattening Merino Lambs. Animals 2018, 8, 83. https://doi.org/10.3390/ani8060083
Frutos J, Andrés S, Trevisi E, Yáñez-Ruiz DR, López S, Santos A, Giráldez FJ. Early Feed Restriction Programs Metabolic Disorders in Fattening Merino Lambs. Animals. 2018; 8(6):83. https://doi.org/10.3390/ani8060083
Chicago/Turabian StyleFrutos, Javier, Sonia Andrés, Erminio Trevisi, David R. Yáñez-Ruiz, Secundino López, Alba Santos, and F. Javier Giráldez. 2018. "Early Feed Restriction Programs Metabolic Disorders in Fattening Merino Lambs" Animals 8, no. 6: 83. https://doi.org/10.3390/ani8060083
APA StyleFrutos, J., Andrés, S., Trevisi, E., Yáñez-Ruiz, D. R., López, S., Santos, A., & Giráldez, F. J. (2018). Early Feed Restriction Programs Metabolic Disorders in Fattening Merino Lambs. Animals, 8(6), 83. https://doi.org/10.3390/ani8060083