1. Introduction
The Andalusian Black cattle breed is located in the Autonomous Community of Andalusia, Spain, from which it takes its name. It is scattered in two main regions: the first in the Sierra Morena areas of Córdoba and Huelva, and the second in the lowlands of Sevilla, Cádiz, and Huelva [
1] (
Figure 1). Currently, it is classified as an endangered native breed for conservation purposes, with an officially declared population of 1797 animals in 22 officially declared farms as of the year 2020 and an average annual population growth of 6.31% in the period from 2009 to 2020 [
2].
Historically, this breed was specialized for agricultural work, but with the advent of agricultural mechanization, its focus shifted towards meat production, leading it to more mountainous areas and making use of mountain pastures. It has contributed to ecological and social balance along with other breeds. As a result, it has great potential for organic meat production, thus improving the income of producers by offering a 100% native quality brand [
3].
Animals of this breed are characterized by a coat, skin, and hair of uniform black color with variations of “morcillo” black on the lower legs, testicular region, and udders. They also have black mucous membranes and white horns with black tips (
Figure 2 and
Figure 3). Their profile is straight with considerable variation towards convexity, moderate proportion, and a tendency towards hypermetry with marked sexual dimorphism (
Figure 2 and
Figure 3). The established morphological rating system is based on visual evaluation and a point-based method conducted at 24 months of age, with a rating by region from 1 to 10. Animals with a rating below 5 are disqualified. Subsequently, these values are multiplied by weighting coefficients, resulting in classifications from insufficient to excellent [
3,
4].
Since 2004, with the establishment of the Association of Breeders of the Andalusian Black Cattle Breed and the corresponding approval of conservation programs for the breed in 2012 and 2020 by the Ministry of Agriculture, Fisheries, and Food (MAPA), the breed has made a significant recovery thanks to the implementation of a pedigree book and performance monitoring. The breed conservation program focuses on meat production in extensive systems, with seasonal supplementation for industrial crossbreeding with specialized breeds. As a result, most farms obtain purebred or crossbred calves that are sent to commercial feedlots [
3,
6].
One of the advantages of this breed is its reproductive aptitude, as it exhibits high fertility and longevity under challenging production conditions, where adaptability and hardiness play a significant role. Females reach their first heat cycle between 18 and 20 months of age, depending on their development and nutrition, achieving an 80% fertility rate and 95% conception rate with a 1.15% incidence of twin births [
2,
5]. Under traditional weaning, the 6–7-month-old calves weigh between 190 and 220 kg, reaching an average adult weight of 550–600 kg in females and 850–900 kg in males [
2].
Established in 1273 by Alfonso X the Wise, the Mesta Council brought together shepherds from León and Castile, forming an Iberian association. Alfonso X granted them significant privileges, including exemption from military service and the right to testify in trials, as well as rites of passage and pasturage. Over time, additional royal privileges were bestowed upon the Mesta, accompanied by protective measures against farmers. This sparked numerous legal disputes throughout the Middle Ages. In 1836, the Mesta was abolished, partly due to conflicts with Portugal, which rendered many livestock routes impassable [
