
Effect of Freezing Conditions on Fecal Bacterial Composition in Pigs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals and Fecal Sample Processing
2.2. Genomic DNA Isolation
2.3. Quantitative PCR
2.4. Statistical Analyses
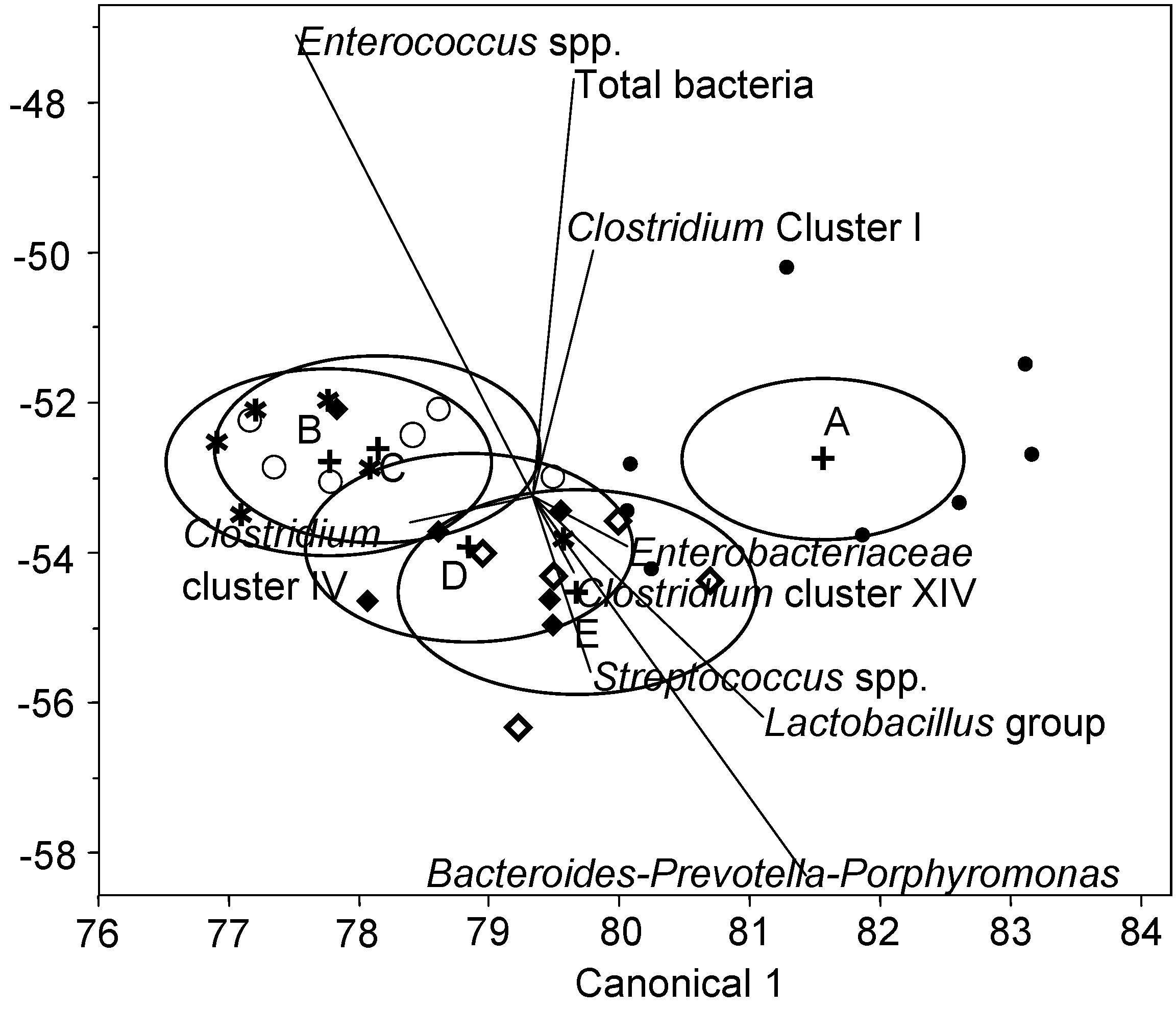
3. Results and Discussion
3.1. DNA Yield and Total Bacterial 16S rRNA GENE Abundance
3.2. Bacterial Composition
4. Conclusions
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Young, V.B.; Kahn, S.A.; Schmidt, T.M.; Chang, E.B. Studying the enteric microbiome in inflammatory bowel diseases: Getting through the Growing Pains and Moving Forward. Front. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Isaacsen, R.E. The pig gut microbial diversity: Understanding the pig gut microbial ecology through the next Generation High Throughput Sequencing. Vet. Microbiol. 2015, 177, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Schokker, D.; Veninga, G.; Vastenhouw, S.A.; Bossers, A.; de Bree, F.M.; Kaal-Lansbergen, L.M.; Rebel, J.M.; Smits, M.A. Early life microbial colonization of the gut and intestinal development differ between genetically divergent broiler lines. BMC Genomics. 2015. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Mann, E.; Schmitz-Esser, S.; Wagner, M.; Ritzmann, M.; Zebeli, Q. Changing dietary calcium-phosphorus level and cereal source selectively alters abundance of bacteria and metabolites in the upper gastrointestinal tracts of weaned pigs. Appl. Environ. Microbiol. 2013, 79, 7264–7272. [Google Scholar] [CrossRef] [PubMed]
- Jahanbakhsh, S.; Kabore, K.P.; Fravalo, P.; Letellier, A.; Fairbrother, J.M. Impact of medicated feed along with clay mineral supplementation on Escherichia coli resistance to antimicrobial agents in pigs after weaning in field conditions. Res. Vet. Sci. 2015, 102, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Stensland, I.; Kim, J.C.; Bowring, B.; Collins, A.M.; Mansfield, J.P.; Pluske, J.R. A comparison of diets supplemented with a feed additive containing organic acids, cinnamaldehyde and a permeabilizing complex, or zinc oxide, on post-weaning diarrhoea, selected bacterial populations, blood measures and performance in weaned pigs experimentally infected with enterotoxigenic E. coli. Animals 2015, 5, 1147–1168. [Google Scholar] [PubMed]
- Bahl, M.I.; Bergström, A.; Licht, T.R. Freezing fecal samples prior to DNA extraction affects the Firmicutes to Bacteroidetes ratio determined by downstream quantitative PCR analysis. FEMS Microbiol. Lett. 2012, 329, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Maukonen, J.; Simões, C.; Saarela, M. The currently used commercial DNA-extraction methods give different results of clostridial and actinobacterial populations derived from human fecal samples. FEMS Microbiol. Ecol. 2012, 79, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Kittelmann, S.; Miri, V.H.; Zethof, M.; Noel, S.J.; Waghorn, G.C.; Janssen, P.H. Effect of DNA extraction methods and sampling techniques on the apparent structure of cow and sheep rumen microbial communities. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Donovan, S.M. Human microbiota-associated swine: Current Progress and Future Opportunities. ILAR J. 2015, 56, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Schmitz-Esser, S.; Mann, E.; Grüll, D.; Molnar, T.; Zebeli, Q. Adaptation of the cecal bacterial microbiome of growing pigs in response to resistant starch type 4. Appl. Environ. Microbiol. 2015, 81, 8489–8499. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Hooda, S.; Pieper, R.; Zijlstra, R.T.; van Kessel, A.G.; Mosenthin, R.; Gänzle, M.G. Nonstarch polysaccharides modulate bacterial microbiota, pathways for butyrate production, and abundance of pathogenic Escherichia coli in the pig gastrointestinal tract. Appl. Environ. Microbiol. 2010, 76, 3692–3701. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. J. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Cardona, S.; Eck, A.; Cassellas, M.; Gallart, M.; Alastrue, C.; Dore, J.; Azpiroz, F.; Roca, J.; Guarner, F.; Manichanh, C. Storage conditions of intestinal microbiota matter in metagenomic analysis. BMC Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouhy, F.; Deane, J.; Rea, M.C.; O’Sullivan, Ó.; Ross, R.P.; O’Callaghan, G.; Plant, B.J.; Stanton, C. The effects of freezing on faecal microbiota as determined using MiSeq sequencing and culture-based investigations. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, I.M.; Ringel-Kulka, T.; Siddle, J.P.; Klaenhammer, T.R.; Ringel, Y. Charaterization of the fecal mcirobiota using high-throughput sequencing reveals a stable microbial community during storage. PLoS ONE 2012. [Google Scholar] [CrossRef]
- Mullins, C.R.; Mamedova, L.K.; Carpenter, A.J.; Ying, Y.; Allen, M.S.; Yoon, I.; Bradford, B.J. Analysis of rumen microbial populations in lactating dairy cattle fed diets varying in carbohydrate profiles and Saccharomyces cerevisiae fermentation product. J. Dairy Sci. 2013, 96, 5872–5881. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Snap-Freezing in Liquid N2 | No | No | Yes | No | Yes | SEM | Contrasts (p-Value) | ||
|---|---|---|---|---|---|---|---|---|---|
| Storage Temperature | No | −20 °C | −20 °C | −80 °C | −80 °C | DNA Extraction from Fresh vs. Frozen Feces | Storage at −20 °C vs. −80 °C | Snap-Freezing vs. None Prior to Freeze-Storage | |
| Log10 Gene Copies/G feces | |||||||||
| Total Bacteria | 10.8 | 10.1 | 10.1 | 10.2 | 10.2 | 0.09 | <0.001 | 0.176 | 0.800 |
| Lactobacillus group | 7.6 | 7.4 | 7.4 | 7.5 | 7.4 | 0.35 | 0.021 | 0.477 | 0.892 |
| Enterococcus spp. | 5.7 | 5.5 | 5.6 | 5.6 | 5.6 | 0.34 | 0.018 | 0.718 | 0.771 |