7].
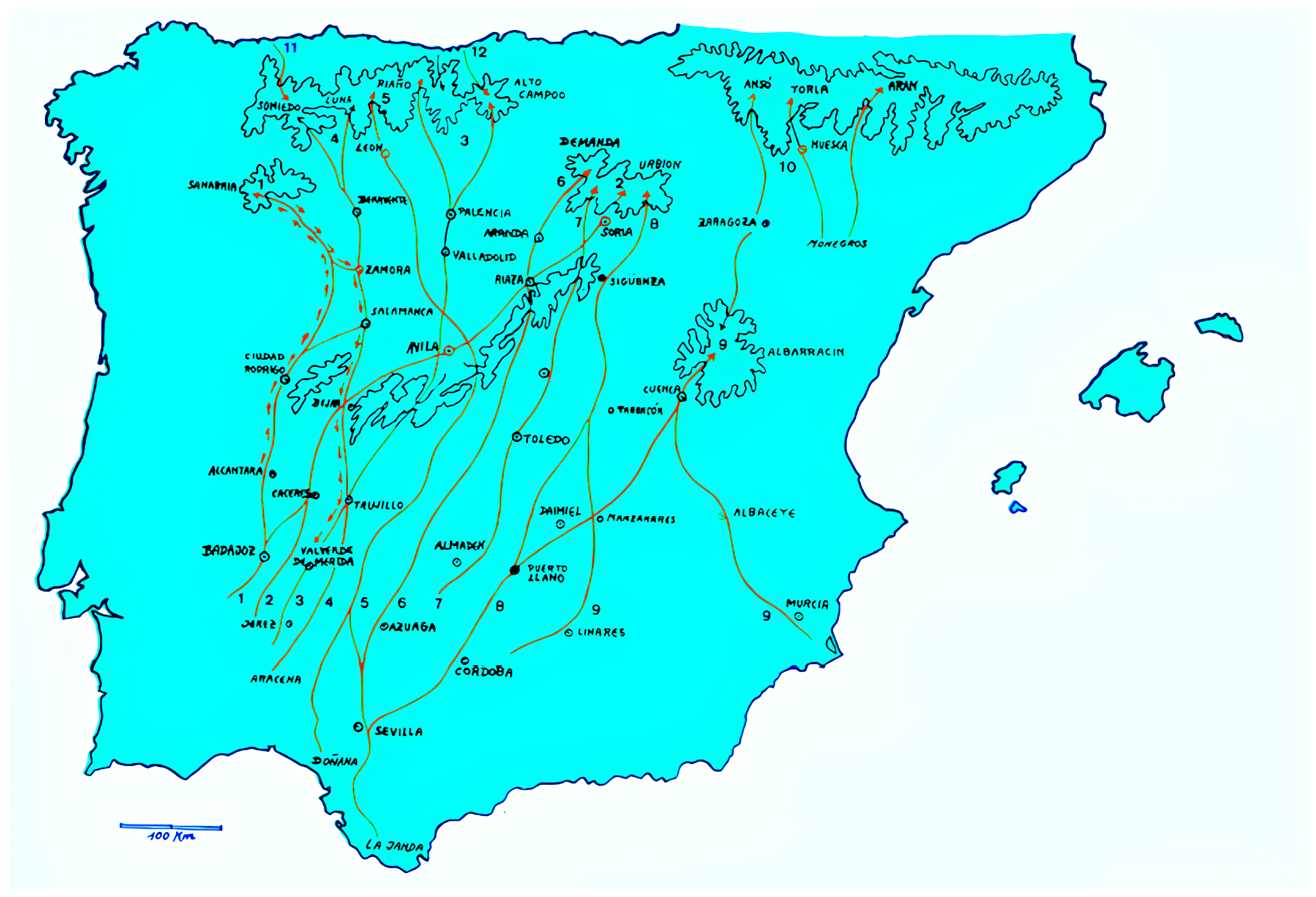
However, the Cañadas Reales played their part as diversity routes of domestic animal genetic resources from the north of the Iberian peninsula to the south, and even transboundarily into Portugal (
Figure 4). These routes played a pivotal role in shaping Iberian livestock breeds. These ancient transhumance routes served as corridors for the seasonal migration of large herds of sheep and parallelly other livestock which joined them for labor or transport, connecting disparate landscapes and fostering genetic exchange among local populations (
Figure 5).
As a result, the movement of livestock along the Cañadas Reales not only influenced the genetic composition of various breeds but also contributed to the establishment of connections and similarities among them. The intricate web of crossbreeding and intermingling that occurred along these routes has left a lasting impact on the diversity and adaptability of Spanish livestock. Today, the legacy of the Cañadas Reales persists in the distinct characteristics and shared traits observed in different breeds, highlighting the enduring influence of historical transhumance on the genetic landscape of Spain’s livestock.
The transhumance routes not only facilitated the movement of large sheep flocks but also served as pathways for the migration of cattle herds [
8]. Cattle played a multifaceted role in medieval Spain, serving not only as a source of meat and dairy but also as essential draft animals for agricultural labor. The intricate network of the Cañadas Reales allowed for the intermingling of cattle breeds from diverse regions, leading to the exchange of genetic material and the emergence of drifted populations. The adaptability and strength of these draft animals were crucial for the agricultural productivity of various regions along the migration routes. As a result, the genetic diversity routes established by the Mesta influenced not only the characteristics of sheep breeds but also shaped the traits and capabilities of cattle, leaving a lasting imprint on the historical and contemporary livestock populations in Spain [
9].
The implemented conservation program’s primary objective is to maintain the breed and improve the maternal line. This means preserving genetic diversity levels (minimizing inbreeding effects) and enhancing maternal reproductive efficiency, with a focus on the calving interval. Also, at the same time, it means preserving ancestral reservoirs of genes and their interconnection routes, which are the only resources that can shed light about the process of configuration that endangered breeds were exposed to.
The primary objective of this study is to develop a model for analyzing pedigree completeness in a downward manner, with a focus on understanding the repercussions of ancestors and founders. The aim is to not only assess the structure of the population and its genetic variability but also to uncover the historical routes and directions that genetic diversity followed during the configuration of the breed. By evaluating the connections between genetic diversity and demographic parameters, the model seeks to measure the existing gene flux and quantify the risk of genetic diversity loss. Additionally, this study aims to determine the endangerment degree of authochtonous bovines and other endangered animal small populations. This comprehensive analysis will serve as a foundation for suggesting effective conservation strategies, taking into account the historical context and dynamics that have shaped the genetic landscape of these populations.
4. Discussion
The Andalusian Black breed experienced a significant population decline historically due to agricultural mechanization and the introduction of foreign meat-focused breeds. Initially described between 1918 and 1944, it was officially recognized as a breed between 1981 and 2002 in the Spanish Livestock Breeds Catalog. Its original focus on both work and meat was altered by these changes [
2,
21].
The Association of Breeders of the Andalusian Black Cattle Breed was founded in 2004 and has managed the pedigree book since 2005. Since 2012, it has been classified as an endangered native breed, prompting a conservation program. As a result, registered animals have risen from 1252 in 2009 to 2121 by 2020 [
2]. Presently, due to its reproductive characteristics, the breed is used for breeding pure animals and for crossbreeding with specialized breeds for veal production [
26].
The Andalusian Black cattle breed has remained in existence across five provinces (Seville, Córdoba, Huelva, Jaén, and Cádiz), spanning 22 municipalities and 28 herds. In contrast, the Mertolenga breed in Portugal is present in 700 herds, with 216 under genealogical control [
27]; the Alentejanean breed is distributed into 120 herds [
28]; and the Maremmana breed, in Italy, is distributed among 309 herds [
29], highlighting a difference in conservation efforts.
The average number of animals per herd in the Andalusian Black breed decreased significantly from 305.54 in the historical population to 88.28 in the current population (2003–2020). In contrast, the Mertolenga breed has an average of 103.5 registered cows per herd, showing a difference in population size trends [
27]. On the other hand, with the Alentejana breed, a similar value of 85.5 ± 63.4 registered cows is observed [
28]. The main cause of this decrease is due to the introduction of foreign breeds (replacement) as well as the use of females for crossbreeding (terminal fattening calves), as has occurred in the Polish Red breed [
30,
31].
In the current population of the Andalusian Black breed, there has been a decrease in the number of bulls and an increase in the number of cows, reflected in the cow-to-bull ratio. This ratio has shifted from 1.38 to 3.17, while the cow-to-bull paired ratio has risen from 26.1 to 36.71. This contrasts with the Polish Red breed, which has a reported ratio of 9.3 [
32] and is a breed of international significance because it is the only native breed in this country with the characteristics of resistance and adaptability, such as a high content of total solids, resistance to harsh environments and diseases (tuberculosis and mastitis), good fertility, and longevity [
30].
Likewise, the male-to-female ratio’s impact on the short-term survival of founder genes is noteworthy, although population contraction or expansion and family size variation have more substantial effects [
33]. The intensity of selection may contribute to the reduction of this ratio, as seen in the Indubrasil and Gyr breeds from 1993 to 2001 in Brazil [
34,
35].
Regarding parameters such as the average number of offspring per paired bull, it increased by 11.58 (from 80.18 to 91.76), with a reduction in cows of 2.65 (from 11.24 to 8.59). These figures surpassed those published for the Polish Red breed, indicating an increasing reproductive rate through the maternal line. The average age at reproduction decreased to 1.64 years in bulls and 2.65 years in cows, suggesting a reduced selection and retention of bulls and an increased selection of cows, along with shorter herd tenure.
The selection of animals for reproduction drastically declined, with a 54.14% decrease in bulls and a 32.67% decrease in cows in the current population. The intensity of selection was 0.58% for males and 32.32% for females, reflecting a reduced interest among breeders in this breed or, for males, the utilization of males from other specialized crossbreeding breeds, similar to the Indubrasil breed [
34].
The number of traced generations for both populations was seven, with Pedigree Completeness Index values greater than 0.5 in the first and second generations. While these values fall below the threshold of 0.6 for reliability in estimates, they have steadily improved since 2000, when the breed was included in the official catalog of Spanish bovine breeds, alongside enhanced management of genealogical records and genetic improvement plans by the association [
2]. These values align with other bovine breeds undergoing conservation processes, such as the Curraleiro Pé Duro in Brazil, Italian Pontremolese, Sardo Modicana, and German Angler and Red and White dual-purpose breeds [
36,
37,
38], but are lower than those in commercial–industrial dairy and beef cattle breeds with extensive record-keeping and genetic evaluation histories [
39,
40,
41,
42,
43,
44,
45].