| Streptococcus spp. | 7.9 | 7.6 | 7.6 | 7.7 | 7.7 | 0.28 | 0.035 | 0.454 | 0.685 |
| Clostridium cluster XIV | 9.3 | 8.9 | 9.0 | 9.0 | 9.1 | 0.07 | <0.001 | 0.075 | 0.257 |
| Clostridium cluster IV | 8.7 | 8.24 | 8.25 | 8.42 | 8.27 | 0.12 | <0.001 | 0.303 | 0.461 |
| Clostridium cluster I | 8.0 | 7.7 | 7.8 | 7.8 | 7.7 | 0.34 | 0.022 | 0.750 | 0.684 |
| Bacteroides-Prevotella-Porphyromonas | 10.2 | 9.7 | 9.7 | 9.8 | 9.8 | 0.08 | <0.001 | 0.033 | 0.665 |
| Enterobacteriaceae | 7.1 | 6.7 | 6.9 | 7.0 | 6.9 | 0.60 | 0.011 | 0.060 | 0.516 |
| Log10 gene copies/ng DNA | |||||||||
| Total Bacteria | 6.5 | 6.3 | 6.3 | 6.3 | 6.3 | 0.04 | <0.001 | 0.666 | 0.885 |
| Lactobacillus Group | 3.4 | 3.6 | 3.6 | 3.6 | 3.6 | 0.34 | <0.001 | 0.104 | 0.945 |
| Enterococcus spp. | 1.4 | 1.7 | 1.7 | 1.6 | 1.6 | 0.34 | <0.001 | 0.032 | 0.907 |
| Streptococcus spp. | 3.6 | 3.8 | 3.8 | 3.7 | 3.7 | 0.29 | 0.004 | 0.241 | 0.798 |
| Clostridium Cluster XIV | 5.1 | 5.2 | 5.2 | 5.1 | 5.2 | 0.05 | 0.276 | 0.583 | 0.429 |
| Clostridium Cluster IV | 4.4 | 4.4 | 4.4 | 4.4 | 4.3 | 0.09 | 0.693 | 0.532 | 0.103 |
| Clostridium cluster I | 3.7 | 3.9 | 3.9 | 3.8 | 3.8 | 0.35 | 0.032 | 0.119 | 0.827 |
| Bacteroides-Prevotella-Porphyromonas | 5.9 | 6.0 | 5.9 | 5.9 | 5.9 | 0.07 | 0.636 | 0.616 | 0.671 |
| Enterobacteriaceae | 2.9 | 3.0 | 3.1 | 3.1 | 3.1 | 0.59 | 0.013 | 0.749 | 0.532 |
| Relative Abundance of Bacterial Groups Expressed as Proportion of Total Bacteria (%) | |||||||||
| Lactobacillus Group | 0.61 | 0.96 | 0.92 | 0.91 | 1.02 | 0.265 | <0.011 | 0.850 | 0.740 |
| Enterococcus spp. | 0.007 | 0.014 | 0.013 | 0.013 | 0.011 | 0.006 | 0.041 | 0.668 | 0.449 |
| Streptococcus spp. | 0.34 | 0.73 | 0.74 | 0.68 | 0.70 | 0.244 | <0.001 | 0.603 | 0.869 |
| Clostridium Cluster XIV | 6.26 | 7.35 | 7.97 | 7.40 | 8.54 | 1.239 | 0.236 | 0.794 | 0.460 |
| Clostridium Cluster IV | 1.09 | 1.66 | 1.53 | 1.81 | 1.22 | 0.266 | 0.007 | 0.595 | 0.031 |
| Clostridium Cluster I | 1.54 | 1.69 | 1.61 | 1.74 | 1.60 | 0.894 | 0.461 | 0.903 | 0.500 |
| Bacteroides-Prevotella-Porphyromonas | 25.77 | 41.72 | 42.74 | 42.41 | 42.59 | 7.059 | <0.001 | 0.931 | 0.848 |
| Enterobacteriaceae | 0.11 | 0.26 | 0.23 | 0.21 | 0.18 | 0.179 | 0.069 | 0.335 | 0.611 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metzler-Zebeli, B.U.; Lawlor, P.G.; Magowan, E.; Zebeli, Q. Effect of Freezing Conditions on Fecal Bacterial Composition in Pigs. Animals 2016, 6, 18. https://doi.org/10.3390/ani6030018
Metzler-Zebeli BU, Lawlor PG, Magowan E, Zebeli Q. Effect of Freezing Conditions on Fecal Bacterial Composition in Pigs. Animals. 2016; 6(3):18. https://doi.org/10.3390/ani6030018
Chicago/Turabian StyleMetzler-Zebeli, Barbara U., Peadar G. Lawlor, Elizabeth Magowan, and Qendrim Zebeli. 2016. "Effect of Freezing Conditions on Fecal Bacterial Composition in Pigs" Animals 6, no. 3: 18. https://doi.org/10.3390/ani6030018