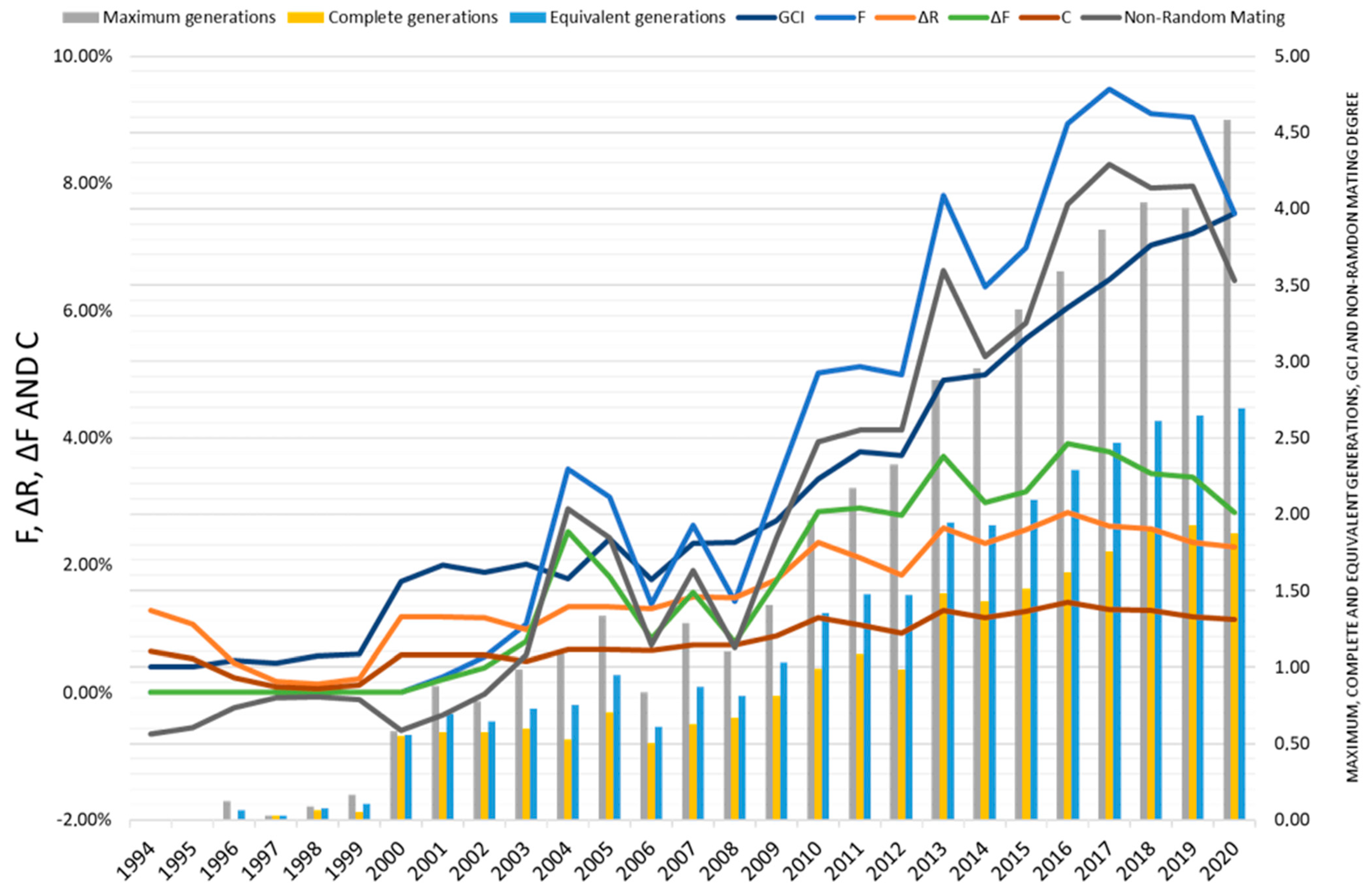
Figure 6 depicts the parameters GCI, F, ΔR, ΔF, and non-random mating, showing increases in all parameters from the year 2000. F, non-random mating, and GCI exhibit irregular increases, particularly in the periods 2003–2004, 2006–2007, and 2012–2013, due to the mating of closely related animals and increased genealogical information [
26]. However, there has been a notable reduction in F since 2019 due to conservation policies [
2]. The average value of F for the current population of the Andalusian Black cattle breed reaches 5.94%, which is significantly higher than other endangered breeds in Spain and Europe [
46,
47,
48]. Higher values were reported in breeds like Mucca Pisana in Italy and Mertolenga in Portugal [
27,
37].
Variable GCI values have been observed across species, ranging from higher values of 9.11 to 9.65 for Spanish, Arabian, and Hispano–Arabian horse breeds [
49] to similar values of 4.08 in the Brazilian Somali sheep breed [
50]. These discrepancies can be attributed to GCI reliance on pedigree integrity and the number of generations, which are strongly influenced by conservation husbandry practices specific to each species, where genealogical registration has historically varied in importance.
For instance, the differing purposes and values of domestic animals play a significant role. Horses, often used for sport, leisure, or transportation, rely heavily on genealogical information for assessing abilities, traits, and value. Conversely, donkeys, presumed to have lost their original function, face challenges due to incomplete pedigrees. Poultry or rabbits, primarily used for meat production, prioritize factors like growth rate, feed efficiency, or carcass quality over genealogical information [
51].
Additionally, variations exist in genetic diversity and domestication levels among domestic animals. Species like cattle, sheep, and goats, with a long history of domestication and selective breeding, benefit from genealogical information for tracing origins, relationships, and genetic improvement. In contrast, cats, dogs, and pigs, with a more recent and complex domestication history, may face challenges in obtaining or interpreting genealogical information if not registered in official studbooks [
52].
Moreover, differences in legal and zootechnical frameworks for genealogical information registration across the European Union further contribute. Directives exist for the breeding and trade of equidae and purebred breeding animals [
53], necessitating breed societies, studbooks, and identification systems. However, no such directives exist for hybrid breeding pigs or other domestic animals, leaving genealogical information registration at the discretion of Member States or breeders [
53].
In the case of the Andalusian Black cattle breed, values have been less than 50% complete since the second generation. However, there has been an increase in GCI over time, indicating that conservation efforts towards registering genealogical information are becoming effective. This may facilitate decision-making for driving conservation strategies based on reliable parameters.
The ΔF, ΔR, and C metrics have exhibited a moderate increase from 1999, reaching a peak in 2016, followed by a trend toward reduction due to the implementation of genealogical record-keeping in 2012 (
Figure 6). ΔF values were 2.71 and 3.17 for the historical and current populations, respectively, and they were comparable to values reported for other endangered Italian breeds like Sarda and Sarda Bruna [
46]. However, compared to the Spanish Bruna dels Pirineus breed, ΔF was notably lower (0.99%) [
46]. ΔR showed a value of 2.1%, similar to values found in the Mertolenga breed (2.05%) in Portugal and Colombian breeds like Horned Costeño, San Martinero, Blanco Orejinegro, and Romosinuano. Furthermore, ΔR values slightly exceeded those of C, which were 2.9% and 3.6% for the historical and current populations, respectively. Both C and ΔR values have displayed a downward trend since 2016, due to measures implemented to control parentage using genetic markers and limiting the use of breeding stock that endangers breed genetic diversity.
The non-random mating values (α) were 0.049 and 0.061 for the historical and current populations, respectively, with positive values observed since 2003, though experiencing a reduction since 2019. This suggests ineffective matings, with individuals having higher-than-average coancestry or relatedness mated more frequently, akin to observations in commercial breeds like Jersey in the USA. Additionally, α values were consistent with F, indicating a pattern of positive assortative mating when mating related individuals.
The effective number of equivalent founders (fe = 91.14) for the breed was comparable to that of the Mertolenga breed in Portugal (fe = 87.9), yet was much higher (+64%) than that reported for the Berrenda Negra breed in Spain (fe = 58) and lower than that of the Berrenda Colorada breed (fe = 140) in Spain. This suggests an imbalance in genetic contribution among founders, which is reflected in
Table 4, where a small number of individuals account for a significant portion of the genetic diversity. Similarly, the effective number of ancestors (fa = 42) was lower than that of the Mertolenga breed and the Berrenda Negra and Colorada breeds. Both fe and fa are influenced by population size, indicating a genetically small population [
27,
54].
The fa/fe ratio in this study was 0.46, which was notably lower compared to several other breeds, including Berrenda Colorada, Berrenda Negra, Morucha, Mertolenga, Lageana, and Tabapua, which exhibited ratios greater than 1 [
27,
46,
54,
55,
56,
57]. Conversely, proportions in the Pinzgau cattle breed and the Marchigiana and Bosmara breeds in Brazil were similar, indicating a genetic bottleneck within the Negra Andaluza breed, affecting its genetic diversity, akin to observations in commercial dairy cattle breeds [
58].
Historically, the Andalusian Black cattle breed may have faced a genetic bottleneck due to factors such as agricultural mechanization, crossbreeding, lack of genetic programs, subpopulation segmentation and isolation, and unstructured breeding efforts [
4], contributing to population decline and genetic variation loss.
Equivalent founder genomes (fg) indicate the maintenance of founder genes in the population for a particular locus, revealing causes of gene loss through segregation [
58]. In the Negra Andaluza breed, the fg value was 46.83, similar to the Mertolenga breed in Portugal [
27]. The fg/fe ratio was 0.51, suggesting the impact of genetic drift, excluding founder contributions to genetic diversity, and was higher compared to the Simmental and Braunvieh breeds in Austria [
59].
In terms of inbreeding and average relatedness increase in the reference population (both parents known), minimal values were observed (0.21% and 0.27%, respectively), indicating the positive effects of genealogical control measures and improvement plans, as evidenced by reductions in F and AR in recent years (
Figure 6).
The Andalusian Black cattle breed exhibited a genetic diversity of 98.93% and a Genetic Conservation Index (total population) of 2.75, akin to other breeds under conservation programs such as Berrenda Negra and Berrenda Colorada in Spain [
54]. However, the loss of genetic diversity was 1.07, which is relatively low compared to commercial dairy breeds experiencing losses between 3 and 11% [
39].
Genetic diversity loss (GDL) due to genetic bottlenecks and drift was relatively low (1.07), being nearly double the GDL due to unequal founder contributions (0.55), as observed in the Guernsey breed [
39]. In contrast, other conservation breeds like Berrenda Negra and Colorada exhibited lower GDL values [
54], indicating genetic bottlenecks and drift as primary causes of genetic diversity loss in the Andalusian Black cattle breed.
Chakraborty [
60] emphasizes the importance of understanding population ecology, demographics, and social behavior to detect population structure. This is particularly relevant for domestic and endangered populations, where anthropologic and administrative components must also be considered. F-statistics, describing inbreeding at the individual (F
IS), subpopulation (F
ST), and population (F
IT) levels are commonly used for this purpose.
For the Andalusian Black breed, analysis was conducted at the herd (28), municipality (23), and provincial (5) levels. F
IS values were 0.036, 0.021, and 0.039, while F
ST was 0.085, 0.069, and 0.011 and F
IT was 0.052, 0.049, and 0.049, respectively. These values indicate a low-to-moderate level of inbreeding and differentiation at all levels, which is comparable to other breeds under conservation programs and intensive production breeds like Brown Swiss, Sahiwal, and Braford [
36,
61,
62,
63,
64,
65].
Positive but small F
IS values suggest some inbreeding within herds, municipalities, and provinces, while positive but small F
ST values indicate some differentiation among these levels. F
IT values, similar to F
IS values, suggest that within-population inbreeding contributes mainly to total inbreeding, with little effect from among-population differentiation [
66]. These values reflect the positive effects of inbreeding control measures in the breed (
Figure 6).
F
ST values close to 0 suggest similar allele frequencies at all levels, indicating a lack of genetic structure. A slight reduction in heterozygosity is observed at the provincial level, while greater reductions are seen at the municipality and herd levels. F
IT analysis reveals an average reduction of 4.9% in heterozygosity at the municipality and provincial levels and 5.2% at the herd level, which is comparable to findings in the Wagyu breed [
67].
The inbreeding levels for herd, municipality, and province were 0.095, 0.078, and 0.022, respectively, similar to coancestry averages of 0.095, 0.079, and 0.022. These values indicate that subpopulations maintain low and adequate levels of inbreeding, aligning with recommendations for conservation.
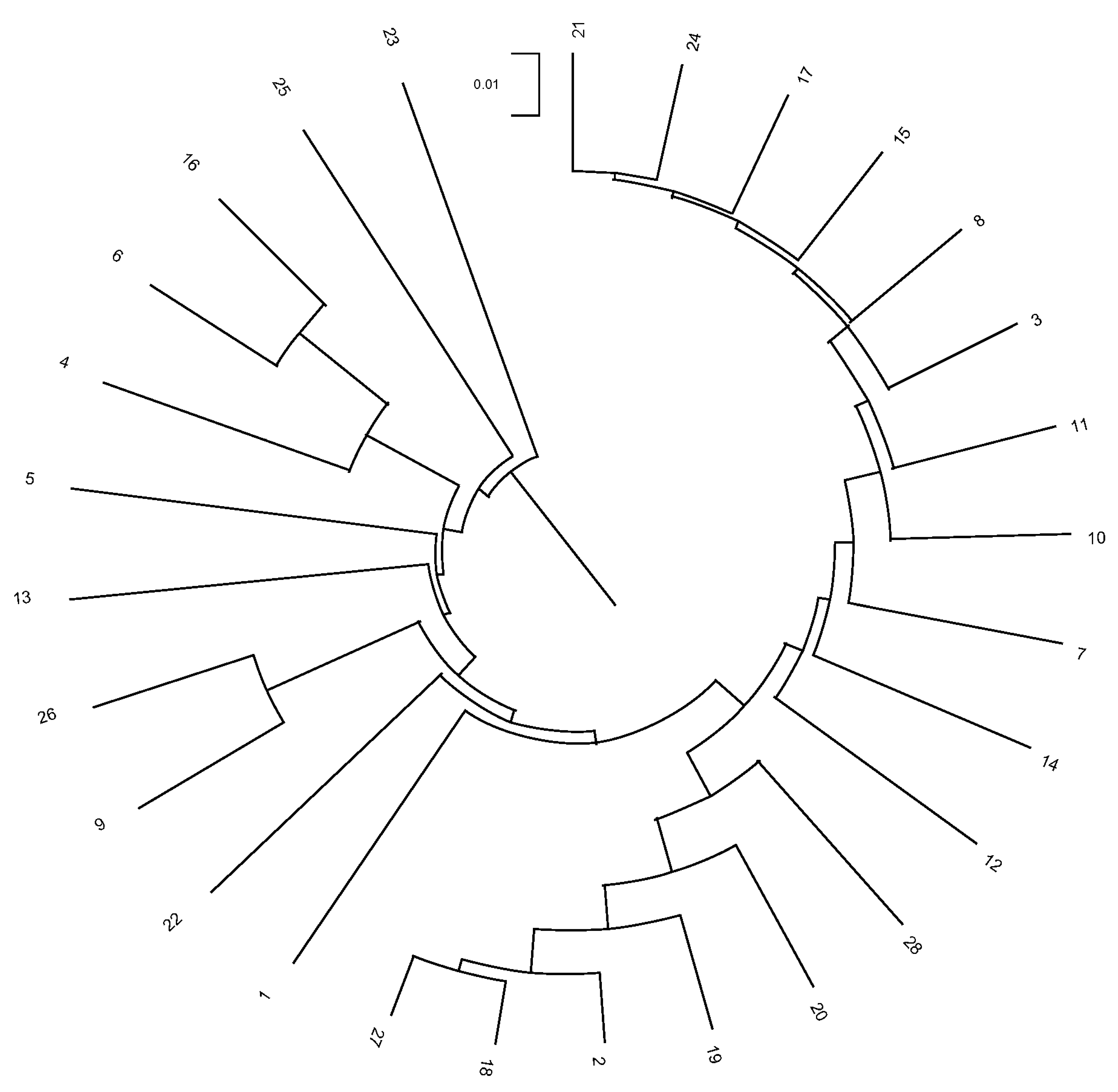
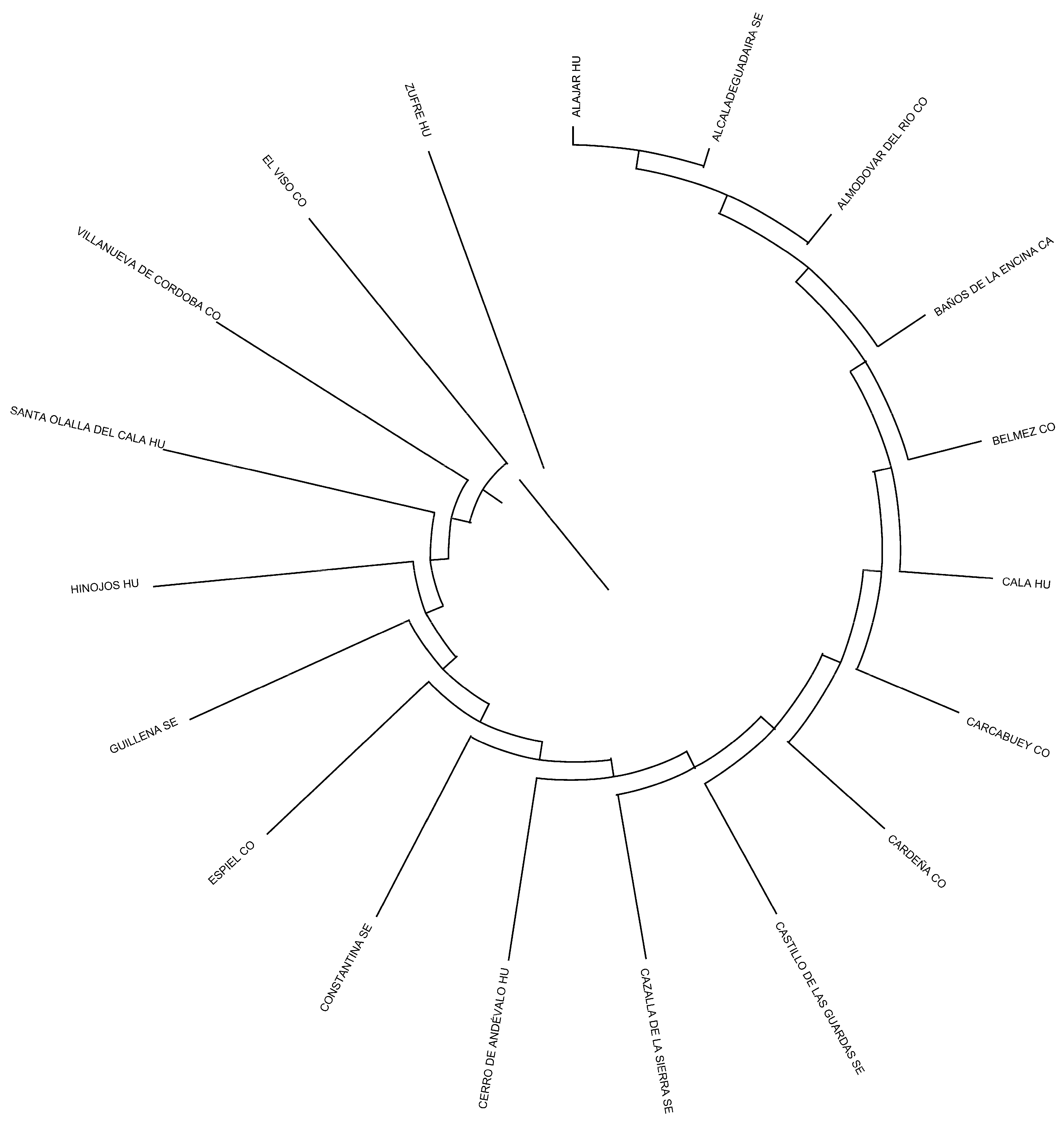
The Nei distances show similarity at the herd and municipality levels (378 and 253, respectively) but considerable distance at the provincial level (10). Clade formation in
Figure 7 and
Figure 8 reveals distinct groupings: Cadiz and Jaen (0.047 and 0.016, respectively) and a third split into Sevilla and Cordoba–Huelva (0.010, 0.007, and 0.007, respectively). This suggests a limited genetic relationship between Cadiz and Jaen with Sevilla, Huelva, and Cordoba (which exhibit a greater genetic relationship).
At the municipality level, Barbate de Franco (Cadiz Province) is closely related to Espiel, Villanueva del Duque, and Cabra in Cordoba. Similarly, municipalities from Huelva and Sevilla appear alongside Cordoba, which are geographically close. In Jaen Province, only Baños de la Encina shows a relationship to municipalities in Sevilla and Huelva, which are also geographically proximate.
The Western and Eastern Sorian Cañadas Reales, ancient routes used by the Mesta, an association of cow and sheep farmers practicing transhumance in Spain, span approximately 800 km and 700 km, respectively [
68,
69]. The Eastern route starts in Soria and extends to Sevilla, crossing territories inhabited by Andalusian Black cattle, such as Cádiz, Jaén, Sevilla, Córdoba, and Huelva. Although not directly connected, the Western Sorian intersects with the Cañada Real de la Plata near Guijuelo, Salamanca [
69].
These routes form part of a network of ancient paths facilitating livestock movement between pastures throughout the seasons [
70]. The Eastern Sorian Cañada Real links to provinces like Sevilla, Huelva, and Córdoba through vías pecuarias, which are reserved paths for livestock passage. These paths, including veredas and coladas, vary in width and length [
55]. Examples include Vereda de Hornachuelos (20.89 m wide, 86.5 km long), Vereda de la Plata (20.89 m wide, 125.5 km long), and Vereda de la Sierra (20.89 m wide, 215 km long), and they connect different provinces through various towns like Hornachuelos, Palma del Río, Peñaflor, Lora del Río, Sanlúcar la Mayor, Villamanrique de la Condesa, and Aracena [
70].
The Andalusian Black cattle belong to the Black Iberian group, alongside breeds like the Avileña–Negra Ibérica and the Preta [
71]. Originating from the ancient Tronco Negro Ibérico, they share characteristics such as a black coat, medium size, and adaptability to harsh environments. Genetic studies suggest a closer relationship with Preta cattle than other Iberian breeds like the Black Avileña, possibly due to a shared genetic reservoir in the Aracena mountains, which are part of the Sierra Morena range along the Spain–Portugal border [
71].
The Sierra Morena, connecting with Portugal’s Serra da Estrela, acts as a natural barrier between the countries and is home to the Aracena mountains, a protected area within the Sierra de Aracena and Picos de Aroche Natural Park. Despite high genetic diversity and low differentiation among these breeds, geographic isolation, environmental adaptation, and selective pressures may have contributed to some genetic differences, as reported for the same and other species [
72,
73,
74].
Nei distances reveal moderate genetic differentiation among herds and municipalities of Andalusian Black cattle, which is likely influenced by geographic distance, limited gene flow, or local adaptation. However, there is minimal genetic variation among provinces, potentially due to historical transhumance or shared selection criteria.
When provinces are considered, genetic clustering illustrates relationships among them and their municipalities. Provinces like Sevilla, Huelva, and Córdoba show greater genetic relatedness, while Cádiz and Jaén exhibit less connection, which is possibly due to geographic isolation or environmental differences.
Territorial segregation plays a crucial role in preserving genetic diversity in endangered native cattle breeds. By restricting movement and interbreeding to specific regions, unique genetic traits can be maintained, preventing dilution of the gene pool from crossbreeding. This approach facilitates focused breeding programs tailored to local environments, ensuring the conservation of valuable genetic adaptations. Overall, territorial segregation supports the long-term survival and sustainability of endangered breeds.
As part of future strategies, exploring the runs of homozygosity in the Andalusian Black cattle population using genomic data is recommended. This approach would significantly enhance the depth of the study and enable the validation of findings across different populations. Leveraging genomic data allows for a more comprehensive understanding of the genetic structure, diversity, and potential inbreeding patterns within this breed. Moreover, analyzing runs of homozygosity can provide valuable insights into the historical demography, selection pressures, and adaptation mechanisms of Andalusian Black cattle. Ultimately, integrating genomic analyses into the research framework will contribute to more informed conservation and breeding strategies aimed at preserving the unique genetic heritage of this endangered breed.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}